










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
En los mamíferos se conoce que las condiciones de desarrollo en el momento de la concepción, durante la gestación y el período neonatal afectan la salud posterior al nacimiento a largo plazo. Este fenómeno, se asocia con modificaciones en la expresión génica debido a mecanismos epigenéticos inducidos por el medio ambiente (Robles y Chavatte-Palmer, 2017).
En Cuba, los términos “epigenética” y “programación fetal” se han desarrollado escasamente en el campo de la ganadería, de ahí la necesidad de informarlos por su importancia en la producción animal, en el peso al nacer y al destete, rasgos que son por los que se evalúan los animales jóvenes.
DESARROLLO
La eficiencia de la especie bubalina en el país, se deprime paulatinamente, el énfasis en los análisis técnicos, se realiza sobre los rasgos que indican el comienzo de la vida reproductiva de hembras y machos, por lo que erróneamente no se consideran dos etapas previas trascendentales: 1) el estado de la búfala al momento de la gestación y durante esta; 2) los períodos posteriores al nacimiento de los bucerros, que son los responsables de que alcancen tempranamente la pubertad, madurez sexual y la edad al sacrificio de los machos.
En este artículo se referirá el concepto de programación fetal y los resultados obtenidos en Cuba en el peso al nacer y al destete, por la importancia que representan para la estabilidad de la cría de búfalos y el aporte de alimentos a la población. La información científica sobre el tema y su impacto en las diferentes funciones de humanos y animales es muy amplia (Kiefer y Perrier, 2019; Takeda et al., 2019; Mi et al., 2021; Breton et al., 2021 López Aguilar et al.,2020 ), no obstante, se reseñan algunos temas.
Epigenética, programación fetal o del desarrollo
La epigenética hace referencia al estudio de los factores que, sin corresponder a elementos de la genética clásica, interaccionan en la expresión de los genes. Estos elementos determinados por factores ambientales y genotipos subyacentes, influyen en la expresión fenotípica del ganado (producción de leche, calidad de la lana, resistencia a enfermedades, crecimiento y desarrollo, etc.), intervienen en la determinación de la ontogenia o desarrollo de un organismo, desde la fecundación del cigoto en la reproducción sexual hasta su senescencia y en la regulación heredable de la expresión génica sin cambio en la secuencia de nucleótidos (Lacal y Ventura, 2018; Skvortsova Iovino y Bogdanović, 2018; Aristizábal et al., 2019; Beaujean et al., 2020; Wu y Sirard, 2020; Breton et al., 2021; Ibeagha-Awemu y Yu, 2021; Peral-Sanchez et al., 2022), la Figura 1 muestra de manera esquemática los factores epigenéticos en la producción animal. La epigenética intergeneracional representa la transmisión de las marcas epigenéticas de una generación a la próxima (Pang et al., 2017).
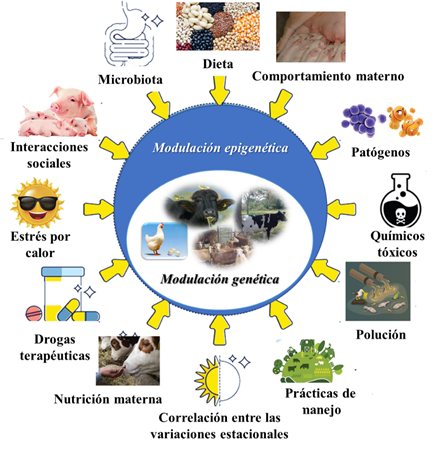
Fig 1 Factores epigenéticos que responden a factores ambientales externos e internos e interactúan con el genotipo subyacente para influir en los resultados del fenotipo (Ibeagha-Awemu y Yu, 2021).
Según Anastasiadi et al. (2021), las consecuencias de la variación epigenómica varían según las fuentes (intrínseca, genética, extrínseca), la previsibilidad ambiental es un factor clave para determinar las consecuencias de la herencia epigenética sobre el fenotipo y la aptitud, finalmente consideran que la herencia epigenética es otra pieza del rompecabezas de la herencia no genética, aunque la prevalencia, las fuentes, la persistencia y las consecuencias fenotípicas de las marcas epigenéticas hereditarias en los taxones siguen sin estar claras.
