










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Habanera de Ciencias Médicas
versión On-line ISSN 1729-519X
Rev haban cienc méd v.9 n.2 Ciudad de La Habana abr.-jun. 2010
CIENCIAS BÁSICAS BIOMÉDICAS
Universidad de Ciencias Médicas de la Habana
Facultad de Ciencias Médicas "Victoria de Girón"
El receptor del factor de crecimiento epidérmico y su papel en el desarrollo tumoral
Epidermal growth factor receptor and its role in tumor development
Tania Lahera Sánchez1, Obdulio Juan González Hernández2
1Especialista Primer Grado en MGI. Residente de 3er año de Inmunología.Santa Catalina núm. 27 entre Delicias y Buenaventura.10 de Octubre. Ciudad de La Habana. 052492626. tania@cngen.sld.cu
2Especialista Primer Grado en MGI. Residente de 2do año de Bioestadística. Santa Catalina núm. 27 entre Delicias y Buenaventura.10 de Octubre. Ciudad de La Habana. 052718294
RESUMEN
El EGFR pertenece a la familia de receptores tirosina quinasa, y está representado por 4 miembros (HER1 o EGFR, HER2, HER3 y HER4), cuyos ligandos son un grupo de factores de crecimiento en especial el EGF y TGF alfa. El EGFR es activado por dimerización, la cual depende de la unión del ligando, aunque también puede ocurrir cuando hay sobreexpresión y alteraciones estructurales del receptor. Además de su función durante la embriogénesis, el EGFR tiene un papel fundamental en el desarrollo tumoral en el que se han encontrado diferentes mutaciones que se correlacionan con un peor pronóstico. En consecuencia, constituye una diana en la terapia de cáncer, mediante anticuerpos monoclonales e inhibidores de la actividad tirosina quinasa. Este artículo ofrece una visión actual del conocimiento sobre EGFR haciendo énfasis en su estructura, mecanismo de activación, papel en el desarrollo normal y tumoral, así como las principales estrategias terapéuticas para su inhibición.
Palabras clave: Receptor del factor de crecimiento epidérmico, activación del receptor del factor de crecimiento epidérmico, embriogénesis, cáncer.
ABSTRACT
EGFR is a member of the tyrosine kinase family and it is represented by 4 members (ErbB1 or EGFR, ErbB2, ErbB3 and ErbB4) whose ligands are a group of factors of growth especially EGF and TGF alpha. EGFR is activated by dimerización, which depends on the union of the ligand, although it can also happen when there are sobreexpresion and structural alterations of the receptor. Besides their role during the embriogenesis, EGFR plays a fundamental role in the tumoral development where they have been different mutations that are correlated with a worse prognosis. In consequence it constitutes a target in the cancer therapy, by means of monoclonales antibodies and tyrosine kinase inhibitors. This article offers a current vision of the EGFR espècially in its structure, activation mechanism, role in the normal and tumoral development, as well as the therapeutic main strategies for its inhibition.
Key words: Epidermal growth factor receptor, activation of epidermal growth factor receptor, embriogenesis,cáncer.
INTRODUCCIÓN
El receptor del factor de crecimiento epidérmico (EGFR, ErbB) pertenece a la superfamilia de receptores localizados en la membrana plasmática que presentan actividad tirosina quinasa intrínseca.1-3
El factor de crecimiento epidérmico (EGF) fue identificado en 1962 por Stanley Cohen, mientras que el receptor fue purificado y caracterizado por el mismo autor en 1980. Fue el primer receptor tirosina quinasa en descubrirse y la mayor parte de los mecanismos de activación de este tipo de receptores fueron establecidos a partir del EGFR.4,5
El EGFR es filogenéticamente muy antiguo, ya que existen homólogos en diversos invertebrados, como por ejemplo el receptor Let-23 en el gusano nemátodo Caenorhabditis elegans y receptor Der en la mosca de la fruta Drosophila melanogaster. Además, se han descrito en invertebrados varios ligandos de estos receptores.6
La señalización a través del EGFR es crucial en el desarrollo embrionario, su papel en el desarrollo epitelial, la proliferación y la organogénesis, ha quedado claramente demostrado. Sin embargo, su importancia no se limita a esto, pues existen múltiples evidencias que sostienen que el EGFR tiene una función fundamental en la transformación y progresión tumoral.
