










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
De acuerdo con Smol y Stoermer (2010) las diatomeas constituyen un importante grupo de microfósiles formadores de rocas. Evolucionaron rápidamente durante el Cenozoico y, a diferencia de otros microfósiles, se encuentran en sedimentos marinos, salobres y lacustres y, por lo tanto, son útiles para la datación y la correlación de sedimentos acumulados en distintos ambientes. La familia Trochosiraceae constituye un importante grupo de diatomeas bentónicas con una edad relativamente reducida, aunque ampliamente distribuida en el Paleógeno (Zhuze, Proshkina-Lavrenko y Sheshukova 1949).
Las diatomeas conforman uno de los grupos taxonómicos más numerosos, notables (Hernández-Almeida y Siqueiros-Beltrones 2008) y productivos (Booth 1986; MacIntyre, Geider y Miller 1996). Sin embargo, en Cuba el conocimiento de la flora fósil de diatomeas es parcial y para muchas regiones, entre las que destaca la Formación Senado, se carece de estudios. La escasez de información florística se convierte en una limitante para los estudios biogeográficos de diatomeas y tiene como consecuencia la subestimación de su distribución geográfica (Vanormelingen, Verleyen y Vyverman 2008). En Cuba, el estudio de las diatomeas marinas es insuficiente y se ha limitado en dar a conocer los listados de los géneros y especies planctónicas y bentónicas presentes en diversas regiones del país (López y Vinogradova 1972, 1974; Nagy y Coutin 1976; López y Borrero 1977; Hajós, Nagy y Radocz 1980; Moreira et al. 2007).
La Formación Senado que al inicio se llamó “Olistostroma Senado” tiene cierto grado de desarrollo en afloramientos y se registra por algunos pozos petroleros al sur de la Sierra de Cubitas (Flores y Auer 1949; Belmustakov et al. 1981). De esta unidad sólo se había reportado anteriormente fauna fósil redepositada del Cretácico. Por su definida posición estratigráfica se le ha asignado una edad Eoceno Medio parte alta, fundamentalmente por estar situada sobre las formaciones El Embarcadero y Lesca en el pozo Pontezuela No. 1.
Debido al escaso conocimiento existente de las diatomeas fósiles en Cuba y en particular en la unidad Senado, se hizo necesaria esta investigación. Con base a lo anterior, el objetivo de este estudio es dar a conocer el primer registro de Distephanosira architecturalis (Brun) Gleser en la Formación Senado, provincia Camagüey, Cuba Central proporcionando su descripción con base en microscopia biológica.
MATERIALES Y MÉTODOS
Para el desarrollo de este trabajo se estudiaron diversos materiales geológicos procedentes de la zona central del territorio cubano. Para ello se recopilaron los principales trabajos publicados e inéditos que tratan sobre esta región y se consultaron de forma directa diferentes especialistas que han trabajado la geología de Cuba Central. Posteriormente, se realizó un análisis crítico de la información previa y se examinó la literatura especializada con un específico grado de estudio geólogo-paleontológico para la región de Cuba Central.
De acuerdo con De Huelbes (2013) la litología diagnóstica de la unidad comprende una secuencia olistostrómica policomponente, constituida por olistolitos de serpentinitas y calizas en una matriz brechoso-conglomerática con clastos de vulcanitas, gabroides, calizas, areniscas con intercalaciones de capas y paquetes de areniscas, grauvacas y areniscas y limolitas serpentiníticas. Yace discordantemente sobre las formaciones El Embarcadero, Lesca y las del Grupo Remedios. Está cubierta discordantemente por la Formación Nuevitas.
La preparación de las muestras fue llevada a cabo en el Laboratorio de Estratigrafía y Micropaleontología del CEINPET y estuvo basada en la Norma Ramal para la preparación de Láminas Delgadas (SD) y sección en suspensión (SP) de muestras de rocas del departamento de muestrería del CEINPET. El procesamiento de muestras para el aislamiento del taxón y residuos de paredes orgánicas se realizó de acuerdo con el procedimiento estándar para procesar sedimentos marinos terrígenos con enriquecimiento en líquido pesado.
