










 Servicios personalizados
Servicios personalizados texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Zenaida macroura L. (paloma rabiche) es la especie del orden Columbiforme más abundante en Cuba. Es considerada una de las especies de caza más importantes del país, por lo que tradicionalmente ha sido de gran interés para los cazadores, tanto nacionales como los que visitan la isla. Este interés está relacionado con sus principales características, según Acosta y Ponce de León (2004) en Chamizo y García (2004): excelentes aptitudes cinegéticas, que se evidencian a través de un rápido y zigzagueante vuelo, sobre todo cuando descubre la presencia del cazador y carne de excelente calidad.
Es una especie muy abundante desde el Sur de Canadá y los Estados Unidos de América hasta el Sur de México y las Antillas, según Acosta y Ponce de León (2004) en Chamizo y García (2004). Esta paloma es frecuente en potreros y terrenos llanos asociados a parches de bosques, campos cultivados, zonas residenciales y hasta parques urbanos. García (2017), al estudiar el comportamiento de la especie en diferentes biotopos obtuvo que el hábitat más importante para esta lo constituyen los biotopos agrícolas con cobertura. En estos biotopos, a pesar de existir un claro predominio de las plantas herbáceas, que constituyen la base de su alimentación, también se encuentra un considerable número de especies arbóreas y arbustivas, las que se ubican en linderos que bordean los caminos rurales, las cercas perimetrales y árboles aislados en las áreas de potreros. Estos árboles le sirven a la especie como percha para el descanso, refugio y sustrato para la nidificación, contrario a su comportamiento en las áreas boscosas dado su pobre potencial de alimentación, así como la propia estructura de la vegetación.
Uno de los principales aspectos que deben considerarse en la gestión de los recursos cinegéticos es el conocimiento del estado actual de sanidad y vitalidad de los mismos, así como los factores que inciden en la dinámica poblacional de las especies. Según Peiró (2003), el análisis biosanitario de las poblaciones cinegéticas ha de ser considerado como una parte de su aprovechamiento sostenible.
Núñez et al., (2011) exponen que la evaluación de la carga parasitaria individual debería proporcionar información relevante acerca del estado de salud en las aves. Debido a que Z. macroura es una especie no solo con valor ecológico, sino también cinegético y por tanto económico y recreativo, es importante conocer su estado sanitario, así como los parásitos que se relacionan con ella.
A las enfermedades de las especies de vida silvestre no se ha dado suficiente importancia ecológica, sino hasta la década de 1990 Simonette (1995) en Di Mare (2003) y sobre todo con especies en vías de extinción (Cooper, 1989 en Di Mare, 2003). Existen tres grupos de parásitos que pueden afectar los Columbidos: los artrópodos (ácaros, garrapatas y piojos), protozoarios (parásitos unicelulares que viven dentro de los hospederos) y helmintos (gusanos redondos).
Dentro de estos grupos se encuentran representantes con diversos requerimientos microambientales que les permiten invadir áreas externas del cuerpo como plumas, pecho, ano; así como áreas internas a lo largo del tracto digestivo y el sistema sanguíneo (Di Mare, 2003).
De acuerdo con Schmidt y Roberts (1984), citados por Pazmiño (2007), la avifauna silvestre generalmente se encuentra infectada por varias especies de parásitos, considerando que la mayoría de las aves silvestres toleran su carga de parásitos adecuadamente; sin embargo, estos animales mueren cuando la infección es alta, a causa de los diversos estragos causados por estos.
Los ectoparásitos en las aves son aquellas especies que viven en la piel, pelo o pluma de estas. Mientras que para Podulka et al., (2004) los ectoparásitos son importantes en la adaptabilidad de las aves, aunque las presencias de algunos de ellos pueden tener poco efecto en las aves, una gran infestación o carga parasitaria puede deteriorar la salud de un ave, así como la reproducción de esta. Las aves son anfitrionas de muchas clases de parásitos "chupadores de sangre", especialmente las moscas del plumaje (familia Hippoboscidae), las pulgas, piojos, ácaros y garrapatas.
