










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El consumo de fructooligosacáridos (FOS) proporciona beneficios a la salud de Seres humanos y animales. (1 La síntesis de estos prebióticos naturales ocurre por vía enzimática a partir de la sacarosa. El primer ciclo de fructosilación puede originar 1-kestosa (αglu [1,2] βfru [1,2] βfru), 6-kestosa (αglu [1,2]βfru [6,2] βfru) o neokestosa (βfru [2,6] αglu [1,2] βfru), en dependencia del tipo y el origen de la enzima empleada en la catálisis. El trisacárido puede extenderse mediante reacciones sucesivas de transfructosilación. La ingestión de 1-kestosa, 6-kestosa o neokestosa estimula de forma diferencial distintas especies bifidobacterias y lactobacilos que residen en el tracto intestinal.2),(3
Las compañías productoras de FOS realizan la conversión del azúcar caña o remolacha con catalizadores propios, no disponibles en el mercado, constituidos por β-fructofuranosidasas (EC 3.2.1.26) o fructosiltransferasas (EC 2.4.1.100) de hongos de los géneros Aspergillus o Aureobasidium. Estas enzimas sintetizan exclusivamente FOS lineales de enlaces β (2→1). La proporción porcentual de tri- (1-kestosa), tetra- (nistosa) y penta-sacárido (fructosilnistosa) es aproximadamente 37:53:10 al momento de máximo rendimiento de FOS totales (de 55 % a 60 %, p/p). Actualmente, no existe disponibilidad comercial de FOS de enlaces β (2→6) debido a la ausencia de catalizadores enzimáticos apropiados.4
La enzima β-fructosidasa (BfrA, EC 3.2.1.26) de la bacteria hipertermófila Thermotoga maritima se distingue por lograr la inversión total de la sacarosa a alta concentración de 600 g/L a 800 g/L y elevada temperatura de 60 °C a 70 °C. En la etapa inicial de la reacción se sintetizan bajos niveles de una mezcla de FOS.5),(6 La disponibilidad de la estructura 3D de BfrA y la identificación de los aminoácidos del sitio activo que interactúan con el ligando sacarosa7),(8 hacen posible establecer un diseño racional de mutaciones a realizar en el gen a fin de incrementar la relación de las actividades transferasa/hidrolasa de la enzima. El aumento de la reacción de transfructosilación (molécula aceptora del residuo fructosilo: sacarosa o fructano) en detrimento de la reacción de hidrólisis (molécula aceptora del residuo fructosilo: agua) resulta en un mayor rendimiento de FOS.
La enzima sacarosa: sacarosa 1-fructosiltransferasa (1-SST, EC 2.4.1.99) inicia la síntesis de fructanos en plantas. El principal producto de la reacción de 1-SST con la sacarosa es 1-kestosa. Las fructosiltransferasas vegetales, incluida 1-SST, son glicoproteínas que evolucionaron a partir de invertasas vacuolares mediante mutaciones puntuales que favorecieron la reacción de síntesis de FOS unido a la disminución o pérdida de la actividad hidrolítica ancestral.9),(10
La producción con fines biotecnológicos de las enzimas BfrA y 1-SST a partir de fuentes naturales no es una opción técnica ni económicamente factible. Como opción de hospedero recombinante de interés biotecnológico se destaca la levadura Pichia pastoris porque presenta estatus GRAS (generally regarded as safe), carece de actividad sacarolítica endógena y tiene la capacidad de secretar y glicosilar eficientemente proteínas foráneas.11 BfrA y 1-SST son enzimas con aplicación en la industria alimentaria. Se recomienda, por tanto, explorar el uso de un sistema de expresión constitutiva en lugar del sistema tradicional de expresión inducida por metanol, compuesto inflamable y tóxico a la salud.