La primera etapa del tratamiento de rocas incluyó: 1) limpieza del material de muestra sólido de la contaminación de la superficie; 2) trituración preliminar de la roca seca (peso de aproximadamente 100-150 g); 3) hervir en una solución de pirofosfato de sodio (Na4P2O7 * 10H2O) para la desintegración, seguido de lavado (cada 2 horas) para eliminar las partículas de arcilla; 3) centrifugación de la fracción aislada en un líquido pesado (K2CdI4) con una densidad de 2.3 g/cm3 (10 min. a una velocidad de 2000 rpm) para la separación de componentes biosilíceo de las partículas minerales más pesadas; 4) lavado de la fracción ligera del líquido pesado por centrifugación. Se realizaron preparaciones estándar para el análisis de diatomeas para su observación en el microscopio. Se utilizaron cubreobjetos de 18 mm x 18 mm y un medio adherente especial Bálsamo de Canadá con un índice de refracción de 1.54.
La segunda etapa, incluyó la selección de microfósiles de paredes orgánicas, que consistió en: 1) exposición al 70% de ácido fluorhídrico (HF) para disolver componentes silíceos; 2) exposición al ácido clorhídrico para eliminar geles de fluorosilicato, y luego lavar la muestra en agua destilada y añadirle glicerina. Las observaciones de los taxones se realizaron bajo el microscopio biológico MEIJI. Las fotomicrografías fueron tomadas con una cámara automática Infinity, modelo DK5000, acoplada a los microscopios triloculares para láminas delgadas.
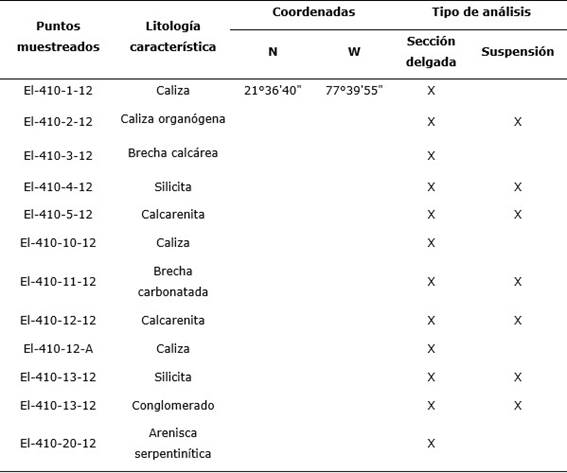
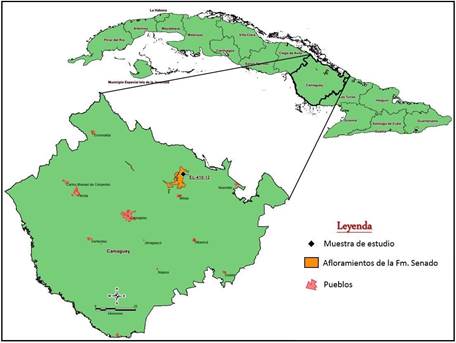
Ubicación geográfica de los pozos y tipos de muestreos
Fueron analizadas 12 muestras del área de Cairije (EL, colectadas por el geólogo Evelio Linares en el año 2012) correspondientes a la Formación Senado (Tabla 1, Figura 1). Todas se examinaron mediante el método de sección delgada, y siete por suspensión.
Determinaciones petrográficas
El método seguido para el estudio petrográfico fue la observación visual bajo el microscopio óptico y biológico. Para realizar las descripciones petrográficas de las rocas carbonatadas se tuvo en cuenta la clasificación de Dunham (1962), modificada y ampliada por Embry y Klovan (1971) (Figura 2).

Figura 2 Clasificación de las calizas, basadas en su textura deposicional según Dunham (1962) modificada y ampliada por Embry y Klovan (1971).