Como parte de los efectos epigenéticos surgió el término “programación del desarrollo o programación fetal”, desarrollado por el Dr. David Barker en Inglaterra y se sustentó en datos epidemiológicos vinculados a los efectos de la desnutrición prenatal sobre la salud humana a largo plazo (Khanal y Nielsen, 2017; Reynolds et al., 2019).
Se denomina programación fetal al conjunto de estímulos recibidos durante la vida intrauterina, capaces de interferir en el desarrollo fetal y tener consecuencias permanentes en la vida adulta (Abruzeese et al., 2018; Lunesu et al., 2020), es decir la exposición a determinados factores en este periodo de susceptibilidad, puede modular los procesos del desarrollo vinculados en la formación del fenotipo del adulto. Debido a su alcance, comenzaron los trabajos en animales domésticos, con la finalidad de observar cuales eran los efectos de la nutrición durante etapas tempranas, intermedias y finales de la gestación sobre las crías y sus implicaciones a largo plazo sobre los parámetros productivos, reproductivos e inmunológicos (Torres Diez, 2018), así como los cambios provocados por el estrés por calor (Wu, Zhang y Li, 2020).
Si bien, no se encontraron referencias en relación a este tema en búfalos de agua específicamente, varios autores que enfocaron sus pesquisas, en las etapas inmediatas a la concepción en otros rumiantes, demostraron que la programación del desarrollo, al menos en términos de efectos nutricionales maternos, comienza desde muy temprano en la gestación (Crouse et al., 2017; McLean et al., 2017).
El impacto de la desnutrición prenatal esta modulado por el momento de la gestación en el cual ocurre, la intensidad de la restricción nutricional y la duración del periodo de restricción. En la primera mitad de la gestación los requerimientos nutricionales del feto son relativamente bajos, es la etapa en la cual ocurre el máximo crecimiento, desarrollo y vascularización de la placenta y la organogénesis en el feto, lo cual resulta crítico para el normal crecimiento y desarrollo durante toda la vida del individuo (Reynolds y Vonnahme, 2016).
Las crías que reciben una restricción nutricional durante la primera mitad de la gestación, aun con sea apropiada hasta el final de ese período, pueden nacer con pesos normales, sin embargo, son portadoras de muchas de las características fenotípicas de congéneres que sufrieren restricción nutricional a lo largo de todo el periodo de gestación, dentro de estas se encuentran rasgos económicamente importantes, como mortalidad perinatal, disfunción de órganos esenciales, disminuir el crecimiento postnatal y depreciación de la calidad del animal y de la carne (Greenwood Calyton y Bell, 2017).
El potencial de crecimiento muscular se basa en la formación adecuada de esas durante el período prenatal, ya que no hay aumento en el número de ellas posterior al nacimiento, en relación con la cantidad de fibras musculares, Costa et al. (2021a), expresan que en hembras rumiantes gestantes criadas en pastos, la restricción alimenticia debido a la estacionalidad del forraje puede afectar el desarrollo de la descendencia, en esta situación, los nutrientes de la madre se priorizan para la formación de tejidos vitales fetales en lugar de tejidos secundarios, como el músculo esquelético, reduciendo eficiencia en la producción de carne. El deterioro causado por la restricción proteica en vacas para la producción de carne durante la mitad de la gestación, implican una reducción duradera del número de fibras musculares en la descendencia, además de alterar en mayor proporción el metabolismo muscular de fibra de tipo glucolítico en una etapa temprana de la vida, que puede revertirse dependiendo del ambiente postnatal.
De manera similar, la restricción energética durante la última etapa de la gestación contribuye a la baja regulación de los genes implicados en el metabolismo oxidativo, y consecuentemente favorece la baja eficiencia del metabolismo glucolítico en el músculo esquelético de los terneros (Sanglard et al., 2018), sin embargo, en cabritos recién nacidos hijos de madres con restricción en su alimentación en diferentes estadios de la gestación, no se observaron diferencias fenotípicas en ese músculo (Costa et al., 2019).
Al respecto, Costa, Gionbelli y Duarte (2021b) refirieron que la manipulación nutricional durante el período intrauterino contribuye a lograr características deseables de la calidad de la carne, como el marmoleado y la terneza.