Estudiar el parasitismo en aves silvestres es de suma importancia, ya que puede ser un reservorio natural que de alguna manera puede afectar a las aves en producción (Medellín et al., 2019).
Los estudios existentes sobre las parasitosis en la Z. macroura son exiguos y fragmentados en diferentes regiones del mundo, destacándose los realizados en los EUA por Galloway y Palma (2008) y en México por Medellín et al., (2019). En Cuba, a partir de las fuentes consultadas se encontró que existe escasa información acerca de los ectoparásitos que afectan las poblaciones y la condición corporal de esta especie. Resulta pues una necesidad investigar en el país la incidencia de ectoparásitos en especies silvestres como Z. macroura para lograr un manejo adecuado de sus poblaciones.
El objetivo de este estudio estuvo dirigido a determinar los ectoparásitos presentes en la población de Zenaida macroura, así como su incidencia según las clases de edad, sexo y época del año.
Materiales y métodos
La investigación se realizó en el área de caza "Damují" del municipio de Rodas, provincia de Cienfuegos, Cuba (Figura 1), cuyo centro se localiza en las coordenadas geográficas 22°21,804' N y 80°33,430' W, abarcando un total de 645,8 ha. A efectos de la presente investigación se seleccionó esta zona porque reúne las características representativas de los biotopos más usados por la especie objeto de estudio en Cuba.
El clima de la zona es tropical poco húmedo de llanura, ampliamente difundido hacia la zona central y occidental de la provincia de Cienfuegos. Este está caracterizado por ser caliente, húmedo y lluvioso entre los meses de mayo a octubre y seco, poco húmedo y temperatura menor de noviembre a abril. La temperatura media anual oscila entre los 22-24 °C y la precipitación media anual es de 1000 a 1100 mm (Barcia y Castillo, 2015).
El relieve es llano en casi toda el área y con predominio de los suelos ferralítico rojo típico y fersialítico pardo rojizo típico. El uso mayoritario del suelo es el cultivo agrícola y la ganadería con una amplia red de linderos o setos que dividen estas áreas. Una gran parte del área está dedicada a los cultivos agrícolas de forma intensiva (Manihot esculenta, Ipomea batatas y Zea mays entre otros), donde predominan las rotaciones de cultivos menores con riego artificial durante todo el año, pero también presentan pastos y plantas herbáceas bordeando los canales de riego que se mantienen con agua la mayor parte del año. Existen áreas dedicadas a la ganadería en las cuales predominan los pastos, tanto naturales como artificiales (Andropogon annulatus, Panicum maximun, Eleusine indica), aunque en algunos casos aparecen áreas de potreros invadidas por el Dichostachys cynerea (marabú) y otras arvenses. La parte restante de la zona (63 ha) está ocupada por un bosque natural semicaducifolio (Samanea saman, Bursera simaruba, Guazuma tomentosa) sobre suelo calizo, que en algunos sitios presenta un aceptable grado de conservación y en otros aparecen áreas abiertas (claros) de cultivos abandonados.
Los trabajos de campo consistieron en la toma de muestras de los ejemplares cazados de esta Z. macroura durante todos los meses del año. La caza fue realizada al azar con escopeta de cartuchos en recorridos a pie que abarcaron toda el área y cada ejemplar abatido fue inmediatamente etiquetada siguiendo el orden de captura y colocado en una bolsa plástica individual sellada (con el cuidado de cerrar inmediatamente la bolsa para evitar escape de ectoparásitos), luego se procedió a revisar minuciosamente las plumas, alas, pecho, dorso, ano, patas, pico y cabeza, para detectar parásitos externos. Con una pinza fina se colectaron los parásitos y fueron colocados en un frasco con alcohol al 70 % hasta lograr su identificación. Una vez colectados los ectoparásitos en cada ejemplar se les efectuó el pesaje con una balanza de 0,01 g de precisión y la disección para conocer el sexo, clasificándolos luego según la clase de edad (adulto o pichón). Con estos resultados se procedió a valorar la incidencia de estos en la población estudiada mediante el índice de prevalencia.