Atendiendo a estos antecedentes y a la necesidad de desarrollar catalizadores enzimáticos novedosos que conviertan eficientemente la sacarosa en FOS de cadenas cortas y diferentes tipos de enlaces, se establecieron los siguientes objetivos de trabajo: obtener y expresar en Pichia pastoris mutantes de una β-fructosidasa bacteriana con aumento en la relación de las actividades fructosilación/hidrólisis y que rinda mayoritariamente FOS enlazados β(2→6); obtener un clon de P. pastoris que secrete altos niveles de una fructosiltransferasa vegetal especializada en la síntesis de 1-kestosa; obtener preparados enzimáticos de alta actividad y estabilidad destinado a la producción comercial de FOS de cadenas cortas.
Métodos
Microorganismos y medios de cultivo
Las cepas X-33 (tipo salvaje) y GS115 (auxotrófica a His4) de Pichia pastoris (Invitrogen) se utilizaron como los huéspedes de expresión.
Pichia pastoris se creció a 30 °C en los medios YPG (1 % [p/v( extracto de levadura, 2 % [p/v( peptona, 2 % glicerol [v/v () o YNB-G (1,34 % [p/v( base nitrogenada de levadura sin aminoácidos, 2 % [p/v( glucosa) suplementado o no con L-histidina (100 g/mL).
El medio de la fermentación en fase discontinua consistió en 5 % (v/v) glicerol, 0,5 % (p/v) levadura extracto, 2,2 % (p/v) (NH4)2SO4, 1,82 % (p/v) K2HPO4, 0,75 % (p/v) MgSO4 y 0,05 % (p/v) CaCl2 con vitaminas y trazas preparado según lo recomendado por Cregg y col. (1987). Para el gen constitutivo en la expresión el medio de alimentación consistió en un 50 % (v/v) glicerol. Cuando fue necesario el glicerol se reemplazó por azúcar de caña al 5 % y 50 % (p/v) en el medio de alimentación.
Obtención de variantes mutadas de la enzima β-fructosidasa (BfrA) de Thermotoga marítima y expresión en Pichia pastoris
Reacciones en cadena de la polimerasa para crear las sustituciones dirigidas al sitio W14Y, W14Y-N16S y W14Y-W256Y se realizaron con ADN de Pfu polimerasa (Promega, EE. UU.) se utilizó como molde el plásmido pALS263 que contiene el gen de la β-fructosidasa (BfrA) de Thermotoga maritima MSB8 (GenBank Accession No. AJ001073) con secuencia de codones optimizada para la expresión en Pichia pastoris.6 Los productos de PCR digeridos con EcoRI-KpnI (87 pb) sustituyendo uno (W14Y) o dos tripletes (W14Y-N16S) fueron usados para reemplazar el fragmento EcoRI-KpnI correspondiente en el plásmido pALS263 para construir los plásmidos pALS266 y pALS267, respectivamente. El producto de PCR digerido KpnI-PstI (689 pb) que crea la mutación W256Y se utilizó para reemplazar el KpnI-PstI correspondiente al fragmento en el plásmido pALS266 para construir el plásmido pALS268. La secuenciación de ADN se usó para verificar que solo las mutaciones deseadas estuvieran presentes y que la fusión de BfrA a la secuencia del péptido, señal del factor α de Saccharomyces cerevisiae, y a las seis histidinas ocurrieron en el marco correcto en todas las construcciones obtenidas. El vector vacío pGAPZαC (control negativo) y los plásmidos pALS263, pALS266, pALS267 y pALS268 se linealizaron con AvrII y electroporaron en la P. pastoris cepa X-33 y los transformantes fueron seleccionados en placas YPG con zeocina (100 μg mL−1).
