










 Servicios personalizados
Servicios personalizados texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Los bosques de neblina montanos tropicales son ecosistemas de alta importancia en el mundo, abarcan alrededor de 380 000 km2 que representa un 2,5 % de los conocidos bosques tropicales (Rubb et al., 2004), esta importancia viene dada por su excepcional diversidad de flora que albergan al demostrar una alta riqueza de especies por unidad de área, siendo fuente importante de endemismos.
Estos se comportan como eficientes ecosistemas en cuanto a servicios ecosistémicos como la captura de carbono (Álvarez-Arteaga et al., 2013; Eller et al., 2020; Knowles et al., 2020) tanto en su biomasa arbórea y principalmente en el suelo, siendo superado sólo por ecosistemas altoandinos (páramos-puna) con alta concentración de carbono orgánico (Dilas-Jiménez y Huamán Jiménez, 2020; Zimmermann et al., 2010). Sin embargo, estos bosques se ubican en las zonas más sensibles con anomalías climáticas a diferencia de otros ecosistemas terrestres en el mundo (Seddon et al., 2016) y son amenazados con los efectos del cambio climático y principalmente son fragmentados y degradados con el cambio de uso de la tierra (Moreira et al., 2021).
En el Perú, los bosques de neblina montanos tropicales se ubican mayormente desde 1 000 hasta 3 000 m s.n.m (Van de Weg et al., 2014), en la zona norte peruana principalmente se ubican en las regiones de Cajamarca, Piura y amazonas, regiones con amplia diversidad florística al alrededor de 17 000 especies, 800 de estas endémicas (Sagástegui et al., 2003).
En la región Cajamarca, los bosques de neblina montanos tropicales se ubican principalmente en los territorios de las provincias de Jaén y San Ignacio (MINAM, 2014), los cuales por albergar especies maderables con alta demanda como Retrophyllum rospigliosii, Prumnopitys harmsiana (podocarpáceas) están sufriendo tala selectiva, además están amenazados por las altas tasas de deforestación en la zona alrededor 4 000 hectáreas año-1(Llerena et al., 2010). No obstante, hay escasa información sobre aspectos ecológicos e hidroecológicos que brinda la flora arbórea en estos tipos de bosque, en general, son muy escasos los estudios realizados y publicados para este tipo de bosques en el norte del Perú (Sagástegui et al., 2003; Seddon et al., 2016).
En la actualidad, Perú impulsa estudios sobre diversidad del componente arbóreo principalmente en el bosque amazónico, basados en parcelas de muestreo con dimensión mínima de una hectárea (de área contigua) y metodologías estandarizadas (Marcelo-Peña y Reynel, 2014). Sin embargo, para los bosques de neblina en la zona norte son muy pocos los registros de estudios publicados, entre ellos uno realizado por Peña y Pariente (2015) en un bosque de neblina en la provincia de San Ignacio, Cajamarca, a una altitud de 2 150 m s.n.m registró 308 individuos distribuidos en 31 familias, 30 géneros y 39 especies, siendo las cinco familias más con mayor índice de valor de importancia, en orden descendente, Podocarpaceae, Lauraceae, Rubiaceae, Melastomataceae y Clusiaceae. Marcelo-Peña y Arroyo (2013) en estudio realizado en bosque de neblina de la provincia de Jaén, Cajamarca, sobre los 2 100 msnm registró dos nuevas especies para la ciencia de la familia Magnoliaceae, Magnolia jaenensis cuyas primeras colectas se hicieron con este estudio y M. manguillo.
Ante ello, el presente estudio tuvo el objetivo de analizar la composición, diversidad y estructura de la vegetación arbórea en un área de bosque de neblina montano tropical sobre 2 100 m s.n.m en el norte del Perú.
MATERIALES Y MÉTODOS
Área de estudio
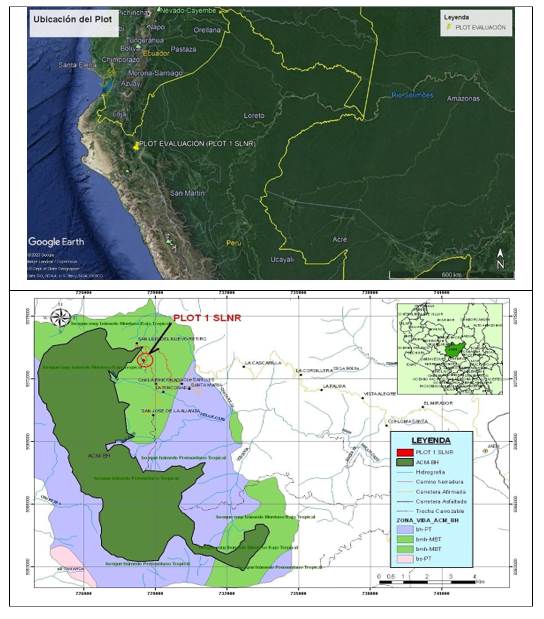
La investigación se desarrolló en una zona de relictos de bosque de neblina montano tropical, a una altitud promedio de 2 170 m s.n.m, localizado en la región Cajamarca, provincia de Jaén, distrito de Huabal, en el caserío San Luis del Nuevo Retiro, ubicado en las coordenadas UTM 0728564 este y 9372902 norte, zona 17M (Datum WGS 84). El sitio se ubica en el área de amortiguamiento del Área de Conservación Municipal Bosques de Huamantanga (ACM-BH).