El Índice de prevalencia (Ip) permite conocer cuántas aves de un grupo definido están enfermas en un determinado momento, por lo tanto, nos indica el peso o la abundancia del evento que soporta la población (Bush et al., 1997 en González et al., 2004a). Fue calculado por la siguiente fórmula (Ecuación 1).
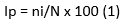
Donde:
ni |
= representa el número de palomas infestadas por la especie i de parásito |
N |
= representa el número total de palomas analizadas |
A los datos obtenidos en un primer momento se aplicaron técnicas de estadística descriptiva, con la finalidad de resumirlos y facilitar su interpretación (medidas de tendencia central, cálculo de porcentajes por clases de edad y sexos). Luego se les realizó un análisis exploratorio de los datos a través de los test de normalidad Kolmogorov-Smirnov, con el fin de conocer el comportamiento de la distribución y así determinar el tipo de análisis estadístico a utilizar (paramétrico o no paramétrico). Al realizar el análisis entre el peso total de los ejemplares parasitados y los no parasitados en las diferentes categorías, se aplicó el test de la T-Student según el comportamiento de normalidad en los datos obtenidos. Con el objetivo de comparar la media de la muestra obtenida con un valor de referencia de la especie en la literatura, se utilizó el test de T-Student (Martínez et al., 2009). En el procesamiento de la información se emplearon los programas Excel y SPSS versión 15.0 para Windows.
Resultados y discusión
Se estudiaron un total de 78 ejemplares. En el total de muestras colectadas solo se encontraron dos ectoparásitos: Lynchia americana, conocida como mosca de gavilán y un ácaro (aún sin clasificar) que se encontró en los tarsos de la mayoría de los ejemplares capturados. Para Núñez et al., (2011) el estudio del parasitismo en aves silvestres todavía tiene algunas limitaciones importantes. En primer lugar, el conocimiento taxonómico de los parásitos aviares es todavía muy incompleto.
De las 78 aves estudiadas, 26 (34 %) presentó alguna clase de ectoparásito, el grupo predominante fue el de los ácaros podales, los cuales constituyeron el 26 % de las aves parasitadas, seguidos por la Lynchia americana con el 8 % de la población parasitada.
Las moscas planas o del plumaje son un grupo de ectoparásitos de la familia Hippoboscidae que se alimentan de la sangre de las aves, son parecidas a las moscas domésticas, tienen el cuerpo sumamente aplastado, lo que le permite moverse con holgura entre el contorno de las plumas. Son conocidas también junto a otros ectoparásitos como los mosquitos, piojos y pulgas por ser trasmisores de enfermedades como la viruela.
Como se aprecia en la Figura 2, la Lynchia americana solo se encontró en los meses de marzo, mayo, junio y agosto, correspondiendo al mes de junio el mayor Índice de prevalencia (Ip) (37,5 %), coincidiendo con la época de máxima reproducción de la Z. macroura y el mayor número de pichones en la población. El Índice de prevalencia en adultos fue de un 4,7 % y en los pichones de un 21,4 %. Estos meses corresponden al período más lluvioso de nuestro país. Al analizar el comportamiento del Índice de prevalencia en ambos períodos se aprecia una marcada presencia de este ectoparásito en el más lluvioso (Figura 3).