Expresión de múltiples copias del gen 1-SST de Schedonorus arundinaceus en P. pastoris
Un fragmento de ADN de 1668 pb que codifica la región madura de 548-aa de 1-SST de Schedonorus arundinaceus se amplificó por PCR utilizando un ADN complementario de longitud completa10 como plantilla y los cebadores 5′GGGGCGAATTCCGCCGACGGCGGGTTC y 5′TGGTTCTAGATACAAGTCGTCGTTCGTG con sustituciones de bases para crear los sitios de flanqueo EcoRI y XbaI. El producto amplificado (Sa1-SST sin codón de parada) se digirió EcoRI-XbaI y se insertó en los sitios correspondientes del vector de expresión pGAPZαC (Invitrogen, Carlsbad, CA, USA) para dar lugar al plásmido pALS214. Para la multimerización in vitro del casete de expresión constitutiva de la Sa1-SST, el fragmento BglII-BamHI de 2,82 kb del plásmido pALS214 se sometió a ciclos repetitivos de cortes y autoligado. Multímetros de ~17 kb se recuperaron de geles de agarosa e insertaron en el sitio BamHI del vector pAO815 resistente a la ampicilina (Invitrogen, Carlsbad, CA, USA).
El plásmido resultante pALS227 se confirmó que contiene seis copias continuas del casete de expresión Sa1-SST y el gen His4 flanqueados por las regiones 5'AOX1 y 3'AOX1 por eventos de doble recombinación homóloga. Los clones de una copia correspondiente al plásmido pALS214 linearizado con AvrII fueron electroporados en la cepa salvaje. X-33 y los transformantes se seleccionaron en placas YPG con zeocina (100 μg/mL). Para los clones constitutivos de seis copias, el fragmento BglII de 22,3 kb del plásmido pALS227 se electroporó en la cepa GS115 auxotrófica a his4 y los transformantes se seleccionaron en placas YNB-G sin suplementación con histidina. La electroporación celular se realizó según lo descrito en el manual de expresión de Pichia (Invitrogen-Thermo Fisher Scientific). Mediante ensayos de retransformación de la levadura recombinante se obtuvieron clones con hasta 9 copias del transgén 1-SST.
Cultivo de Pichia pastoris en fermentador con incremento de fuente de carbono
El crecimiento de los clones de levadura recombinante se realizó en un fermentador INFORS HT de 7,5 L (5 L de volumen de trabajo) con 3,5 L de medio de fermentación con control de temperatura (30 °C) y pH (5,5). El valor de pH se ajustó mediante la adición de amoníaco o ácido fosfórico, según la necesidad. El oxígeno disuelto en el cultivo se mantuvo alrededor del 20 % por variación automática de la velocidad de agitación (de 500 rpm a 900 rpm) y flujo de aire (de 1 vvm a 2 vvm). En la fase de cultivo incrementado se añadió 1,5 L del medio (50 % [p/v] de azúcar de caña y 0,5 % [p/v] de extracto de levadura) a un flujo constante creciente en el intervalo o de 5,4 mL/h a 8,2 mL/h, por litro. Las condiciones de operación durante la fase de cultivo incrementado fueron 30 °C, pH 5,5, agitación 900 rpm y la aireación de 2 vvm. El tiempo total de fermentación fue de 3 d.
Ensayos enzimáticos
Una unidad de actividad de BfrA se definió como la cantidad de enzima requerida para liberar 1 μmol de glucosa por minuto a velocidad inicial de la reacción con 120 mM sacarosa, tampón de acetato de sodio 100 mM (pH 5,5) a 60 °C. La actividad global β-fructosidasa de las variantes se estimaron como la cinética de la liberación de glucosa, que es concomitante en las reacciones tanto con la hidrolítica (liberación de fructosa) como con la transfructosilación (síntesis de FOS). Una unidad de Sa1-SSTrec se define como la cantidad de enzima requerida para liberar 1 μmol de glucosa por minuto en la velocidad inicial de la reacción con sacarosa 1,46 M (500 g/L), 100 mM acetato de sodio (pH 5,5) a 30 °C. Los azúcares reductores en las mezclas de reacción se cuantificaron mediante el método colorimétrico del ácido dinitrosalicílico (DNSA) mediante calibración curvas de glucosa (para sustrato sacarosa) o fructosa (para sustratos rafinosa y fructanos).