El área de estudio, siguiendo el criterio de Zonas de Vida (Holdridge, 1978), pertenece a un bosque muy húmedo-Montano Bajo Tropical (bmh-MBT), con pendientes pronunciadas y de fisiografía ondulada, las temperaturas van desde los 12º C hasta los 17º C, y precipitaciones anuales entre 1900 mm y 3800 mm. Los suelos se caracterizan por ser ácidos y abundante materia orgánica (OIKOSlab, 2013 citado por Peña y Pariente, 2015) (Figura 1).
Instalación de la parcela de evaluación
Para la instalación de la parcela permanente se siguió la metodología usada por diversos autores RAINFOR, 2016; Synnott, 1979). La selección del sitio para la instalación fue un relicto de bosque de propiedad particular a fin de salvaguardar la conservación de la parcela para futuras remediciones, así como por su estado de conservación en cuanto a la flora existente. La localización de la parcela, el plaqueado y registro de árboles mayores o iguales a 10 cm de diámetro a la altura del pecho (DAP), así como de las primeras colecciones botánicas se realizó en el primer semestre del 2008 (Figura 2).
Colección botánica y registro de información
La colección botánica se desarrolló en dos momentos, la primera colección se ejecutó en el primer semestre 2008 y la segunda colección, al segundo semestre del mismo año, esto principalmente para las muestras que fueron colectadas como estériles en la primera colección, a fin de facilitar el trabajo de identificación; este trabajo se realizó utilizando el equipo estándar, así como los materiales y procedimientos recomendados para este tipo de trabajo (Rodríguez y Rojas, 2006). Para la herborización de las muestras colectadas se siguió metodología estandarizada en estos casos (Rodríguez y Rojas, 2006; Rotta et al., 2008). Los especímenes colectados fueron prensados, previa preservación con una solución de 60% de agua y 40 % de alcohol de 96º y trasladados desde campo hasta la ciudad de Jaén donde se hicieron los procedimientos de secado y montaje de muestras (Marcelo-Peña et al., 2011; Rotta et al., 2008). Los especímenes colectados, secados y montados fueron enviados al Herbario MOL de la Universidad Nacional Agraria La Molina para su identificación taxonómica.
Manejo y análisis de información
Para el manejo ordenado de datos, en campo se codificó con placas de aluminio a cada individuo censado dentro de la parcela permanente, para lo cual se utilizó códigos numéricos de 6 dígitos, siendo este mismo el código de colección, por ejemplo, código 108220, haciendo la lectura de izquierda a derecha, el primer número indica el número de Parcela (Parcela 1), los siguientes dos números indican el número de subparcela dentro de la parcela permanente (subparcela 08) y finalmente los tres últimos números indican el número del individuo (individuo 220).
Con el fin de conocer la efectividad del muestreo, en la medición de individuos realizados por subparcela se llevó el conteo del número de especies que incrementaron a medida que se incrementó el número de subparcelas analizadas, con lo cual se elaboró un gráfico de curva área-especie (Matteucci y Colma, 1982; Chu, Smith y Solow, 2014).
Con la información de la identificación taxonómica recibida del Herbario MOL, se procedió al análisis de las siguientes variables (Antón y Reynel, 2004; Matteucci y Colma, 1982):
Diversidad: se determinó el número de individuos por ha, número de familias, número de especies. Asimismo, se determinó los siguientes índices de diversidad (Ecuación 1); (Ecuación 2); (Ecuación 3) y (Ecuación 4):
Riqueza específica (S):
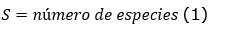
Índice de dominancia de Simpson (D):
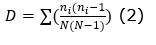
Índice de diversidad de Simpson (1-D, Simpson, 1949):
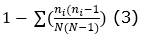
Índice de equidad de Shannon-Wiener (H, Hill, 1973):
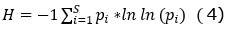
Composición: se determinó las familias y géneros más abundantes, especies endémicas y raras.