Determinaciones paleontológicas
El contenido diatomológico se determinó mediante la consulta a la siguiente literatura especializada: Schmidt et al. (1874); Zhuze, Proshkina-Lavrenko y Sheshukova (1949); Fenner et al. (1978); Fenner (1985); Makarova et al. (1992); Gladenkov y Barron (1995). Además, se utilizó el apoyo de bases de datos online ( http://www.algaebase.org , https://www.diatombase.org ).
RESULTADOS Y DISCUSIÓN
Descripción de la especie Distephanosira architecturalis
Phylum Bacillariophyta (Haeckel 1878)
Clase Coscinodiscophyceae Round y Crawford (1990)
Orden Paraliales Crawford en Round et al. 1990
Familia Paraliaceae Crawford en Round et al. 1990
Género Distephanosira Gleser, 1992
Referencia tipo: Distephanosira architecturalis (Brun) Gleser nov. comb. En Makarova, Gleser, Moisseeva y Nikolaev, 1992: 68, pl. 56, figs. 1-9.
Sinonimias: Melosira architecturalisBrun 1892; Schmidt et al. 1874-1959, pl. 177, figs. 45-50
Cyclotella hannaeKanaya (1957), pp. 82-4, pl. 3, figs. 10-14.
Lysigonium architecturale (Brun) Kuntze 1898: 412
Descripción: Valva de 8-10 micrones de altura. Valvas de 10 a 40 micrones de diámetro. En la parte frontal de la valva, areolas en filas radiales regulares, aproximadamente 14 areolas en 10 µm, se rompen de manera desigual cerca de un gran campo central, ocupando aproximadamente la mitad del diámetro de la valva. Campo con pequeños tubérculos o espinas, que son paredes cubiertas de aréolas modificadas, a veces hay aréolas únicas entre los tubérculos. En la curva, filas verticales de 2 areolas, 7-8 en 10 µm, sus filas transversales forman 2 anillos regulares. Los tabiques verticales del anillo superior de las areolas terminan en espinas conectoras marginales cónicas cortas, de aproximadamente 1 µm, 7-10 en 10 µm, formando un anillo exterior en el borde del pliegue y el frente de la valva; en los huecos entre ellos, el anillo de borde de perforaciones redondas.
Las espinas conectoras del anillo interior son más gruesas, romas, de planta irregularmente elíptica, de aproximadamente 1 µm de altura, ubicadas en un anillo marginal estrecho y sin estructura, aproximadamente 10 espinas en 10 µm. Los pines de ambos anillos están escalonados. Las excrecencias de dos labios son muy pequeñas y se abren dentro de la valva con una hendidura bordeada paralela al borde de la valva (Makarova et al. 1992).
Distribución Estratigráfica: Esta especie es característica del Eoceno Inferior-Medio, con especímenes encontrados en el Eoceno Superior, Oligoceno Inferior-Superior (Makarova et al. 1992).
La bioestratigrafía está vinculada a la escala de tiempo de polaridad geomagnética utilizada por la Expedición 320/321 (Expedition 320/321 Scientists 2010; Westerhold et al. 2012).
El taxón determinado en este trabajo (Tabla 2) se corresponde con la especie descrita por Fenner et al. (1978); Fenner (1985); Gladenkov y Barron (1995), Makarova et al. (1992).

CONCLUSIONES
Se reporta por vez primera para la Formación Senado en Cuba Central, la especie de diatomea marina Distephanosira architecturalis (Brun) Gleser.
La presente determinación micropaleontológica constituye un nuevo aporte al conocimiento bioestratigráfico, representada en la Formación Senado y permite considerar la parte inferior de dicho corte con una distribución estratigráfica desde el Eoceno Medio. La especie Distephanosira architecturalis (Brun) Gleser es encontrada en latitudes bajas desde el Eoceno Inferior hasta el Oligoceno Inferior-Superior.

