










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Históricamente, el estudio del Neógeno en Cuba ha sido de gran importancia para las Geociencias. Las rocas de la cobertura del Neógeno ocupan una extensión considerable en el archipiélago cubano, prácticamente desde el extremo occidental hasta el oriental, cubriendo alrededor de la tercera parte de la superficie del territorio nacional. Estas rocas se caracterizan por un total predominio del origen sedimentario y la abundancia de macro y microfósiles, especialmente de foraminíferos.
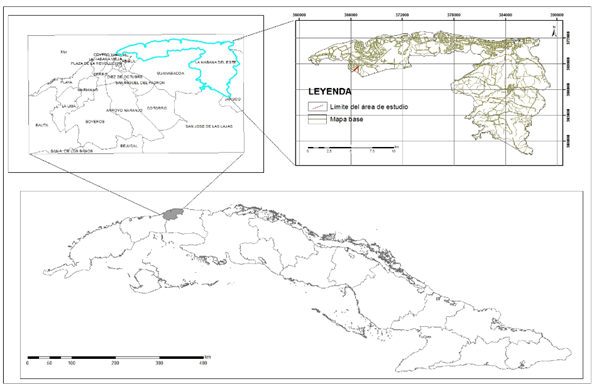
Geográficamente, la localidad cotipo de la Formación Cojímar, designada por Brönnimann y Rigassi (1963) se ubica en un corte de 5 m x 70 m en el lado occidental de la carretera Casa Blanca-reparto Camilo Cienfuegos, en el lado oriental del Hospital Naval, provincia La Habana. Posee coordenadas Lambert x=363 450, y=369 700, en la hoja topográfica La Habana, 3785 III (Albear & Iturralde-Vinent 1985).
Desde el punto de vista geológico, la Formación Cojímar descansa concordantemente sobre las formaciones Colón (parte indiferenciada y su Miembro Coliseo) y Jaruco, y discordantemente sobre las formaciones Consuelo, Encrucijada, Guanajay, Jabaco, Grupo Nazareno (parte indiferenciada), Peñalver, Punta Brava, Tinguaro, Vía Blanca, y los grupos Mariel (formaciones Madruga y Capdevila) y Universidad (formaciones Príncipe y Toledo), y las ultramafitas. Está cubierta concordantemente por la Formación Güines y discordantemente por Vedado y Villaroja. Transiciona lateralmente a las formaciones Caobas, Güines y Paso Real (Iturralde-Vinent 2011).
Esta unidad está compuesta por dos tipos litológicos. Presenta margas calcáreas a margas arcillosas que transicionan hasta gredas de color gris, las cuales se alteran al amarillo crema oscuro hasta el amarillo blancuzco. Su textura es comúnmente gruesa, granular en las gredas, producto de la presencia de arena fina. El otro tipo de roca común son las calizas biógenas de grano fino a medio, entre casi puras y arcillosas, sin transición con las margas. Su color es blanco a blanco crema y su textura es fina a media (Iturralde-Vinent 2011).
Entre los estudios dedicados a la Formación Cojímar se destaca el de Palmer (1934) en el que se describe originalmente la formación, y se ofrece su ubicación geográfica, así como la descripción litológica y geológica. Años más tarde, el propio autor (Palmer 1940, 1941) redescribe muy detalladamente la litología de la formación y por primera vez reporta los foraminíferos bentónicos y el ambiente de deposición. Con posterioridad otros autores aportan nuevos datos, ejemplo Bermúdez (1950) es el primero que correlaciona esta formación con otras unidades litoestratigráficas en Centroamérica, Haití y Florida, y establece su localidad tipo y cotipos.
Brönnimann & Rigassi (1963) e Iturralde (1969a, 1969b) describen los foraminíferos planctónicos en la formación; este último autor además caracteriza con mayor nivel de detalle el ambiente de deposición y establece por primera vez las biozonaciones a partir de foraminíferos planctónicos. Linares & Zuazo (1985) reportan por vez primera la presencia de ostrácodos en la formación.
Como se puede apreciar, la mayoría de los trabajos relacionados con la Formación Cojímar estuvieron encaminados a definir y esclarecer la litología y su posición geológica. Solo dos estudios, realizados por Palmer e Iturralde, se enfocan hacia su ambiente de deposición.
Según plantean Molina (2004), Smart, Thomas & Ramsay (2007) y Ravichandran et al. (2020), la Paleoecología intenta deducir las características ambientales a partir de las asociaciones de microfósiles, los cuales son considerados como excelentes marcadores de las condiciones paleoambientales del medio en que habitan. Los foraminíferos bentónicos son indicadores fiables de los cambios en las condiciones paleoecológicas (Gebhardt 1999; Drinia et al. 2007; Holcová & Zágoršek 2008; Hohenegger et al. 2009; Zágoršek et al. 2009; Gupa, Singh & Verma 2013; Nouradini et al. 2015).