Variables dasométricas: se determinó el área basal (m2), altura total (m).
Estructura de la vegetación: se calculó la Densidad o Abundancia, la Frecuencia y la Dominancia de las especies identificadas. Con estos tres datos se obtuvo el Índice de Valor de Importancia (IVI) del total de especies, este índice permite medir cómo las especies contribuyen en la estructura de un ecosistema (Cottam y Curtis, 1956; Ragavan et al., 2015).
RESULTADOS Y DISCUSIÓN
Composición florística
Se registró 792 individuos para la parcela permanente de 1ha, pertenecientes a 81 especies de 48 géneros y 33 familias. El cociente de mezcla en este estudio es de 0,10 (81/792) es decir existen unos 10 individuos por cada especie, este resultado es similar a los encontrados en estudios en selva central del Perú (Marcelo-Peña y Reynel, 2014), pero inferior a otros estudios realizados en años anteriores en estas mismas zonas donde los cocientes de mezcla estaban alrededor de 0,22 (Antón y Reynel, 2004). El bajo coeficiente de mezcla encontrado estaría vinculado a la alta proporción de individuos de dos especies Cyathea sp. y Miconia punctata que representaron el 48 % del total.
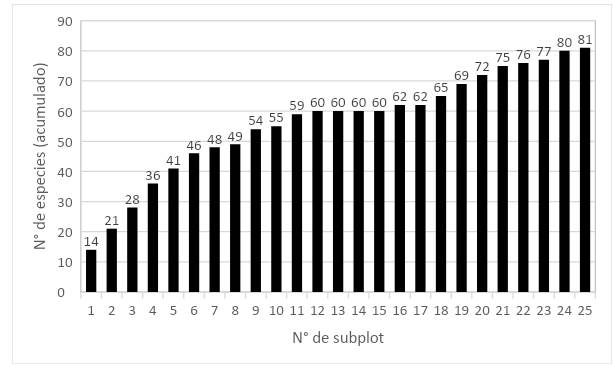
En al análisis de la curva área-especie, del total de especies registradas (81), se encontró que el 67 % de estas se alcanzó a completar en la subparcela 9 y el 85 % al completar la subparcela 19 (Figura 3).
Diversidad florística
La familia Lauraceae (25 especies) es la más rica en especies, seguida de Euphorbiaceae (cinco especies), Melastomataceae, Clusiaceae y Rubiaceae (con cuatro especies cada una). La familia Lauraceae es endémica en estos bosques montanos en el Perú, con mayor abundancia en sus géneros Ocotea y Nectandra(León, 2006), así también presencia importante de la familia Melastomataceae en su género Miconia(Ledo et al., 2012).
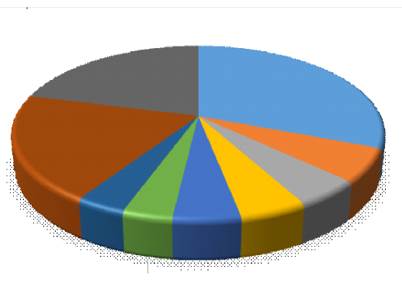
Fig. 4. - Abundancia relativa de especies por familia en el bosque montano de neblina, Jaén, Cajamarca
En evaluación realizada en los bosques de Chinchiquilla, San Ignacio, bosque montano de neblina a 2 150 m s.n.m, se encontró similares resultados en cuanto a las cinco familias con mayor número de individuos como Melastomataceae y Rubiaceae, y al cociente de mezcla encontrado en esta investigación, sin embargo, se evidencia una diferencia saltante debido a que en estos bosques de San Ignacio resalta en número las podocarpáceas o árboles conocidos en la zona como "romerillos" (Peña y Pariente, 2015).
En cuanto a la diversidad alfa según los índices analizados, donde una población sería homogénea si la distribución espacial entre especies es uniforme en la zona de estudio (Matteucci y Colma, 1982), los resultados para el presente estudio son: Riqueza específica (S), con un valor de 81 especies; Índice de dominancia de Simpson (D), con un valor de 0,138; Índice de diversidad de Simpson (1-D), con un valor de 0,862; y el Índice de equidad de Shannon-Wiener (H), con un valor de 2,856. Así, considerando al Índice de Shannon como un indicador de homogeneidad de un ecosistema evaluado (Shi y Zhu, 2009), el bosque evaluado en el presente estudio mostró valores intermedios, similares a los encontrados en un parcela permanente instalada en la misma zona sobre 2500 msnm, (Pérez, 2011), así también similares a resultados de evaluación en el Fundo Génova, Chanchamayo, en tipo de bosque secundario tardío (Antón y Reynel, 2004), pero inferiores a los valores de este índice (3,309) encontrados para el bosque un bosque montano de neblina en Chinchiquilla, San Ignacio, Cajamarca (Peña y Pariente, 2015) e inferiores a resultados de otros bosques premontanos y montanos en La Merced, San Ramón y Satipo en Perú (Marcelo-Peña y Reynel, 2014).
