










 Servicios personalizados
Servicios personalizados texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
La deforestación, a nivel mundial es uno de los procesos de degradación ambiental de alto impacto negativo en la sostenibilidad y competitividad de los ecosistemas, que genera pérdidas de biodiversidad, degradación de suelos, aguas y cambios en los microclimas de las regiones afectadas y del clima a nivel global. A pesar de que en los últimos 25 años la deforestación ha disminuido, aún se siguen deforestando los bosques a nivel mundial (FAO 2018).
En la actualidad, la recuperación de áreas boscosas, la reducción de la tala y la adecuada gestión forestal se ha convertido en actividades prioritarias para restaurar los bosques, la biodiversidad que albergan y los servicios ambientales que brindan, como una estrategia para afrontar los efectos del cambio climático (Aitken et al., 2008; Aguirre-Mendoza et al., 2021c; Bautista-Miranda 2019).
El Ecuador es un país rico en recursos naturales, con una diversidad climática y biológica como bosques tropicales y andinos (Rosillo et al., 2020). Estos son considerados ecosistemas frágiles y vulnerables a los efectos combinados del cambio climático, deforestación y degradación (Aguirre-Mendoza et al., 2017). Al mismo tiempo, presentan potencial para contribuir a mitigar el cambio climático, restaurar funciones ecosistémicas, proveer bienes y servicios ambientales (Herzog et al., 2013).
Estudios en esta formación ecuatoriana demuestran alta diversidad florística de estos bosques, su gran heterogeneidad, diferencias en la composición de especies actual debido a la influencia antrópica y a la fragmentación (Aguirre-Mendoza et al., 2017, Aguirre-Mendoza et al., 2018).
Este estudio se realizó con la finalidad de evaluar la composición florística y estructural del bosque seco tropical del valle Sancán para facilitar elementos que permitan perfeccionar las acciones de manejo y conservación del remanente bosque seco.
MATERIALES Y MÉTODOS
Área de estudio y sitios de muestreo
El estudio se realizó en el sector Quimis, valle de Sancán perteneciente al cantón Jipijapa, ubicado al sur de la provincia de Manabí. Limita al Norte con la parroquia La Pila, al Sur con la comunidad Sancán, al Este con el recinto Cerrito la Asunción y al Oeste con la parroquia Membrillal (Figura 1). El clima es cálido seco con una temperatura entre 21 a 37 0C y precipitación media anual de 600 mm. Los suelos son arcillosos, sobre pendientes medias del 80 %. En esta región predomina el bosque deciduo de tierras bajas y el bosque semideciduo montano bajo y/o pie montano (Aguirre-Mendoza et al., 2021c).

Fig. 1. - Ubicación de las parcelas realizadas en el en el bosque seco tropical, sector Quimis, valle Sancán
En el área de estudio, se establecieron seis parcelas de muestreo con una superficie de 1 000 m2 (20 × 50 m), validadas mediante la curva de acumulación de especies. Para el establecimiento se utilizó brújula, GPS, estacas y piola. Se registraron todos los individuos de las especies arbóreas mayores a 5 cm de diámetro (D1,30). Se identificaron las especies y el número de árboles de cada individuo y se midieron la altura total (h) y diámetro (D1,30). El registro y colecta de los individuos leñosos siguió la metodología de Phillips et al. (2016).
Análisis de datos
Composición florística
Durante el levantamiento de la información en campo, se recolectó material botánico de todas las especies arbóreas existentes en los sitios de muestreo. Las muestras botánicas de todos los individuos fueron identificadas en el herbario ECUAMZ (Herbario Amazónico del Ecuador) del centro de investigación posgrado y conservación Amazónica (CIPCA) de la Universidad Estatal Amazónica. La nomenclatura de los nombres científicos sigue el sistema APG IV.
Índices de diversidad alfa y beta
Con el objetivo de medir la diversidad arbórea del boque se estimó el índice de diversidad de Shannon-Weaver (H'), que expresa la heterogeneidad de una comunidad; y el índice de diversidad de Simpson (1/D), que estima si una comunidad determinada está compuesta por especies con gran dominancia (Aguirre-Mendoza et al., 2021). Para evaluar la diversidad beta del bosque se realizó un análisis de conglomerado para detectar las similitudes entre parcelas. Estos análisis se realizaron con el programa BioDiversity Pro. ver. 2.0.
Cálculo de parámetros estructurales
Se calculó el índice valor importancia ecológica (IVIE) de cada especie el cual relaciona la abundancia relativa (Ar), frecuencia relativa (Fr) y dominancia relativa (Dr) (Manzanilla et al., 2020) (Tabla 1).