Teniendo en cuenta que el cáncer constituye un problema de salud en Cuba y a nivel internacional, la selección de nuevos blancos para la terapia de cáncer es la tendencia principal de múltiples investigaciones en la actualidad. A partir del papel biológico del EGFR en la tumorigénesis y su alta expresión en neoplasias de origen epitelial, se han ensayado varias estrategias para interrumpir esta cascada de señalización, y se ha convertido en un atractivo blanco terapéutico en nuestros días.
Por todo lo anterior, nos sentimos motivados a realizar este trabajo de revisión donde presentaremos una visión actual del conocimiento sobre EGFR haciendo énfasis en su estructura, mecanismo de activación, principales rutas de señalización, así como su función en el desarrollo normal y tumoral. Por último, resumiremos las principales estrategias terapéuticas para su inhibición.
DESARROLLO
La familia del receptor EGF y sus ligandos
El EGFR ha evolucionado desde un receptor y un ligando en Caenorhabditis elegans, un receptor y cinco ligandos en Drosophila melanogaster a cuatro receptores y múltiples ligandos en Homo sapiens.7
En vertebrados, la familia de receptores EGF, de la que forma parte el EGFR (HER1), está constituida además por otros tres receptores identificados hasta la fecha: HER2 (ErbB2), HER3 (ErbB3) y HER4 (ErbB4).
En los cuatro miembros de la familia del EGFR el dominio extracelular está bastante conservado, como ocurre con el dominio proteína quinasa, salvo en el caso de HER3 que no posee actividad tirosina quinasa específica aunque es capaz de unir ATP y transmitir señales mitogénicas mediante su heterodimerización con otros miembros de esta familia. HER 2 es el receptor preferido para la dimerización con otros miembros de la familia del EGFR, lo que le confiere al complejo heterodimérico mayor estabilidad y potencia en la señalización, comparado con otros dímeros. No se han encontrado ligandos específicos para HER 2, por lo que se ha propuesto que esta proteína funciona solamente como un co-receptor. 4-6, 8
Los ligandos que han sido identificados para los receptores ErbB son:
• Para HER1: Factor de crecimiento epidérmico (EGF), Factor de crecimiento tumoral alfa (TGFá),Anfiregulina (AR), Betacelulina (BTC), Epiregulina (EPR), Factor de crecimiento ligado a heparina (HB-EGF).
• Para HER2: No hay ligando conocido.
• Para HER3: Neuregulinas 1 y 2 (NRG1, NRG2).
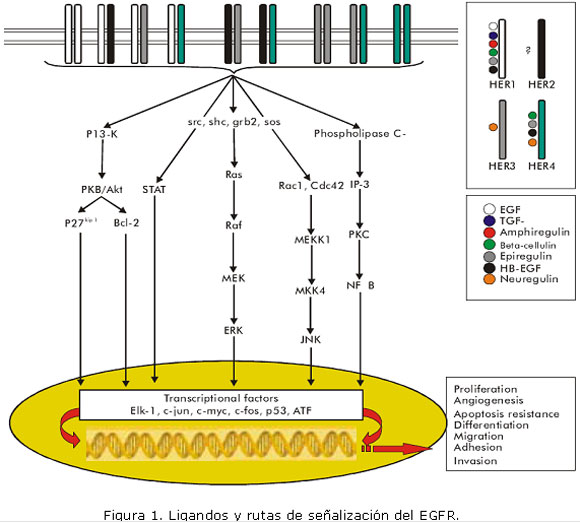
• Para HER4: Betacelulina (BTC), Epiregulina (EPR), Factor de crecimiento ligado a heparina (HB-EGF),Neuregulinas 3 y 4 (NRG3 y NRG4) 8,9 (Figura 1).