Endemismo
Al confrontar la lista de especies con el Catálogo de las Angiospermas y Gimnospermas del Perú(Brako y Zarucchi, 1993); se registró seis especies endémicas estas son:
Brunellia inermis Ruiz y Pav. (Brunelliaceae).
Alchornea brevistyla Pax y Hoffman (Euphorbiaceae).
Mezilaurus cf. opaca Kubitzki y Van der Werf (Lauraceae).
Persea raimondii O. Schmidt (Lauraceae).
Piper calvescentinerve Trelease (Piperaceae).
Roupala monosperma (Ruiz y Pav.) I.M. Johnst. (Proteaceae).
Cabe precisar que se encontró individuos de Podocarpus oleifolius (Podocarpaceae), que es la única familia de coníferas nativas del Perú y que es propia de los bosques nublados en las provincias de Jaén y San Ignacio en Cajamarca, en parcela permanente instalada en los bosques de Chinchiquilla en San Ignacio se encontró que estas podocarpáceas (Prumnopitys harmsiana y Podocarpus glomeratus) representan un 12% del total de individuos (Peña y Pariente, 2015). Estas podocarpáceas son protegidas en el Santuario Nacional Tabaconas Namballe (SNTN) pero son altamente amenazadas por la tala indiscriminada por ser una especie maderable muy requerida en la zona (Elliot, 2009; Llerena et al., 2010).
Asimismo, se destaca la presencia de un individuo que en este estudio se identificó preliminarmente como especie del género Talauma (Magnoliaceae), conocido como "militar. Esto fue base para estudios posteriores en la zona, principalmente en alrededores del poblado de San Luis del Nuevo Retiro, donde efectivamente se confirmó la existencia de especies de Magnoliaceas nuevas para la ciencia, reportándose así dos especies nuevas nombradas como Magnolia jaenensis y M. manguillo, estas nuevas especies a la vez corresponden a primeros registros de Magnolia en bosques montanos sobre los 2 100 m s.n.m (Marcelo-Peña y Arroyo, 2013).
No se descarta que pudiera haber otras especies endémicas o nuevas para la flora peruana e incluso para la ciencia; debido a que varios de los especímenes colectados, fueron registrados sin órganos reproductivos (flores y/o frutos), dificultando la identificación taxonómica de los especímenes hasta el nivel de especie.
Estructura de la vegetación (abundancia, frecuencia y dominancia)
De los 792 individuos censados, las cinco especies más abundantes y que representan un 63% del total de individuos son Cyathea sp. (30,93 %), Miconia punctata (17,17 %), Helicostylis tovarensis (6,69 %), Myrcia sp. (4,80 %) y Hedyosmum angustifolium (3,66 %), ver detalle en el apéndice 1.
Los cinco géneros más abundantes en orden descendente son Cyathea (Pteridophyta) con 245 individuos (30,93 %), Miconia (Melastomataceae) con 140 individuos (17,68 %), Helicostylis (Moraceae) con 53 individuos (6,69 %), Myrcia (Myrtaceae) con 42 individuos (5,30 %) y Ocotea (Lauraceae) con 32 individuos (4,04 %).
Así también las cinco más abundantes en orden son Cyatheaceae (31,31 %), Melastomataceae (18,31 %), Lauraceae (7,58 %), Moraceae (7,20 %) y Myrtaceae (5,30 %). Cyatheaceae fue la familia más abundante en el sitio estudiado. Estas familias son propias de estos tipos de bosques montanos en América (Ledo et al., 2012; Schin-ichiro y Kitayama, 1999), incluso algunas de estas también presentes en otros continentes (Shi y Zhu, 2009).
En cuanto a la frecuencia y dominancia, estas variables guardan estrecha relación con la abundancia de las especies. En la Tabla 1, se muestra el Índice de Valor de Importancia (IVI) para las diez especies más importantes (ver detalle en apéndice 1), donde se puede ver que sólo dos especies Cyathea sp. y Miconia punctata representan cerca de la tercera parte del IVI total.