La nutrición insuficiente o disponibilidad excesiva de nutrientes también alteran el estado epigenético de las células germinales in vivo e in vitro y transmisión a las generaciones posteriores (Guo, Luo y Lin, 2020), se disponen estudios que asocian ambas condiciones previas al apareamiento a una calidad deficiente de los ovocitos, menores tasas de fecundación y desarrollo embrionario, las crías podrían presentar menor recuento de folículos antrales (Mossa et al., 2015), incremento de la edad al primer parto, mayor número de servicios por concepción, menores ganancias de peso, distocias e infertilidad, debido al lento desarrollo fisiológico comparado con los animales cuyas madres no padecieron de la falta de nutrientes.
Las vacas lecheras, que están perdiendo peso en el momento de la concepción, generan embriones y crías comprometidas que traen una marca metabólica única. Un fenómeno similar se ha asociado con diferentes condiciones de cultivo y el procedimiento de fertilización in vitro. Como resultado de lo anterior el embrión se comporta en modo "económico" donde se reducen la traducción, la división celular y la producción de adenosín trifosfato, para adaptarse al entorno percibido. Se han realizado pocos estudios epidemiológicos en bovinos para evaluar si estos cambios resultan en un fenotipo diferente y se requieren más estudios para asociar cambios moleculares específicos en embriones con consecuencias visibles más adelante en la vida (Sirard, 2021), igual sucede con la vulnerabilidad del epigenoma a cambios drásticos de temperatura durante el período de preimplantación (de Barros y Paula-Lopes, 2018). Donkin y Barrès, (2018), expresaron que al igual que las células somáticas, el epigenoma de los espermatozoides es dinámicamente reactivo a una amplia variedad de factores estresantes ambientales.
En relación con la producción de leche, al observar el desempeño productivo durante la primera lactancia de terneras cuyas madres se encontraban en producción durante la confirmación de la gestación, hubo una tendencia hacia la disminución de la producción total durante la primera lactancia (18 kg) y menor duración de la lactancia (23 días), en contraste a que aquellos animales que fueron concebidos por animales que no estaban lactando, esto pudo deberse a que los requerimientos nutricionales de las vacas en gestación y producción no fueron completamente cubiertos (González-Recio, Ugarte y Bach, 2012).
Peso al nacer (PN)
Se reconoce que el peso al nacer refleja el nivel nutricional de la madre durante la gestación, pero que la programación fetal puede ocurrir independientemente del peso al nacer (Reynolds y Caton, 2012; Reynolds et al., 2019). En Cuba, los bucerros mestizos de Buffalypso x Carabao, pesaron entre 37,9 a 42,2 kg con alimentación a base de pastos, miel-urea y alimentos concentrados (García et al., 2003). En la provincia Pinar del Río, la media general fue 37,8±0,03 kg (Fraga, González y Gutiérrez, 2006), superior a la que informaron Martínez et al. (2009) en Granma, con 36,82±3,1 kg y en la empresa Maraguán de Camagüey, cuyo promedio fue 32,0±0,2 kg (Ceró et al., 2017). Los resultados coinciden con los de Brasil, los bucerros de las razas Jaffarabadi, Mediterráneos y Murrah tuvieron pesos al nacer de 36,43; 39,79 y 37,63 kg respectivamente (Mendes et al., 2017).
Peso al destete (PD) y ganancia media diaria (GMD)
En los rebaños en que las búfalas no se ordeñan y las crías tienen acceso a una mayor cantidad de leche, el estado nutricional es mejor, se obtienen animales destetados con mayor peso, que alcanzan rápidamente la madurez sexual o el peso de sacrificio.
Cuando se va a implementar el sistema de cría artificial en la actividad bubalina, hay que tener en consideración que los bucerros tienen un fuerte instinto maternal, las relaciones entre madre e hijo se desarrollan a partir del nacimiento, además exhiben un período más lento de aprendizaje, cuando se comparan con los terneros cruzados, esto implica mayor tiempo para aprender a beber la leche de manera artificial, por tanto, la separación de las madres y sus crías en los búfalos, resulta más estresante para ambos, cuando se comparan con los teneros Bos taurus, aunque no afecta, de manera significativa, la salud y el sistema inmune de los bucerros (Bharti et al., 2015).