Estructura del EGFR (HER1)
En humanos, el gen que codifica el EGFR (HER1) se encuentra en el brazo corto (región p13-p12) del cromosoma 7. Este gen está compuesto por 28 exones que ocupan un segmento de 75 kb y codifican a una proteína precursora de 1 210 aminoácidos que posee una corta secuencia líder hidrofóbica en su extremo N-terminal usada para su inserción en la membrana, siendo posteriormente eliminada por procesamiento proteolítico, Y quedan finalmente 1 186 aminoácidos, los que forman una sola cadena polipeptídica.5
El receptor maduro es una glicoproteína integral de membrana de 170 kDa que está constituida por un dominio extracelular amino terminal donde se encuentra el sitio de unión al ligando, un único dominio transmembranal y un dominio citoplásmico carboxilo terminal, en el que se localiza el sitio catalítico responsable de la actividad tirosina quinasa. 8,10-12
Más de 20 % de la estructura del receptor está representado por residuos glicosilados. Las cadenas glicosílicas del receptor parecen estar implicadas en el correcto plegamiento del mismo, su transporte a la superficie celular y la adquisición de sus funciones. 5
Activación del EGFR
En ausencia de ligandos, los receptores EGF residen en la membrana de la célula de forma inactiva, distribuidos uniformemente por su superficie, en caveolas o colmenas. En condiciones normales, estos receptores tienen muy pocas probabilidades de encontrarse en la superficie celular de forma aleatoria. En cualquier caso, cuando se encuentran en la superficie celular por azar tienen la capacidad de transfosforilarse, actividad que se ve rápidamente inhibida por la actividad fosfatasa basal de la célula.7
Es necesaria la dimerización o la oligomerización del receptor para que presente actividad quinasa, desencadenando cascadas de señalización intracelular. La dimerización puede ser entre dos receptores idénticos (homodimerización) o entre diferentes miembros de la misma familia (heterodimerización).
Si bien el principal mecanismo que provoca la dimerización, y por tanto la activación del EGFR es la unión del ligando,13 existen otras condiciones que pueden provocar la activación como son:
1. Activación de receptores tirosina quinasas, debido al incremento en la expresión del receptor:
El aumento en la expresión es debido a la amplificación de genes o alteraciones transcripcionales. Este aumento del receptor en la superficie celular favorece la colisión azarosa entre moléculas de EGF, con lo cual se favorece su activación. El incremento en la expresión del receptor hace que los niveles de fosforilación sean muy altos, lo cual provoca una saturación de la actividad fosfatasa celular.
2. Activación de receptores tirosina quinasas por alteraciones moleculares:
Las alteraciones moleculares que pueden sufrir los receptores pueden ser de diferentes tipos: mutaciones puntuales y truncaciones. 7
Los aspectos de la sobreexpresión del receptor y las mutaciones serán retomados en el acápite Rol del EGFR en el desarrollo tumoral.
Rutas de señalización
Cuando el ligando extracelular se une al EGFR, se produce la dimerización de este, lo que da lugar a la activación de su tirosina quinasa y la transfosforilación de los residuos de tirosina. La ruta de señalización más conocida y mejor caracterizada de las iniciadas por el EGFR activado es la vía Ras/MAPK, que parece ser imprescindible para la proliferación celular mediada por EGF. Otra vía importante tras la activación del EGFR es la del PI3K (fosfatidil inositol 3 quinasa), la cual genera señales de supervivencia celular y previene la apoptosis. Otras rutas de señalización intracitoplasmáticas que son activadas por el EGFR incluyen a los transductores de señales y activadores de la transcripción (STATs), a la fosfolipasa C gamma 1 (PLC-ã1 y a c-Jun N Terminal quinasa (JNK), las cuales están involucradas en procesos de resistencia a apoptosis, migración celular y proliferación y transformación celular respectivamente (figura 1) .6,8,14-16
Papel de EGFR en el desarrollo normal
La actividad del receptor de EGF es un proceso esencial en el desarrollo del embrión, al estar implicado en la organogénesis de muchos órganos derivados del mesodermo y el ectodermo, tales como cerebro, corazón y pulmón. Frente a su papel crítico en la embriogénesis, en el organismo adulto pierde este papel aunque los receptores de ErbB están involucrados en el desarrollo de los ductos mamarios en la pubertad, la proliferación del lóbulo alveolar en el embarazo y la producción de leche en el postparto. 7,17
Papel de EGFR en el desarrollo tumoral
Entre los principales hallazgos encontrados en múltiples investigaciones sobre cáncer, se encuentran el exceso de expresión de EGFR y las anomalías estructurales en el receptor o sus ligandos.