Tabla 1. - Índice de Valor de Importancia de las diez especies más importantes del bosque estudiado
| FAMILIA | ESPECIE | DENSIDAD RELATIVA | FRECUENCIA RELATIVA | DOMINANCIA RELATIVA | IVI |
|---|---|---|---|---|---|
| CYATHEACEAE |
|
30,93 | 7,96 | 19,86 | 58,76 |
| MELASTOMATACEAE |
|
17,17 | 7,32 | 9,40 | 33,90 |
| MORACEAE |
|
6,69 | 6,05 | 9,22 | 21,96 |
| ANNONACEAE |
|
3,03 | 4,78 | 5,48 | 13,29 |
| MYRTACEAE |
|
4,80 | 5,10 | 3,38 | 13,27 |
| EUPHORBIACEAE |
|
1,89 | 3,50 | 4,75 | 10,15 |
| CECROPIACEAE |
|
3,16 | 3,18 | 2,96 | 9,30 |
| CHLORANTHACEAE |
|
3,66 | 4,14 | 1,49 | 9,29 |
| CLUSIACEAE |
|
1,89 | 3,82 | 3,03 | 8,74 |
| LAURACEAE |
|
1,64 | 2,55 | 3,82 | 8,01 |
La notable abundancia de la Cyatheaceae en el bosque estudiado es debida a que esta familia es representativa de los bosques montanos, se reconocen al menos unas 83 especies del género Cyathea en el Perú (Lehnert, 2011).
Estos resultados también muestran similitud con resultados de otros estudios en ecosistemas similares en Selva Central del Perú, donde también se encontró la dominancia del género Miconia de las melastomatáceas (Lehnert, 2011), aunque en este estudio si bien es una de las familias más abundantes, es mucho menos diversa que las lauráceas (Figura 4).
De acuerdo al análisis de Densidad, Frecuencia y Dominancia de todas especies encontradas en la parcela permanente, el Índice de Valor de Importancia (IVI) encontrado para las cinco primeras especies, en orden decreciente es: Cyathea sp 1 (58,76), Miconia punctata (33,90), Helicostylis tovarensis (21,96), Guatteria dielsiana (13,29) y Myrcia sp 1 (13,27).
Estos resultados difieren en cuanto a su nivel de importancia para las cinco primeras especies encontradas en los bosques montanos de neblina en la zona de San Ignacio, Cajamarca, donde las especies Prumnopitys harmsiana y Podocarpus glomeratus (podocarpáceas) son las más importantes, seguidas de Cinchona sp., Cecropia sp. y Endlicheria sp. (Peña y Pariente, 2015). Especies del género Miconia (melastomataceae), especies del género Guatteria (annonaceae) y especies de Moraceas se reportan entre las primeras 10 especies de importancia en bosques montanos y premontanos en el Perú (Antón y Reynel, 2004; Marcelo-Peña y Reynel, 2014; Peña y Pariente, 2015). En el Apéndice 1, se muestra el Índice de Valor de Importancia (IVI) del total de las especies.
Análisis las variables dasométricas y volumétricas las especies
De todos los individuos censados dentro, el diámetro (DAP) promedio fue 19,64 cm, con una varianza de 90,5; el área basal total es 29,62 m2, el área basal promedio por individuo es 0,037 m2, con una varianza de 0,002; la altura total de los árboles tiene un promedio de 10,53m, con una varianza de 13,86; y la altura comercial tiene un promedio de 6,72 m, con una varianza de 5,19. En evaluación realizada en la misma zona boscosa pero a una altitud de 2543 msnm se encontró un DAP promedio de 16,44 cm (Pérez, 2011) lo cual indica que a mayor altitud los diámetros de los árboles reducen, mientras que en otro bosque de neblina en San Ignacio, Cajamarca cuya Parcela permanente se instaló a una altura similar al presente estudio se encontró un DAP promedio de 25.20 cm, este mayor diámetro fue influenciado por la alta presencia de individuos de la familia Podocarpaceae que presentaron las mayores altura y diámetros, pues corresponden a especies forestales maderables de la zona (Peña y Pariente, 2015).
CONCLUSIONES
Los bosques de neblina montanos tropicales ubicados en la zona alta de la provincia de Jaén tienen importante diversidad de especies, así como especies endémicas y potencialidad de albergar especies nuevas para la ciencia.
La familia Lauraceae con 15 especies registradas fue la familia más diversa en el bosque estudiado, confirmándose así que las lauráceas son una de las familias más diversas en los bosques montanos en el Perú.
En cuanto a la estructura vegetal del bosque estudiado, la alta abundancia de la familia Cyatheaceae y, así como de la1familia Melastomataceae confirma la representatividad de especies de estas familias en los bosques montanos de neblina.