Por considerar que los estudios paleoecológicos existentes sobre la Formación Cojímar a partir de géneros o especies de foraminíferos son insuficientes para mostrar la evolución paleoambiental de esa unidad en el intervalo de deposición, se plantea como objetivos de este estudio actualizar la taxonomía y sistemática de los taxa de foraminíferos planctónicos y bentónicos, reevaluar la biozonación con foraminíferos planctónicos y aportar datos sobre el ambiente de sedimentación en la Formación Cojímar
MATERIALES Y MÉTODOS
Se llevó a cabo un muestreo detallado en el hipoestratotipo 1 el cual se encuentra situado en el lado oriental del Hospital Naval (Figura 1). Cada 15 m se tomaron tres muestras y se identificaron debidamente para su posterior determinación. En el laboratorio, el material fue sumergido en agua corriente durante 24 h para su disgregación. Transcurrido ese tiempo, se procedió a introducirlas en peróxido de hidrógeno, para la eliminación de arcilla. Las muestras se prepararon mediante la técnica de lavado con tamices de malla de luz decreciente (1; 0,125; 0,8 y 0,063 mm). Luego fueron secadas en una estufa, a 80oC. Finalmente, se realizó la separación de los microfósiles, que fueron adheridos a los porta-objetivos con pegamento de tipo tragacanto, para su posterior determinación. El estudio cuantitativo se realizó en la segunda y tercera fracciones. Las muestras fueron analizadas en un microscopio estereoscópico Discovery V8 ZEISS©, con aumentos de 10x a 80x.
Como parte del trabajo de gabinete se procedió a realizar una exhaustiva revisión de la literatura disponible relacionada con el tema (Palmer 1934, 1940a, 1940b, 1941a y 1941b; Bermúdez 1950; Brönnimann & Rigassi 1963; Iturralde 1969a y 1969b, Piotrowska et al. 1981; Kennett y Srinivasan 1983; Albear & Iturralde-Vinent 1985; Linares & Zuazo 1985, Franco & Delgado 1998). Además, se consultó literatura especializada como Ellis & Messina (1940); Bermúdez (1949, 1961), el Léxico Estratigráfico de Cuba (Franco et al. 1992; De Huelbes 2013); Sansores & Flores-Covarrubias (1972); Stainforth et al. (1975); Bolli, Beckmann & Saunders (1994); Molina (1996); Boersma (1998); Franco & Delgado (1998); Murray (2006), Compendio de Geología de Cuba y del Caribe (Iturralde Vinent 2011); Jones (2014); Domínguez-Samalea, Pérez & Rojas (2015), así como la obra inédita Características del Neógeno en el archipiélago cubano, de Gutiérrez, Furrazola & Díaz. La organización sistemática de las especies de foraminíferos planctónicos ha estado sustentada por la sistemática de Loeblich & Tappan (1988) y otras fuentes bibliográficas como Bolli, Saunders & Perch-Nielsen (1985); Iaccarino et al. (2007) y Boudagher-Fadel (2013). En este trabajo se ha asumido el esquema bioestratigráfico de Wade et al. (2011), que adopta biozonaciones y bioeventos calibrados contra la escala de tiempo de polaridad geomagnética y astronómica. La organización sistemática de las especies de foraminíferos bentónicos queda establecida según la clasificación de Pawlowski, Holzmann & Tyszka (2013). Para todas las especies de foraminíferos, tanto planctónicos como bentónicos, se consultaron bases de datos especializadas disponibles en internet, tales como:
RESULTADOS Y DISCUSIÓN
Sistemática y Clasificación
De acuerdo con la sistemática de Bolli, Saunders & Perch-Nielsen (1985); Loeblich & Tappan (1988); Iaccarino et al. (2007) y Boudagher-Fadel (2013), se determinaron 40 taxa de foraminíferos planctónicos agrupados en tres superfamilias, cinco familias, dos subfamilias y 12 géneros (Información Complementar, A1). Según la clasificación establecida por Pawlowski et al. (2013) para foraminíferos bentónicos, se reportan 226 taxa, agrupados en seis órdenes, 26 superfamilias, 54 familias, 48 subfamilias y 102 géneros (Información Complementar, A2). En negrita se destacan los nuevos reportes micropaleontológicos para la formación estudiada. Se constataron que 39 taxa sufrieron cambios de nomenclatura y clasificación (Tabla 1).