El EGFR se expresa en células normales en niveles que van desde 20 000 hasta 200 000 copias por célula. Sin embargo, en células de tumores de origen epitelial como el de pulmón, cabeza y cuello, páncreas, ovario, colon, riñón, vejiga y en los gliomas, el EGFR está sobreexpresado y puede llegar a niveles 20 veces mayores que lo normal. También se ha demostrado que la sobreexpresión de EGFR se correlaciona con un peor pronóstico, mayor índice de proliferación, mayor capacidad invasiva y reducción de sobrevida.18
La presencia de un número excesivamente alto de copias de EGFR en la célula provoca un aumento de la sensibilidad a sus ligandos que, incluso a concentraciones muy bajas, son capaces de estimular las células e inducir proliferación celular. Por otro lado, el proceso de internalización de los receptores en estas circunstancias, es más lento porque se excede la capacidad de endocitosis de la célula, por lo que estas no pueden reprimir adecuadamente la transmisión de las señales mitogénicas que se generan de una forma continuada.6
También es frecuente en una gran variedad de tumores la coexpresión aumentada de los receptores homólogos HER2 y HER3. Aunque HER2 no tiene un ligando natural conocido, la heterodimerización con EGFR (HER1) reduce la tasa de endocitosis y degradación, y promueven su reciclaje hacia la superficie celular. La expresión incrementada de HER2 conlleva un incremento de la actividad de la MAPK dependiente de EGFR (HER1) activado por ligando, y aumenta la capacidad de transformación y el crecimiento del cáncer.6
En relación con las anomalías estructurales en el receptor por mutaciones vale decir que se ha identificado un gran número de deleciones en las porciones extracelulares y en menor medida en la porción intracelular del EGFR .Se han reportado 3 deleciones del dominio extracelular que conducen a la definición de los tipos I, II y III (EGFRvI, EGFRvII y EGFvIII)
La variante I (EGFRvI) se caracteriza por la deleción de todo el dominio extracelular. El receptor es constitutivamente activo y no puede ser regulado por el ligando. En la variante II (EGFRvII), ocurre la deleción de 83 aminoácidos de la porción extracelular del receptor. Este receptor mantiene la capacidad de unir el ligando, pero presenta un aumento de su actividad tirosina quinasa. La variante III (EGFRvIII) es la mutación más común e implica la deleción de los exones 2 al 7 y la consecutiva pérdida de los residuos 6 al 273 en el dominio extracelular, lo cual conduce a un receptor constitutivamente activo, que no se regula nega tivamente por endocitosis. Esta mutación también provoca un cambio conformacional en el receptor que estimula la actividad tirosina quinasa y aumenta la capacidad tumorogénica de las células que lo expresan.5,19,20
La actividad incontrolada EGFR se ha implicado en muchos factores del crecimiento tumoral, incluyendo la promoción de proliferación celular, la angiogénesis, la invasión, la metástasis y la supervivencia.21
Proliferación celular
La activación del EGFR induce la expresión de ciclina D1, que promueve la progresión del ciclo celular desde la fase G1 a la fase S, y favorece una rápida y descontrolada proliferación celular.22
Angiogénesis
La señalización a través del EGFR incrementa la producción de factores pro-angiogénicos como el factor de crecimiento del endotelio vascular (VEGF), la interleucina 8 y el factor básico de crecimiento de fibroblastos (b-FGF), y favorece el proceso de angiogénesis que resulta vital para el crecimiento tumoral).23
Invasión celular
Se ha demostrado que la activación del EGFR induce el desprendimiento de las células de la matriz extracelular e induce la síntesis de matriz-metaloproteasas (MMP) en especial MMP9, favoreciendo la degradación de la membrana basal, la invasión celular y el proceso de metastización. 24,25
Apoptosis
Se ha propuesto que la activación del receptor induce la expresión de Bcl2, una proteína anti-apoptótica, favoreciendo la sobrevida de las células tumorales. 8,15
El EGFR como diana terapéutica
Como se ha expuesto hasta ahora, existe un amplio sustento experimental y clínico sobre el papel clave del EGFR en muchos tumores, constituyendo una excelente diana terapéutica. De las múltiples estrategias estudiadas para inhibir al receptor, dos aproximaciones terapéuticas se han mostrado como las más consistentes y son las más ampliamente desarrolladas en la clínica:
- Los anticuerpos monoclonales, que se unen al dominio externo del receptor con alta afinidad, compitiendo con sus ligandos naturales y bloqueando así, la activación de aquel. Ellos comparten una secuencia de distintos mecanismos de acción: unión extracelular, internalización del complejo receptor-anticuerpo, inhibición de la vía de señalización mediada por EGF y potencial estimulación de la respuesta inmunológica.