Fig. 2 - Comportamiento del Índice de prevalencia de Lynchia americana en la población de Zenaida macroura a través del año
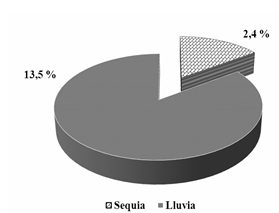
Fig. 3 - Comportamiento del Índice de prevalencia de Lynchia americana en período más lluvioso y período poco lluvioso en el área de estudio
Similar resultado reporta Di Mare (2003) en su estudio sobre Zenaida asiatica (paloma de alas blanca) en Tempisque, Costa Rica, donde reporta dentro de los ectoparásitos encontrados una Pseudolynchia canariensis (mosca hematófaga) en el plumaje de las palomas analizadas; aunque su incidencia en la muestra estudiada fue muy baja (1 % de índice de prevalencia). En cuanto al análisis de la prevalencia mensual de los ectoparásitos en Tempisque, Costa Rica, los meses de mayor afectación en la población correspondieron a los de la época de lluvias (marzo, julio, agosto y octubre). Leherman (1993) en Di Mare (2003) indica que una gran incidencia de ectoparásitos en la época de reproducción puede provocar el abandono de nidos activos en respuesta a una alta densidad de ectoparásitos.
En la literatura revisada, hasta el momento en Cuba, no se han desarrollado estudios de forma sistemática sobre el Índice de prevalencia de este ectoparásito en la población de Z. macroura, ni de ninguna otra especie de paloma del país, por lo que no se cuenta con parámetros que permitan identificar qué resultado de Índice de prevalencia en una población puede acarrear problemas de salud en esta.
Al analizar la relación de parasitismo por esta mosca entre ambos sexos en la población estudiada, se comprobó la no existencia de grandes diferencias en los resultados, mostrándose bajos estos en ambos sexos. Existió una ligera superioridad de los valores obtenidos en los machos con respecto a las hembras (Figura 4). Estos resultados registrados coinciden con los obtenidos por González et al., (2004) a en el estudio de parásitos gastrointestinales y externos de la Columba livia (paloma doméstica) en la ciudad de Chillan, Chile, donde comprobó que no hubo diferencias significativas entre la infestación por ectoparásitos entre los sexos. Según estos autores podría estar relacionado con el escaso dimorfismo sexual entre palomas, además de presentar hábitos similares en individuos de ambos sexos. No ocurrió igual en el estudio realizado en Ñuble, Chile por González et al. (2004)b al estudiar la fauna parasitaria de la Zenaida auriculata (tórtola común). Estos autores exponen que en la relación hembra-macho se puede observar una predominancia de las hembras con respecto a los machos, siendo esta más notoria en los ectoparásitos menos frecuentes Hohors-tiella sp. y Bonomiella sp.
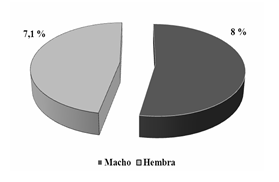
Fig. 4 - Comportamiento del Índice de prevalencia de Lynchia americana entre sexos en la población de Zenaida macroura
En la Figura 5, se puede observar el comportamiento del Índice de prevalencia en ambas clases de edad en la población de Z. macroura estudiada, destacándose la gran incidencia de este ectoparásito en las palomas jóvenes. Leherman (1993) en Di Mare (2003) plantea que los ectoparásitos chupadores como las moscas del plumaje, al chupar sangre producen heridas muy dolorosas, especialmente en los pichones jóvenes. Es, por tanto, esta incidencia un factor clave para el éxito reproductivo de la especie. Al respecto, Núñez et al., (2011) expuso que, en las aves, la condición física influye notablemente sobre el comportamiento petitorio de los pollos en el nido.