Estructura diamétrica
La estructura diamétrica del bosque se determinó considerando el número de individuos con D1,30 cm ≥ 5 cm y la pertenencia a las clases diamétricas establecidas por Aguirre-Mendoza et al. (2017) en función de determinar la distribución diamétrica de la vegetación.
RESULTADOS Y DISCUSIÓN
La curva de acumulación de especies indica que el número de parcelas muestreadas fue suficiente para representar la riqueza florística del bosque (Figura 2).

Composición florística
En el estudio, se registraron 21 especies, 20 géneros, 14 familias y un total de 236 individuos, resultados similares a los reportados por Jiménez et al. (2021) quienes reportaron 31 especies, 29 géneros y 16 familias en siete apiarios activos distribuidos dentro del recinto Quimís, y muy diferentes a lo reportado por Maldonado et al. (2018) en un bosque tropical de montaña que encontró 81 especies arbóreas.
Dentro de las especies identificadas, Pithecellobium arboreum es clasificada como Vulnerable (VU) y Croton rivinifolius en peligro de extinción (EN) según el Libro Rojo de la UICN (UICN, 2020), es decir que tiene un alto riesgo de extinción en la naturaleza. En este sentido, ambas especies enfrentan un riesgo de extinción o deterioro poblacional a mediano plazo.
Las especies Colicodendron scabridum y Bursera graveolens, según el Libro Rojo de UICN, están categorizadas como de menor preocupación (LC), es decir que no están en peligro crítico, al igual que Ceiba trichistandra, siendo esta última la más utilizada por las comunidades circundantes.
Las familias con mayor diversidad de especies fueron: Fabaceae con cinco especies, Capparaceae, Euphorbiaceae y Malvaceae con dos y el resto de familias representadas por una sola especie (Figura 3), resultados semejantes a lo reportado por Aguirre et al. (2021a) quienes registran a: Fabaceae, Asteraceae, Malvaceae y Rubiaceae como las familias de mayor diversidad de especies en el bosque seco del sector "El Tabanco", parroquia Mangahurco, Zapotillo, Ecuador. Igualmente, Jiménez et al. (2021) reportan a las familias Fabaceae, Ericaceae y Malvaceae como las más diversas. Según estos estudios las familias sobresalientes en los bosques secos son: Fabaceae y Malvaceae.

Parámetros estructurales
Las especies con mayor presencia en el bosque fueron Bursera graveolens, Geoffroea spinosa y Jacquinia sprucei, que también coinciden con las más abundantes. Ceiba trichistandra, Jacquinia sprucei y Bursera graveolens fueron las especies más frecuentes del bosque, mientras que Ceiba trichistandra, Bursera graveolens y Geoffroea spinosa son las especies dominantes.
Las especies con mayor valor de importancia fueron Ceiba trichistandra con 33,16 %, Bursera graveolens con 11,46 % y Geoffroea spinosa con 6,99 % (Tabla 2), lo que coincide con lo observado por Aguirre et al. (2021c) quienes reportan también como especie sobresaliente a Ceiba trichistandra. De la misma manera, Jiménez et al. (2019) indican que Ceiba trichistandra es la de mayor importancia y que es una especie típica y característica de los bosques secos del Ecuador y Perú, en comparación con otros bosques secos del mundo.
Por otro lado, se observó que entre las especies con menor IVIE se encuentran Machaerium millei con 0,85 % y Croton rivinifolius y Pithecellobium excelsum con 0,71 % cada una, resultados diferentes a los reportados por Aguirre et al. (2021b) que indican a Gynoxys nítida, Piper ecuadorense y Meliosma arenosa como especies con menor IVIE en el bosque Huashapamba, Loja, Ecuador; a Aguirre et al. (2021c) en Zapotepamba donde la especie ecológicamente menos importante fue Melinis minutiflora. Mientras que en parcela permanente del bosque seco del sector El Tabanco, Mangahurco, Ecuador las especies ecológicamente menos importantes son Machaerium millei y Terminalia valverdae(Aguirre et al., 2021a). La diferencia encontrada posiblemente se deba al grado de intervención antrópica y madurez de la vegetación.