- Las pequeñas moléculas inhibidoras de la actividad enzimática tirosina quinasa del receptor. (TKIs). Son moléculas sintéticas fundamentalmente derivados quinazolínicos, de bajo peso molecular que interactúan con el dominio tirosina quinasa intracelular de varios receptores, incluyendo EGFR, inhibiendo la fosforilación inducida por ligando. Estas moléculas son generalmente competidores reversibles del sitio de unión al ATP en el dominio catalítico intracelular de la tirosina quinasa.1,8,21
CONCLUSIONES
El receptor del EGF constituye un blanco de gran interés dado su importante papel en el desarrollo tumoral, y representa una diana molecular en las actuales y futuras estrategias terapéuticas contra el cáncer.
REFERENCIAS BIBLIOGRÁFICAS
1. Hubbard SR. EGF receptor inhibition: Attacks on multiple fronts. Cancer Cell. (7):287-288; April, 2005.
2. Normanno N, De Luca A, Bianco C, Strizzi L, Mancino M, Maiello MR, et al. Epidermal growth factor receptor (EGFR) signaling in cancer. Gene. 366(1):2-16;2006.
3. Gschwind A, Fischer OM, Ullrich A. The discovery of receptor tyrosine kinases: Targets for cancer therapy. Nature Rev. Cancer. 4: 361-370;2004.
4. Alfonso G, Cobos C, Rey CA, Tovar J, Alfonso C, Clavijo D. Biología, patobiología, bioclínica y farmacoterapéutica del factor de crecimiento epitelial (EGF) y el estrés celular en la especie humana. Universitas Medica. 47 (3): 258-276;2006. Disponible en: http://med.javeriana.edu.co/publi/vniversitas/serial/v47n3/4%20Biologia%20patobilolg%EDa.pdf [Consultado 12 de mayo 2009].
5. Nair P. Epidermal growth factor receptor family and its role in cancer progression. Current Science. 88(6): 890-898; March 25,2005.
6. Villalobo A, Palomo PI, Ruano MJ. El receptor del factor de crecimiento epidérmico.2000. Disponible en: http://caibco.ucv.ve/caibco/vitae/VitaeCinco/ Articulos/BiologiaCelular/introduc.htm Consultado 12 de mayo 2009].
7. Fernández JC, Pérez V. El receptor de EGF (EGFR): una diana terapéutica para el tratamiento del cáncer y sus inhibidores. Biocancer 3; 2006. Disponible en: http://www.biocancer.com/?q=system/ files/EGFR.pdf [Consultado 12 de mayo 2009].
8. Diaz A, Lage A. Therapies based on inhibitors of the epidermal growth factor receptor: reaching for the future. Biotecnologia aplicada. 24(1):10 -18;2007.