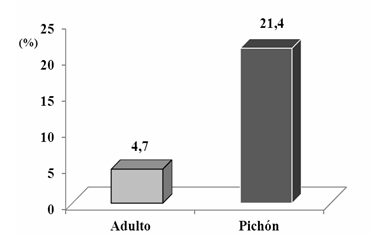
Fig. 5 - Comportamiento de Índice de prevalencia de Lynchia americana en las clases de edad en la población de Zenaida macroura
En la presente investigación es importante destacar que en las aves parasitadas por Lynchia americana no se detectaron cambios patológicos notorios al momento de las necropsias en las muestras estudiadas; ni al realizar la prueba de T de Student para comparar una media con un valor de referencia, escogido en este caso, los valores medios del peso corporal de las aves parasitadas vs peso promedio de la población del área de estudio (Tabla 1). El peso total de un ave, a pesar de ser una de las medidas más simples, es de gran importancia ya que además de ser de gran utilidad para conocer las necesidades energéticas de la especie, es un buen indicador de sus posibilidades de supervivencia, por ser un indicador de las reservas necesarias para procesos vitales como la migración, reproducción y muda (Acosta et al., 2013). Por tanto, valores por debajo de los medios o normales para una especie deben ser tomados como anomalía en algún aspecto de la calidad de hábitat o la salud de los individuos.
Tabla 1 - Resultados obtenidos al realizar la prueba de T-Student para comparar una media con un valor de referencia entre el peso promedio de la población de Zenaida macroura en el área de estudio vs el peso promedio de las aves parasitadas por Lynchia americana

A pesar de no existir diferencias estadísticas significativas, se puede observar superioridad en el peso promedio de las aves no infestadas por Lynchia americana, lo que está dado por la posible depauperación progresiva que pueden sufrir las aves infestadas debido a las enfermedades que pueden contraer producto de las picadas de estas moscas o a la pérdida constante de sangre por la alimentación de los ectoparásitos. Al respecto, plantean Núñez et al., (2011) que el análisis de la condición física a través de variables morfométricas es importante; este permite comprender como afectan los patógenos la población parasitada.
Entre los artrópodos parásitos más comunes en las aves que pueden afectar la condición física individual están los malófagos, que se alimentan normalmente de las plumas, la piel o los productos de la piel. Los ectoparásitos más frecuentes, sin embargo, son los ácaros (Núñez et al., 2011).
Estas parasitosis producidas por ácaros en palomas de diferentes géneros pueden variar en cuanto a su intensidad y especies de parásitos. Como resultado del análisis de los ejemplares de Z. macroura estudiados en nuestro caso, solo fue encontrado un ácaro parasitando las patas; este nivel de parasitosis encontrada en la población del área de caza Damují puede considerarse baja si es comparada con los resultados obtenidos por González et al., (2004)b al estudiar la fauna parasitaria de Zenaida auriculata (tórtola común) en Ñuble, Chile, donde se identificaron un total de ocho ácaros de diferentes géneros. En estudio realizado en la ciudad de Chillan, Chile, sobre la parasitosis en Columba livia (paloma doméstica) por González et al., (2004a), solo el uno por ciento de la muestra estaba afectada por un único ácaro (Dipaegidia columbae).
La Figura 6 permite apreciar el comportamiento del Índice de prevalencia en los ejemplares de paloma afectados con ácaros podales en relación con los meses del año. Así, este ectoparásito se encuentra prácticamente en ambos períodos del año; siendo marzo, abril y agosto los meses de mayor prevalencia, los que corresponden con el período más lluvioso. Los ácaros podales hallados no mostraron un patrón estacional definido. Si bien se observaron fluctuaciones en los Índices de prevalencia a lo largo del año, podría denominarse como una especie común en la comunidad.
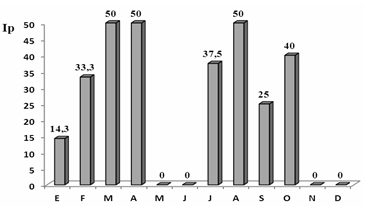
Fig. 6 - Comportamiento del Índice de prevalencia de ácaros podales en la población de Zenaida macroura a través del año
Los resultados expuestos anteriormente son confirmados con la relación obtenida al calcular el Índice de prevalencia en las dos épocas bien marcadas del país (período más lluvioso y período menos lluvioso), como se muestra en la Figura 7. Resulta evidente la poca diferencia entre los valores obtenidos.