Se reportan taxones de foraminíferos que no habían sido descritos anteriormente para la formación. Dentro de los planctónicos, se encuentran Catapsydrax cf. C. dissimilis, Globigerinoides cf. G. obliquus, G. primordius y Hastigerina praesiphonifera. En los bentónicos se reportan, Planulina cf. P. mantaensis, Anomalinoides flintii, Amphistegina tuberculata, Dentalinoides sp. Como nuevo reporte para Cuba se encuentra el taxón Gyroidinoides cibaoensis (Figura 2).
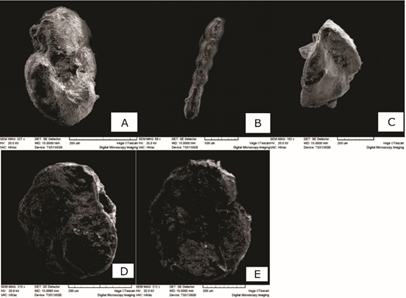
Descripción de la especie Gyroidinoides cibaoensis
Gyroidinoides cibaoensis (Bermúdez 1949) (Sarkar & Gupta 2009, Lám. 5, Figura 6)
Giroidina cibaoensis (Bermúdez 1949)
Referencia tipo: Bermúdez 1949. (Pág. 252, Lám. 17, Figuras 61-63).
Descripción: Testa plano-convexa, lado dorsal plano, lado ventral moderadamente convexo, margen periférico redondeado; unas 15 cámaras forman toda la testa, las primeras son indistintas, 10 cámaras en la última vuelta, la superficie de la cámara casi plana dorsalmente; suturas estrechas, ligeramente oblicuas, al ras con la superficie, claramente visible tanto en la superficie dorsal como en la ventral; ombligo muy pequeño; pared gruesa, lisa, pulida y brillante, no visiblemente perforada; apertura alargada, en la base de la cara septal de la última cámara. Tamaño: Diámetro 0,62 mm. Altura 0,37 mm (Bermúdez 1949).
Distribución Estratigráfica y Paleoambiental: Mioceno Superior. Frecuente en los sedimentos de las formaciones Gurabo y Mao de República Dominicana (Bermúdez 1949). Bhaumik et al. (2007) plantea que esta especie se encuentra presente en bajas concentraciones de oxígeno (Gupta & Thomas 1999; Gupta et al. 2008), suministro de alimentos limitado o en pulsos (Mackensen et al. 1995; De Rijk, Troelstra & Rohling 1999) y oligotrófica (Singh & Gupta 2004)
Biozonación
El análisis bioestratigráfico llevado a cabo en este trabajo ha permitido examinar la mayoría de las biozonas de foraminíferos planctónicos del Mioceno Inferior parte alta - Mioceno Medio parte baja, que figuran en el cuadro bioestratigráfico de Wade et al. (2011).
En las muestras estudiadas de la Formación Cojímar fueron reconocidas las biozonas siguientes que corresponden al Mioceno Medio:
M4a C. dissimilis (17,62 Ma), marcada por la última aparición de C. dissimilis y Dentoglobigerina venezuelana como marcador de zona de rango parcial dentro de la biozona M4a, las cuales corresponden al Mioceno Inferior parte alta.
M6 Orbulina suturalis (15,10 Ma)
M7 Fohsella peripheroacuta (14,23 Ma), marcada por la primera aparición de F. peripheroacuta.
M8 Fohsella praefohsi (13,74 Ma)
Biozona Fohsella fohsi, que coincide con lo reportado por Albear & Iturralde-Vinent (1985) y Linares & Zuazo (1985).
Debido a que no se pudo seguir el registro bioestratigráfico continuo de F. fohsi, no fue posible definir el tope y la base de dicha biozona. No fueron registradas las biozonas M5a Praeorbulina sicana y M5b Praeorbulina glomerosa. Esto pudiera deberse a que el muestreo fue realizado en un solo afloramiento, por lo que no se descarta la posibilidad de que estas especies estén presentes en otros afloramientos de la formación.
Paleoambiente
Los ambientes marinos se clasifican tanto por su profundidad y distancia a la costa como las comunidades biológicas (biocenosis) que los ocupan (Corbí 2010).
En las últimas décadas, algunos autores han sugerido que existe una relación directa entre la forma de la concha de los foraminíferos bentónicos y el microhábitat en que estos se desarrollan, es decir, su modo de vida (Corbí 2010). De acuerdo con estos criterios, las especies trocoespiraladas redondeadas, biconvexas o planoconvexas son características de un modo de vida epifaunal, mientras que las planiespiraladas redondeadas, esféricas, aplanadas ovoides, ahusadas-cilíndricas y aplanadas-ahusadas tienen modo de vida infaunal. Bernard (1986) -citado en Corbí 2010- plantea que la distribución de los morfogrupos infaunales y epifaunales puede ser indicativa de las condiciones de oxigenación del fondo, de modo que la abundancia de foraminíferos infaunales dentro de una asociación puede ser un indicador de condiciones de baja oxigenación sobre el fondo marino.