Kumar et al. (2017), señalaron que en dos grupos de bucerros Murrah, amamantados por sus madres (G1) y en crianza artificial inmediatamente después del nacimiento (G2), aunque todos los bucerros nacieron con un peso corporal similar al nacer, el peso a los 180 días fue significativamente inferior en el grupo G2 con respecto a los del G1 (113,12±2,81 vs 122,77±2,63 kg respectivamente; p˂0,05), de ahí que se reconoce el amamantamiento natural tiene un efecto positivo en el aumento de peso corporal de los bucerros.
Las características de la cría bubalina, en todo el país, en los años siguientes, con insuficiencias en el manejo en la etapa presdestete, bajos niveles nutricionales de las madres, pastos naturales con deficientes manejos, estancamiento de los flujos zootécnicos y el cumplimiento a priori de los planes de producción lecheros, que provocan violaciones en la garantía de la alimentación de las crías y en el momento de secar a las búfalas, han atentado contra el peso al destete de las crías.
Los PD referidos en Cuba son bajos: 122,0±0,3 kg en 255 bucerros de cuatro lecherías de la provincia Granma (Méndez y Fraga, 2010) y en la empresa Maraguán Ceró, Guevara y Cáceres (2013) expresaron que en 768 crías destetadas entre 2006 a 2010, el peso promedio fue de 107,2±1,35 kg, resultados inferiores a los expresados en la literatura de Latinoamérica. En un rebaño lechero Murrah de Brasil, con animales que pastaban en áreas cubiertas con Brachiaria brizantha cv. Marandu, suplementados con ensilaje de maíz o sorgo, el PD a los 8 meses fue de 170,30±24,12 (Andrade, 2011), en búfalos criados en diferentes unidades productivas de Colombia, con edades de 270±19 días, fue de 182±42,46 kg (Bolívar et al., 2012).
En una empresa de la provincia de Camagüey, la GMD promedio al destete para las hembras fue de 407 g/día y en los machos de 419 g/día. Según el número de lactancia de las búfalas, los bucerros hijos de madres de primer parto alcanzaron 369 g/día y las de dos o más partos entre 394 a 446 g/día (Ceró et al., 2017). En estas condiciones es difícil que las hembras obtengan los 400 kg de peso vivo para su incorporación a la reproducción a los dos años de edad, como se estableció en Cuba en el año 2009, al establecer los indicadores zootécnicos mínimos para los rebaños bubalinos, para alcanzar este propósito deben ganar como promedio, entre 550 y 600 kg/día. Con los resultados expuestos, se infiere que hembras y machos a los 24 meses, estarán por debajo de lo óptimo para comenzar sus ciclos reproductivos. El destete con 170 kg de peso a los nueve meses de edad, con madres en ordeño, garantizan animales con 440 kg de peso en 24 meses, y para aquellos que se crían a toda leche, alcanzan un peso relativo de 480 kg en 20 meses.
CONCLUSIONES
Los elementos planteados en la literatura indican que las afectaciones de las futuras crías, comienzan desde el momento de su concepción, que los cuidados deben empezar con la hembra que se va a gestar, que resulta muy conveniente realizar análisis técnicos a partir del conocimiento de la condición corporal de la futura madre, que promover un ambiente uterino óptimo no solo asegurará el desarrollo fetal exitoso, sino que mejorará el rendimiento del crecimiento postnatal y reducirá el riesgo de aparición de enfermedades en los animales adultos y que finalmente, proveer buenas condiciones ambientales, con el objetivo de que las búfalas dispongan del bienestar necesario, deben ser prácticas a incorporar en la crianza de los búfalos de agua en Cuba.
Con la información publicada sobre los pesos al nacer y al destete, no se puede determinar que hubo efectos epigenéticos sobre los bucerros en el momento de la fecundación de sus madres, por lo que se recomienda incluir los temas sobre epigenética y dentro de esta la programación fetal, en futuras investigaciones en el ámbito pecuario, así como un tema en la asignatura Genética en la carrera de Medicina Veterinaria