9. Harris, RC, Chung E.Coffey, RJ. EGF receptor ligands. Exp. Cell Res. 284: 2-13;2003.
10. Díaz A, Suárez E, Blanco R, Badía T, Rivero D, López-Requena A. et al. Functional expression of human-epidermal-growth-factorreceptor in a melanoma cell line. Biotechnol. Appl. Biochem. 48: 21-27;2007.
11.Mark J, Stryer L, Tymoczko J, Macarulla JM. Bioquímica. Edit Reverte. Disponible en: http://books.google.com.cu/books?id=HRr4MNH2YssC&pg= PA397&lpg=PA397&dq=receptor+EGF&source=bl&ots=LStFN2ykdz&sig=HpzcLd57Gfh0ccFZMaLPufpu4q8&hl=es&ei=sVSsvGOMWGtgedzqHfDA&sa=X&oi=book _ result&ct=result& resnum =7 [Consultado 14 de mayo 2009].
12. Epidermal growth factor receptor. 2006. Disponible en: http://en.wikipedia.org/wiki/Epidermal_growth_factor_receptor [Consultado 8 de mayo 2009].
13. Cortes-Reynosa P, Robledo T, Salazar EP. Epidermal growth factor promotes epidermal growth factor receptor nuclear accumulation by a pathway dependent on cytoskeleton integrity in human breast cancer cells. Arch Med Res. 40(5):331-8; Jul,2009.
14. Cobb MH. MAP kinase pathways. Progress in Biophysics and Molecular Biology.71 (3-4): 479-500;1999.
15. Cardone MH, Roy N, Stennicke HR, Salvesen GS, Franke TF, Stanbridge E. et al. Regulation of cell death protease caspase-9 by phosphorylation. Science. 282 (5392): 1318-21; Nov,1998.
16. Lo HW, Hsu SC, Hung MC. EGFR signaling pathway in breast cancers: from traditional signal transduction to direct nuclear translocalization. Breast Cancer Res Treat. 95(3):211-8; Feb,2006.
17. Wells A.EGF receptor. Int J Biochem Cell Biol. 31(6):637-43; Jun,1999.
18. González G, Lage A. Cancer Vaccines for Hormone / Growth Factor Immune Deprivation: A Feasible Approach for Cancer Treatment. Current Cancer Drug Targets. (7): 191-201;2007.
19. Wong AJ, Ruppert JM, Bigner SH, Grzeschik CH, Humphrey PA, Bigner DS, et al. Structural alterations of the epidermal growth factor receptor gene in human gliomas. Proceedings of the National Academy of Sciences of the United States of America. 89 (7):2965-9; Apr,1992.
20. Humphrey P, Gangarosa L, Wong AJ, Archer G, Lund-Johansen M, Bjerkvig R. et al. Deletion-mutant epidermal growth factor receptor in human gliomas: effects of type II mutation on receptor function. Biochemical and Biophysical Research Communications., 178 (3): 1413-20; Aug, 1991.
21. Josep Tabernero. Anticuerpos monoclonales inhibidores de EGFR. Disponible en:
http://webcasts.prous.com/CNIO2005/contents/articles /tabernero.pdf [Consultado 12 de mayo 2009].
22. Lin SY. et al. Nuclear localization of EGF receptor and its potential new role as a transcription factor. Nature Cell Biol. 3: 802-808;2001.
23. Kedar D, Baker C, Killion J, Dinney C, Fidler I. Blockade of the epidermal growth factor receptor signaling inhibits angiogenesis leading to regression of human renal cell carcinoma growing orthotopically in nude mice. Clin. Cáncer Res. 8, 3592-3600;2002.
24. O-Charoenrat P, Rhys-Evans P, Modjtahedi H, Court W, Box G,Eccles S. Overexpression of epidermal growth factor receptor in human head and neck squamous carcinoma cell lines correlates with matrix metalloproteinase-9 expression and in vitro invasion. Int J Cancer. 86: 307-317;2000.
25. Price JT, Wilson HM, Haites NE. EGF increases in vitro invasion, motility and adhesion interactions of the primary renal carcinoma cell line, A704. Eur J Cancer. 32A:1977-1982;1996.


{kind=link}