En las Tablas 2 y 3 se muestran los foraminíferos bentónicos de la Formación Cojímar, agrupados según el morfogrupo y microhábitat al que pertenecen.
Tabla 2 Foraminíferos bentónicos registrados en la Formación Cojímar correspondientes a los morfogrupos que pertenecen al microhábitat (modo de vida) epifaunal - infaunal somero
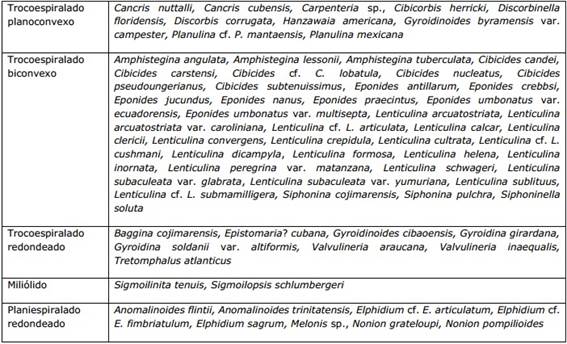
Tabla 3 Foraminíferos bentónicos registrados en la Formación Cojímar correspondientes a los morfogrupos que pertenecen al microhábitat (modo de vida) infaunal profundo

Las asociaciones de foraminíferos bentónicos en la Formación Cojímar están constituidas por morfogrupos mixtos infaunales y epifaunales, en los que predominan los morfogrupos infaunales, indicando esto condiciones de baja oxigenación sobre el fondo, tal como se mencionó anteriormente. Asimismo, dichas asociaciones, están dominadas por especies típicas de ambientes nerítico externo a batial superior, como Bolivina floridana, Cancris nuttalli, Cibicidoides matanzasensis, Cibicidoides sp., Cibicorbis herricki, Gyroidinoides byramensis var. campester, Planulina mantaensis, P. mexicana, Pullenia quinqueloba, Sigmoilopsis schlumbergeri, Trifarina bradyi, Cibicidoides mexicanus, Karreriella bradyi, Sphaeroidina bulloides y Uvigerina pigmea. Además, se encuentran reportadas las especies Eponides antillarum y Nonion grateloupi, características de ambientes nerítico interno.
Las asociaciones de Amphistegina lessonii, Bolivina imporcata, Cassidulina crassa, C. laevigata y C. subglobosa, presentes en zonas nerítica media y, por último, los taxa G. byramensis var. campester, P. mantaensis y Rectuvigerina transversa son exclusivos de ambiente batial superior (van Morkhoven, Berggren & Edwards 1986; Holbourn, Henderson & Macleod 2013; Aubry & Berggren 2011).
Estos datos indican que los sedimentos del Mioceno Inferior parte alta -Mioceno Medio parte baja de la Formación Cojímar fueron depositados en dos ambientes: nerítico y batial superior, entre profundidades de 100 m-500 m (Tabla 4).

Estos resultados corroboran los reportes de Piotrowska et al. (1981), Albear & Iturralde-Vinent (1985) e Iturralde-Vinent (2011). Los citados autores refieren el ambiente de sedimentación entre los 100 m y 300 m de profundidad; sin embargo, en el presente trabajo las asociaciones de foraminíferos bentónicos extienden el ambiente de sedimentación hasta los 500 m de profundidad.
De forma general, las testas de los foraminíferos analizadas presentan un buen estado de conservación, sin signos de abrasión o rotura, de modo que gran parte de las asociaciones registradas se pueden considerar como acumuladas.
CONCLUSIONES
Se actualizó la clasificación y sistemática de las especies y categorías básicas superiores de foraminíferos planctónicos y bentónicos. Se obtuvo un listado de 40 especies de foraminíferos planctónicos y 226 especies de foraminíferos bentónicos.
Se reportaron nuevas especies para la formación, tales como C. cf. C. dissimilis, G. cf. G. obliquus, G. primordius, H. praesiphonifera, P. cf. P. mantaensis, A. flintii, A. tuberculata, Dentalinoides sp. Se obtiene G. cibaoensis como nuevo reporte para Cuba.
Se reconocieron las biozonas de foraminíferos planctónicos siguientes: M4a C. dissimilis, M6 O. suturalis, M7 F. peripheroacuta, M8 F. praefohsi y por último la biozona correspondiente a F. fohsi.
Se determinó la profundidad del ambiente de sedimentación para esta unidad entre 100 m - 500 m de profundidad.
Las asociaciones de foraminíferos bentónicos están compuestas por morfogrupos mixtos infaunales y epifaunales, en las que dominan las formas infaunales, lo que indica condiciones de baja oxigenación sobre el fondo.
Estos resultados contribuyen a la actualización del Léxico Estratigráfico de Cuba.