Análisis de carbohidratos
Los carbohidratos en las mezclas de reacción de 1-SST fueron analizados por cromatografía líquida de alta eficacia (HPLC) utilizando una columna Aminex HPX-42C (0,78 × 30) cm (Bio-Rad, CA, EE. UU.) equipada con un detector de índice refractivo. La temperatura de la columna se mantuvo a 85 ºC. Se usó agua como fase móvil a un flujo de 0,5 mL/min. Las muestras (20 μL) se diluyeron apropiadamente antes de la inyección. Soluciones con cantidades definidas de fructosa, glucosa, sacarosa, 1-kestosa, nistosa y fructosilnistosa se usaron como estándares. Una mezcla de fructosa, rafinosa y estaquiosa se usó como estándar alternativo cuando fue necesario. Los paquetes de programa BioCrom versión 3.0 desarrollado en CIGB (La Habana, Cuba) fueron utilizados para la cuantificación de azúcares.
El análisis de oligosacáridos obtenidos por las reacciones de la BfrA nativa y mutada se obtuvo por cromatografía de intercambio aniónico de alto rendimiento con detección amperométrica pulsada (HPAE-PAD), mediante un sistema Dionex DX-500 IC equipado con bomba GP50, detector electroquímico ED50, inyector automático U3000 y un Columna CarboPac PA200 (3 mm × 250 mm, número de producto 062896 de Thermo Scientific) mantenido a 30 °C. La elución del producto se realizó aplicando un gradiente de acetato de sodio con 100 mM NaOH a 0,5 mL min−1 como sigue: acetato de sodio 5-100 mM en 25 min, 100-400 mM en 60 min, y 10 min para reequilibrio (acetato de sodio 5 mM). Se emplearon curvas estándar de los FOS (1-kestosa, 6 - kestosa, neokestosa, 1,1-kestosa, 1,6-kestosa, 6,6-kestosa y 1,1,1-kestopentosa) para el análisis cuantitativo a través de HPAE-PAD. La glucosa, fructosa y sacarosa se cuantificaron por alto rendimiento cromatografía líquida (HPLC) usando un controlador Waters 600 E Plus sistema (Waters Corp. Milford, MA, EE. UU.) equipado con una refracción detector de índice (Waters 410) y una columna Prevail Carbohydrate ES (250 × 4,6 mm) a 30 °C. Los azúcares se eluyeron con una fase móvil que consiste en una mezcla de acetonitrilo y agua 70:30 (v/v) a un flujo rango de 1 mL min−1. Las muestras se diluyeron adecuadamente antes de la inyección.
Curso temporal de producción de FOS
El efecto de la concentración de sacarosa y la dosificación de FOS sobre la producción de la enzima se examinó en reacciones de curso temporal usando un crudo de la preparación de 1SST que consiste en sobrenadante de cultivo ultrafiltrado de clon PGFT6 x - 308. Todas las mezclas de reacción se prepararon en tampón de acetato de sodio 100 mM (pH 5,5). Las reacciones por lotes se llevaron a cabo en un recipiente de 1 L con agitación a 250 rpm o en un reactor de 7,5 L con agitación a 250 rpm. Las muestras 0,5 mL se extrajeron a intervalos de tiempo y se incubaron en agua hirviendo durante 5 min para detener la reacción antes de ser analizadas por HPLC.