Tabla 2. - Parámetros estructurales de las 10 especies principales del componente arbóreo del bosque seco tropical del sector Quimís, valle Sancán
| Especie | AA | AR (%) | FA | FR (%) | DA | DR (%) | IVIE (%) |
|---|---|---|---|---|---|---|---|
|
|
11 | 4,66 | 6 | 10,00 | 13,61 | 84,82 | 33,16 |
|
|
47 | 19,91 | 5 | 8,33 | 0,99 | 6,14 | 11,46 |
|
|
25 | 10,59 | 5 | 8,33 | 0,29 | 1,84 | 6,92 |
|
|
23 | 9,75 | 6 | 10 | 0,10 | 0,62 | 6,79 |
|
|
19 | 8,05 | 5 | 8,33 | 0,10 | 0,63 | 5,67 |
|
|
16 | 6,78 | 5 | 8,33 | 0,08 | 0,47 | 5,19 |
|
|
17 | 7,20 | 4 | 6,67 | 0,26 | 1,63 | 5,17 |
|
|
18 | 7,63 | 4 | 6,67 | 0,07 | 0,45 | 4,92 |
|
|
10 | 4,24 | 4 | 6,67 | 0,08 | 0,49 | 3,80 |
|
|
9 | 3,81 | 3 | 5,00 | 0,06 | 0,40 | 3,06 |
AA: Abundancia Absoluta, FA: Frecuencia Absoluta, DA: Dominancia Absoluta, AR: Abundancia Relativa, FR: Frecuencia Relativa, DR: Dominancia Relativa, IVIE: Índice de valor de importancia ecológica.
Índice de Shannon (H) y recíproco de Simpson (1/D)
De acuerdo con los resultados, la diversidad de especies según el índice de Shannon se clasifica como una diversidad alta en el área del sector Quimís, evidenciado por el resultado obtenido H=3,83. En este mismo sentido, se destacan las especies Bursera graveolens, en tanto que Croton rivinifolius y Pithecellobium excelsum presentan baja diversidad en la zona de estudio (Tabla 3). Los resultados del recíproco de Simpson Dominancia (1/D) = 0,06 y Simpson (D) =0,91 muestran una expectativa alta para realizar una selección de dos o más individuos al azar de una misma especie en la zona de investigación establecida.
Resultados similares fueron reportados por Aguirre-Mendoza et al. (2017), en el suroccidente del Ecuador que reportaron un valor de 2,51 lo que indica una diversidad media. De la misma manera, el índice de Simpson mostró una alta dominancia (S=0,93); similar a lo reportado por un estudio de la composición florística y estructura de los bosques secos de la provincia de Loja, Ecuador (Aguirre et al., 2013) quienes reportaron una dominancia alta (S= 0,89).
Tabla 3. - Índice de diversidad de Shannon de las especies de estudio del bosque seco tropical en el sector Quimís, valle de Sancá
| Especies (Nombre científico) | Ind por sp. | AR "PI" | Ln "PI" | PI*Ln (PI) | PI* Ln (PI)*-1 |
| 47 | 0,20 | -2,33 | -0,46 | 0,46 | |
| 25 | 0,11 | -3,24 | -0,34 | 0,34 | |
| 23 | 0,10 | -3,36 | -0,33 | 0,33 | |
| 19 | 0,08 | -3,63 | -0,29 | 0,29 | |
| 18 | 0,08 | -3,71 | -0,28 | 0,28 | |
| 17 | 0,07 | -3,80 | -0,27 | 0,27 | |
| 16 | 0,07 | -3,88 | -0,26 | 0,26 | |
| 11 | 0,05 | -4,42 | -0,21 | 0,21 | |
| 10 | 0,04 | -4,56 | -0,19 | 0,19 | |
| 9 | 0,04 | -4,71 | -0,18 | 0,18 | |
| 7 | 0,03 | -5,08 | -0,15 | 0,15 | |
| 6 | 0,03 | -5,30 | -0,13 | 0,13 | |
| 5 | 0,02 | -5,56 | -0,12 | 0,12 | |
| 5 | 0,02 | -5,56 | -0,12 | 0,12 | |
| 5 | 0,02 | -5,56 | -0,12 | 0,12 | |
| 4 | 0,02 | -5,88 | -0,10 | 0,10 | |
| 3 | 0,01 | -6,30 | -0,08 | 0,08 | |
| 2 | 0,01 | -6,88 | -0,06 | 0,06 | |
| 2 | 0,01 | -6,88 | -0,06 | 0,06 | |
| 1 | 0,00 | -7,88 | -0,03 | 0,03 | |
| 1 | 0,00 | -7,88 | -0,03 | 0,03 | |
| Total | 236 | 1 | -106,4 | -3,83 | 3,83 |
Ind por sp. = cantidad de individuos por especies en las seis parcelas; AR= abundancia relativa; PI= proporción del número de individuos de la especie i con respecto al total; Ln= logaritmo natural.