Se considera importante mencionar que, en esta investigación, dentro de la población parasitada, las aves con mayor infestación (carga parasitaria por ejemplar) se encontraron durante los meses de marzo, septiembre y octubre, coincidiendo estos meses de alto índice con el período más lluvioso del país, caracterizado por altas temperaturas y abundantes precipitaciones.

Fig. 7 - Comportamiento del Índice de prevalencia de ácaros podales en dos épocas en el área de estudio
Al analizar el Índice de prevalencia de los ácaros podales entre sexos en la población parasitada de Z. macroura estudiada a través del año, se aprecia que ninguno superó el 50 % o más, lo que constituiría una clara preferencia sobre algún sexo. Vale destacar que los más altos grados de infestación por este ácaro en las aves parasitadas correspondieron a ejemplares machos (Figura 8).
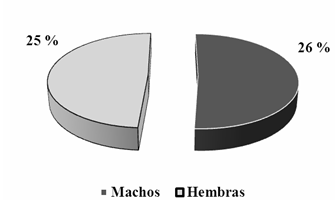
Fig. 8 - Comportamiento del Índice de prevalencia de los ácaros podales entre sexos en la población parasitada de Zenaida macroura
En la Figura 9, se pueden observar los resultados obtenidos al calcular el Índice de prevalencia del ataque de este ácaro en relación con las clases de edad, evidenciándose una mayor incidencia en la población joven. Estos resultados fueron similares a los obtenidos en este estudio, al analizar los ejemplares afectados por Lynchia americana, donde también los índices de infestación por este ectoparásito fueron superiores en la población joven. Constituye pues, una evidencia de la influencia de esta clase de edad (jóvenes) sobre la estructura de las comunidades parásitas en esta Columbidae.
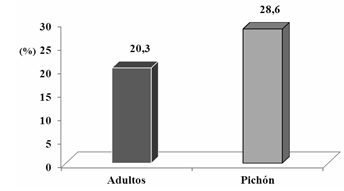
Fig. 9 - Comportamiento del Índice de prevalencia (Ip) de ácaros podales según la clase de edad en la población de Zenaida macroura
La diferencia observada entre ambas clases de edad, y coincidente con el comportamiento de Lynchia americana en la población estudiada, podría explicarse por el poco tiempo de vida que tienen los pichones, que pudiese conducir a una actitud diferente ante el ataque de los parásitos, con una respuesta más débil ante agentes patógenos.
Según Núñez et al., (2011), el sistema inmune de las aves consta de un componente innato y uno específico (o adquirido). El sistema innato responde de forma inmediata pero no específica a muchos agentes patógenos, representa una primera línea de defensa que no confiere inmunidad a largo plazo. La inmunidad adquirida, sin embargo, es capaz de producir una memoria inmunológica que proporciona una defensa mejorada en sucesivos encuentros con ese mismo patógeno. El sistema inmunitario innato está menos sujeto a variaciones ambientales, nutricionales o a los niveles de estrés que el sistema adquirido, y por lo tanto es un buen indicador de inmunocompetencia. Además, refleja la acción de los genes ligados a la producción de anticuerpos naturales, que están sujetos a selección natural.
Como ya se explicó anteriormente, en todas las aves examinadas se tuvo en cuenta la presencia de cambios patológicos notorios al momento de su examen, tanto en el físico como en las necropsias. En ninguno de los casos de ejemplares parasitados por ácaros podales fue detectada algún tipo de anomalía. Para conocer la relación entre el peso de las aves parasitadas por los ácaros podales y la media general de la población (Tabla 2) se realizó el test de T de Student (comparación de la media de una variable con distribución normal con un valor de referencia).