Resultados y discusión
Construcción de mutantes BfrA y expresión constitutiva en Pichia pastoris
Un gen con el uso de codones optimizado que codifica para la proteína intacta β-fructosidasa (BfrA) de la bacteria hipertermófila Thermotoga maritima cepa MSB8 se utilizó como plantilla para reemplazar tres aminoácidos (W14, N16 y W256) estrictamente conservados en el sitio activo de enzimas sacarosa hidrolasas por los aminoácidos que ocupan posiciones equivalentes en enzimas vegetales con actividad fructosiltransferasa. La BfrA nativa y los mutantes construidos (W14Y, W14Y-N16S y W14Y-W256Y) se fusionaron al péptido, señal del factor α de Saccharomyces cerevisiae,y expresaron en Pichia pastoris bajo el control transcripcional del promotor fuerte y constitutivo GAP (gliceraldehído 3P deshidrogenasa). La expresión constitutiva de los tres mutantes o la BfrA nativa no causaron inhibición del crecimiento celular. En todos los casos, la levadura recombinante secretó una enzima termoestable activa. Se detectó actividad sacarasa tanto en el sobrenadante de cultivo de 74 % a 76 % como en la biomasa intacta de 24 % a 26 %.12
Incremento de la relación de las actividades fructosilación/hidrólisis en mutantes BfrA
Las velocidades iniciales de las actividades hidrolasa (VF) y transferasa (VG-F) de cada variante mutada se comparó con la de BfrA nativa en reacciones a alta concentración de sacarosa (1,75 M) y temperaturas de 50 °C, 60 °C y 70 °C. Los 3 mutantes (W14Y, W14Y-N16S y W14Y-W256Y) mostraron similar aumento de 3 a 4 veces en la relación de las actividades transferasa/hidrolasa con respecto a la enzima nativa, independientemente de la temperatura de reacción. En todos los casos la hidrólisis se mantuvo como la reacción predominante, con más del 80 % de residuos fructosilos transferidos al agua (Figura 1).12
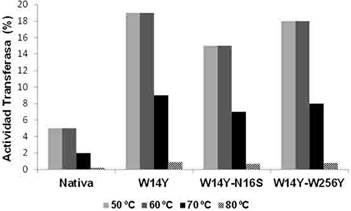
Rendimiento y composición de los productos FOS de BfrA nativa y las variantes mutadas
BfrA y las tres variantes mutadas produjeron una mezcla de FOS con composición mayoritaria de 6-kestosa (tabla 1). El mutante W14Y, de mayor actividad específica 841 U mg−1 y rendimiento global de FOS del 38 % (p/p), representa un candidato atractivo para la producción de FOS enlazados β(2→6) en reacciones a altas temperaturas de 50 °C a 70 °C.12 Del mismo modo, 6K y NK han sido informados como los principales FOS producidos por la β-fructosidasas de levaduras mesofílicas con predominante actividad de hidrólisis sobre la sacarosa.13 A pesar de las propiedades prebióticas de 6K, NK y 1K, el bajo rendimiento de FOS total de BfrA y las β-fructosidasas de levadura (menor del 16 %, p/p) hace a las enzimas de tipo salvaje comercialmente incompetentes para la conversión de sacarosa en fructanos de cadenas cortas.
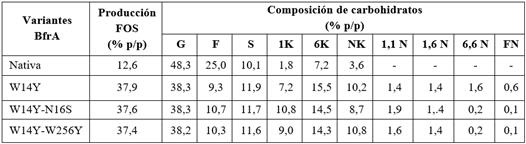
La enzima se reaccionó con 1,75 M con sacarosa, acetato de sodio (pH 5,5) a 60 ºC durante 22 h. Los productos cuantificados en la mezcla de reacción fueron: glucosa (G); fructosa (F); sacarosa (S); 1-kestosa (1 K); 6-kestosa (6 K); neokestosa (NK); 1,1-kestosa (1,1 N); 1,6-kestosa (1,6 N); 6,6-kestosa (6,6 N); 1,1,1-kestopentosa o fructosilnistosa (FN). La producción de FOS total contiene tri-, tetra- y pentasacáridos. Los valores representan las medias de n= 3. La desviación estándar fue inferior al 10 %.