Diversidad beta
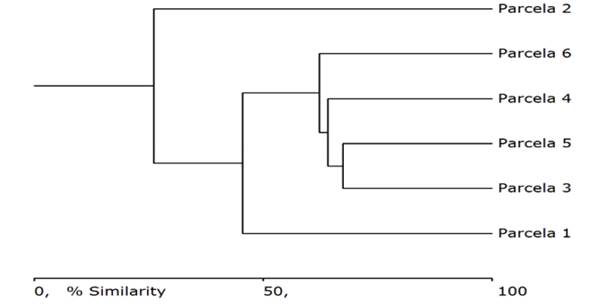
El dendrograma derivado de la matriz de similitud-disimilitud de Bray-Curtis (1957) (Figura 4) agrupa las parcelas de estudio en tres conjuntos bien definidos. En el grupo uno se encuentra la parcela 1, el dos las parcelas 3, 5, 4 y 6, y el tres conformado por la parcela 2. Las agrupaciones están influidas por el número de individuos de las especies con mayor composición florística, debido a que su abundancia resultó muy parecida, a pesar que de que su riqueza fue muy diferente entre los sitios de muestreo.
De manera general, la similitud de las parcelas del bosque seco tropical en el sector Quimís, se asemejan en más de 50 %, con clara heterogeneidad y diversas especies que se desarrollan en esta zona, ubicadas a 200 m de la carretera y 300 m s.n.m, influenciadas por actividades antrópicas como: tala de madera para leña, carpintería y uso de productos no maderables, lo que coincide con lo encontrado por Vistín-Guamantaqui y Espinoza (2021) en el bosque Siempreverde Montano Alto del Parque Nacional Sangay-Ecuador donde reportaron una similitud de especies de más de 50 %, y la tala selectiva, la extracción de leña y de otros productos forestales no madereros, como los factores que más alteran la dinámica del bosque, así como su estructura y composición.
Estructura diamétrica
En la Figura 5, se observa que las dos primeras clases diamétrica agrupan el 89,83 % del total de los individuos, demostrando así que el bosque está formado por árboles delgados en crecimiento. La clase diamétrica cinco (>40 cm) está constituida por seis individuos de Ceiba trichistandra, que ya han alcanzado su madurez, resultando una distribución diamétrica del bosque en forma de "J" invertida, situación que es corroborada por Aguirre et al. (2021c), en el bosque seco de Zapotepamba, Loja, Ecuador, el cual muestra una estructura diamétrica similar a la "J" invertida típica de bosques secos; de la misma manera Astudillo et al. (2019) y Aguirre et al. (2021b) confirma una distribución diamétrica en forma de "J" invertida que es característica de los bosques andinos, corresponde indicar que la comunidad estudiada es autoregenerativa, dado que existen alta concentración de individuos en las clases menores con reducción acentuada en las mayores.
La tendencia de la curva de la "J" invertida también señala que la comunidad vegetal se encuentra en desarrollo hacia etapas de crecimiento y productividad más avanzados, conforme lo afirma Lamprecht (1990), donde los abundantes individuos jóvenes van reemplazando a especímenes que se encuentran en la fase senil, confirmado por Aguirre-Mendoza et al. (2017) y Aguirre et al. (2018) en estudios en bosques andinos del sur de Ecuador.

CONCLUSIONES
La diversidad florística es alta, se ve expresada en la presencia de 21 especies, 20 géneros, 14 familias y 236 individuos. Las familias más diversas fueron: Fabaceae, Capparaceae, Euphorbiaceae y Malvaceae.
Las especies arbóreas ecológicamente importantes fueron: Ceiba trichistandra, Bursera graveolens y Geoffroea spinosa lo que sugiere que es un bosque seco que conserva especies endémicas y características de este tipo de ecosistema.
Las tres primeras clases diamétricas agrupan la mayor cantidad de árboles evaluados, que determina una distribución diamétrica con una tendencia a "J" invertida, indicando que el bosque seco del sector Quimís es un bosque en proceso de recuperación.
AGRADECIMIENTOS
Los autores agradecen al personal que integra el proyecto "Caracterización arbórea del bosque seco tropical y el impacto económico en la población del valle Sancán" de donde fueron generados los resultados. Igualmente agradece a la comuna Sancán y a su presidente que permitió trabajar en el territorio y apoyaron en el levantamiento de información de campo.