Tabla 2 - Resultados obtenidos al realizar la prueba de T-Student para comparar una media con un valor de referencia entre el peso promedio de la población de Zenaida macroura en el área de estudio vs el peso promedio de las aves parasitadas por ácaro podal
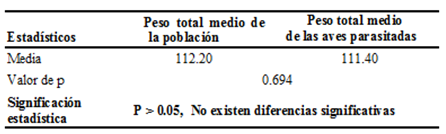
El resultado de esta prueba es similar al obtenido en el análisis de la Lynchia americana. La media de la población es superior a la de la población infestada y aunque no existan significación estadística se aprecia una ligera superioridad en el peso de la población sana, aspecto este a tener en cuenta. Según Núñez et al. (2011), el estado físico de un animal influye sobre su conducta y por lo tanto ha de considerarse en los estudios etológicos. Medir la condición física en una población puede también ser importante para evaluar los beneficios que aportan determinadas variables morfométricas y ayudar con ello a comprender su función adaptativa.
Una vez realizados los análisis de cada ectoparásito encontrado en la población de Z. macroura es indispensable contar con un análisis general de la incidencia de ambos en la población; de conjunto, ambos determinan el estado general de la salud poblacional. Es importante señalar que actualmente se desconoce hasta qué punto los parásitos estudiados afectan o limitan la población de la especie, así como los trastornos fisiológicos que provocan, por lo que se considera un tema a continuar investigando en el futuro.
Al comparar la incidencia de los dos ectoparásitos hallados en la población de Z. macroura se aprecia que la especie dominante resultó ser el ácaro podal, con valores del Índice de prevalencia más constantes en las diferentes épocas del año. A pesar de las variaciones observadas en este ácaro, la especie mantuvo la condición de común en todos los períodos climáticos analizados. Pocos cambios en la composición estacional pudiesen indicar una gran estabilidad tanto en el tiempo como en diversas características del hospedador.
Ninguna de las dos especies de ectoparásitos hallados superó el 55 % o más de Índice de prevalencia, lo que supondría una gran afectación en la población de paloma rabiche. No obstante, los ácaros podales constituyeron la especie con los mayores valores de Índice prevalencia (Ip= 50 %) en la población, mientras que Lynchia americana mostró como máximo valor el 37,5 % de Índice de prevalencia. De las dos especies de ectoparásitos hallados, Lynchia americana mostró una tendencia a encontrarse en pocos hospedadores con intensidades de 1 o 2 parásitos por hospedador, excepto en un solo ejemplar. Este resultado indicaría que estos ectoparásitos pudiesen clasificarse como estacionales. Estos resultados son coincidentes con los obtenidos por Dimare (2003) en su estudio sobre Zenaida asiatica en Tampisque, Costa Rica, donde el Índice de prevalencia del ácaro Falculifer spp. fue del 18 % y solo el 1 % de la mosca hematófaga Pseudolynchia canariensis.
En las fluctuaciones observadas en los Índices de prevalencias de los ectoparásitos entre los períodos climáticos del año, se observó una tendencia de Lynchia americana a variar de manera inversa al aumento del período menos lluvioso; esta diferencia coincide con las mayores abundancias de este ectoparásito observadas en un ejemplar, contrario a la conducta seguida por los ácaros podales que se mantuvieron presentes en la mayoría de los meses del año.
Para Medellín et al., (2019) estas conductas pueden ser el reflejo de los patrones de colonizaciones estacionales de los parásitos, de los cambios en la dieta de los hospedadores entre las dos épocas y/o de cambios en las condiciones climáticas. Las condiciones climáticas cambiantes y la distinta disposición de recursos en las diferentes épocas pueden producir efectos muy importantes sobre las poblaciones parásitas. La alta incidencia de un parásito en un período determinado le confiere una alta predictibilidad en esta época, y por tanto una labor preventiva consecuente con el período anual.
No se observaron grandes diferencias en los Índices de prevalencia entre la especie parásitas respecto a los sexos; este resultado pudiese estar relacionado con la no existencia de grandes desigualdades tróficas o de comportamiento entre machos y hembras y, por tanto, marcadas diferencias ecológicas entre sexos en la población de esta columbida que influyan en las cargas parasitarias. Estos resultados sugieren que el sexo del huésped no es un factor importante en la estructura de las comunidades parasitadas.