Expresión constitutiva del gen 1-SST de Schedonorus arundinaceus en Pichia pastoris
El cDNA 1-SST de Schedonorus arundinaceus (Sa1-SST) codifica una proteína precursora de 654 aa con una secuencia N-terminal de 106 aa (pre- y propéptidos), que se escinde durante el tránsito a vacuola. La región madura (548 aa) de Sa1-SST se fusionó a un péptido, señal de levadura y se expresó en Pichia pastoris bajo el control del promotor constitutivo GAP. El crecimiento del clon simple copia (llamado PGFT1x) en un medio salino con glicerol mostró un rendimiento de biomasa de 102 g/L (peso seco) y la actividad de la enzima recombinante (Sa1-SSTrec) alcanzó 3955 U/L al final de las 72 h de cultivo. La productividad volumétrica fue 55 U/L/h. La expresión constitutiva del transgén no inhibió el crecimiento celular de la levadura.14
Aumento del rendimiento de Sa1-SSTrec mediante el incremento de la dosis génica y el empleo de sacarosa como fuente de carbono
Como estrategia para incrementar la tasa de transcripción del transgén, y con ello el rendimiento de la enzima, el casete de expresión pGAP-1SST se multimerizó in vitro. Multímeros de 3 y 6 casetes se clonaron en vectores con diferentes marcadores de selección, y se integraron vía recombinación homóloga a regiones definidas del genoma de P. pastoris. Mediante análisis de Southern blot y qPCR se determinó que los clones PGFT1x, PGFT6x y PGFT6x-308 portan 1, 6 y 9 copias intactas del transgén. El aumento de la dosis del transgén hasta 9 copias incrementó de forma proporcional el rendimiento y la secreción de la enzima recombinante. La expresión constitutiva del gen 1-SST le confirió a la levadura actividad sacarolítica. La sustitución del glicerol del medio de cultivo por azúcar de caña, una fuente de carbono más barata, incrementó el rendimiento de Sa1-SSTrec hasta 60 U/mL en el clon PGFT6x-308. La glucosa liberada por la acción de la enzima recombinante sobre la sacarosa del medio tuvo un efecto estimulador de la transcripción a partir del promotor GAP.14
Sa1-SSTrec convierte la sacarosa en FOS de cadenas cortas del tipo 1-kestosa y nistosa
Sa1-SSTrec se purificó por cromatografía de intercambio iónico del sobrenadante de cultivo del clon PGFT6x-308 como una glicoproteína activa y estable. Sa1-SSTrec no tiene actividad invertasa sobre la sacarosa, pero sí puede hidrolizar el producto 1-kestosa. El curso de la producción de FOS por Sa1-SSTrec bajo condiciones óptimas de operación se investigó en un reactor de tanque agitado de 7,5 L (Figura 2). La enzima produjo proporciones equimolares de glucosa y trisacárido (GF2) hasta agotarse del 65 al 70 % del sustrato inicial. A partir de este punto comienza la síntesis de tetrasacárido (GF3). La aparición de fructosa libre indica el momento adecuado de inactivar la enzima por calor y alcanzar el valor de FOS (GF2 + GF3) de ~360 g/L, de 55 % a 60 % (p/p) del contenido final de carbohidratos. En ese momento quedan por consumirse 100 g del sustrato sacarosa.14
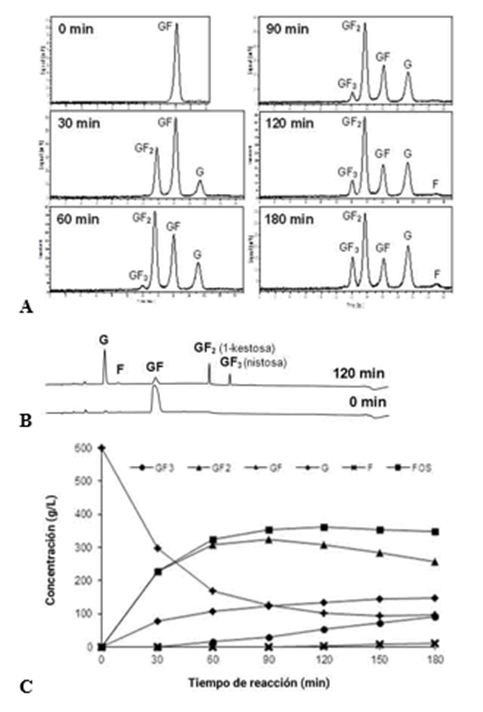
Fig. 2 Curso temporal de la reacción Sa1-SSTrec (9000 U/L) con azúcar de caña (600 g/L) a pH 5,5 en reactor con agitación constante a 250 rpm y temperatura 40 ºC. La composición de los carbohidratos de la mezcla de reacción fue determinada por cromatografía líquida de alta eficacia, HPLC (A) HPAEC-PAD (B). La desviación estándar de las medias (n= 3) en la curva de producto versus tiempo de reacción (C) estuvo pox zcr debajo del 10 %. Leyenda: glucosa (G); fructosa (F); sacarosa (GF); 1-kestosa (GF2); nistosa (GF3).
La caracterización estructural de los productos sintetizados por Sa1-SSTrec se realizó por HPAEC-PAD. El análisis del cromatograma de la mezcla de reacción, al tiempo de máximo rendimiento de FOS totales (120 min), reveló que GF2 y GF3 corresponden a 1-kestosa y nistosa, respectivamente con relación GF2:GF3 de 9:1. La ausencia de picos que pudieran asignarse a 6-kestosa o neokestosa evidencia que Sa1-SSTrec une los residuos de fructosa solo mediante enlaces β (2→1).
Rendimientos de FOS totales similares de 55 % a 60 %, (p/p) se obtienen mediante las fructosiltransferasas fúngicas más termoestables, pero con una proporción 3 veces menor de GF2 debido a un aumento del contenido de GF3 y la formación de GF4.4),(15),(16 La sacarosa es el sustrato donante y aceptor preferencial de Sa1-SSTrec. La síntesis de 1-kestosa como único trisacárido y la ausencia de transferencia de fructosilo al agua, incluso a una baja concentración de sacarosa de 50 mM a 100 mM, denota la alta especialización de Sa1-SST en su función biológica como iniciador de la síntesis de fructanos en una planta monocotiledónea perenne.
Obtención de preparados enzimáticos BfrA y 1-SST para fines comerciales
Cada levadura recombinante se creció en medio salino con azúcar de caña en fermentador, bajo condiciones de cultivo incrementado y parámetros controlados: pH 5,5, temperatura de 28 °C durante 72 h. Los sobrenadantes de cultivo con las enzimas BfrA y 1-SST se sometieron a un proceso simultáneo de concentración y diálisis en tampón de acetato de sodio 100 mM (pH 5,5) y posteriormente se liofilizaron. Los preparados enzimáticos en forma de polvo soluble en agua presentaron alta actividad específica (> 8000 U/g) y elevada pureza proteica (> 50 %), además se mantuvieron estable durante 1 año de almacenamiento a temperaturas de 4 ºC y 25 ºC.
Conclusiones
Las compañías productoras de FOS realizan la conversión de la sacarosa con catalizadores propios, no disponibles en el mercado, constituidos por enzimas de hongos. El presente trabajo describe la producción extracelular en la levadura Pichia pastoris de la enzima 1-SST de la planta Schedonorus arundinaceus y de variantes mutadas la enzima β-fructosidasa de la bacteria Thermotoga marítima. Las enzimas recombinantes presentadas en forma de polvo soluble en agua constituyen catalizadores estables útiles para la producción de distintos FOS de cadenas cortas, azucares prebióticos de alto valor comercial.