El análisis de los resultados hallados en las relaciones entre ambos ectoparásitos y la clase de edad refleja una mayor incidencia de ambos ectoparásitos hacia la población joven, lo que constituye una evidencia de la influencia de la clase edad de los hospedadores sobre la estructura de las comunidades parásitas en estas palomas.
Quizás estas diferencias observadas puedan explicarse por las características propias de cada clase de edad y sus respuestas ante los parásitos; además, existen otros factores como la habilidad de colonización, capacidad infectiva de las especies parásitas, factores ambientales y ecológicos que condicionan el acceso al hospedador. Una vez que el parásito llega al hospedero existen filtros inmunológicos típicos de cada clase de edad que determinan cuáles podrán instalarse y prosperar y cuáles no. Para Núñez et al. (2011), en general, una mayor masa corporal se interpreta como índice de mejor condición física porque indica, presumiblemente, la disponibilidad de reservas de nutrientes que permiten hacer frente a situaciones de elevada demanda. Es entonces indispensable determinar si existen diferencias en la población estudiada entre el peso corporal de los ejemplares parasitados y los no parasitados.
Una vez realizada la prueba de normalidad al peso de los ejemplares capturados separados por las categorías sanos y parasitados (incluye todos los ejemplares con cualquiera de los dos ectoparásitos hallados en este estudio), se procedió a realizar la comparación de las medias obtenidas en ambos grupos aplicando el test de T de Student para muestras independientes.
A pesar de que no se observaron diferencias estadísticamente significativas entre los grupos de aves (parasitadas y no parasitadas; T de Student (P= 0,189; P >0.05), se puede apreciar un valor menor en el peso de las aves parasitadas (Figura 10).
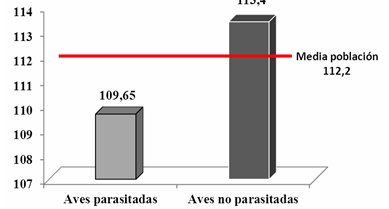
Fig. 10 - Comparación de la media del peso (g) de las aves parasitadas y no parasitadas en relación con la media poblacional de Zenaida macroura
A pesar de no ser fácil establecer las incidencias que pueden generar la parasitosis en un ave, los cambios en la disminución de la masa corporal encontrados en este estudio pueden sugerir deficiencias en el estado de salud general de las aves parasitadas. Por tal razón es aconsejable para determinar el estado de salud de la población de Z. macroura calcular índices relacionados con la masa corporal que puedan ser indicadores confiables del estado de salud poblacional.
Como se ha descrito antes, el estudio de la condición de salud de las poblaciones bajo algún tipo de manejo es necesario, pero este presenta limitaciones por su complejidad: técnicas que se aplican, difícil aplicación en el campo, costo económico de algunos análisis y la existencia de lagunas en el estado de conocimiento de los patógenos que pudiesen encontrarse. Siempre que sea posible la aplicación de diferentes técnicas y su combinación con índices darán resultados más confiables. Por lo tanto, aplicar la repetitividad en el tiempo de un índice de condición es clave en la interpretación de sus resultados. Además, la aplicabilidad de los índices puede variar entre diferentes especies, por lo que sería importante validar el método en cada caso, algo que en la realidad ocurre con muy poca frecuencia (Núñez et al., 2011).
En los ejemplares de Zenaida macroura estudiados se determinó la presencia de dos ectoparásitos Lynchia americana, que se hospedan en el plumaje de la especie y un ácaro desconocido que parasita las patas de la especie, con mayor incidencia de ambos en el período más lluvioso del país, en ejemplares jóvenes y sin diferencia de cargas parasitarias entre sexos. Lynchia americana presentó una alta temporalidad en los ejemplares parasitados, mientras que el ácaro podal estuvo presente en la mayoría de los meses del año.

