










Meu SciELO
 Serviços customizados
Serviços customizadosServiços Personalizados
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista Cubana de Medicina
versão On-line ISSN 1561-302X
Rev cubana med v.47 n.4 Ciudad de la Habana oct.-dez. 2008
TEMAS ACTUALIZADOS
La resistencia a antibióticos en Helicobacter pylori
The resistance to antibiotics in Helicobacter pylori
Mayrín Hernández Power; Orlando Reyes Zamora; Boris L. Rodríguez González
Licenciado en Microbiología. La Habana, Cuba.
RESUMEN
Se hizo una revisión sobre el tema de la resistencia a antibióticos en Helicobacter pylori dirigida a especialistas de la salud, particularmente a los gastroenterólogos. Se sabe que la actualización constante resulta una importante herramienta para combatir la infección por H. pylori y evitar el aumento de la resistencia del microorganismo a los antibióticos de elección en nuestro país. Se abordaron los mecanismos moleculares de acción de los antibióticos más empleados en las terapias erradicadoras (metronidazol, amoxicilina, tetraciclina y claritromicina); así como los genes del microorganismo involucrado en la resistencia a cada antibiótico. Se aportaron datos sobre otros antibióticos menos usados, como la levofloxacina y ciprofloxacina. Se incluyeron, además, los métodos más usados para detectar la resistencia de la bacteria a cada antibiótico.
Palabras clave: Helicobacter pylori, resistencia, antibióticos, métodos de detección.
ABSTRACT
A review on the topic of resistance to antibiotics in Helicobacter pylori directed to health specialists and, particularly, to gastroenterologists was made. It is known that the constant updating is an important tool to fight H. pylori infection and to prevent the increase of the resistance of the microorganism to election antibiotics in our country. The mollecular mechanisms of action of the antibiotics most commonly used in erradication therapies (metronidazole, amoxicillin, tetracycline and claritromicine), as well as the genes of the microorganism involved in the resistance to every antibiotic, were approached. Data on other less used antibiotics, such as levofloxacin and ciprofloxacin were given. The most used methods to detect the resistance of the bacterium to each antibiotic were also included.
Key words: Helicobacter pylori, resistance, antibiotics, detection methods.
INTRODUCCIÓN
Helicobacter pylori es un microorganismo que posee una morfología bacilar y espirilada, es microaerofílico y coloniza eficientemente el estómago humano. Esta bacteria fue aislada por primera vez en 1983 por Marshall y Warren y este descubrimiento revolucionó los estudios de las enfermedades gastrointestinales, los cuales a partir de ese momento tuvieron una óptica diferente.1 La infección por H. pylori ha ido en aumento en las diferentes áreas geográficas, su diseminación resulta mucho mayor en países en vías de desarrollo aunque los estudios se concentran fundamentalmente en países desarrollados que poseen un gran potencial científico. Se estima que más del 50 % de la población mundial está infectada con este patógeno, particularmente en países en vías de desarrollo la tasa de infección sobrepasa el 80 % de la población.2 La persistencia de la infección con H. pylori puede ocasionar la aparición de diversas afecciones como gastritis, úlcera gástrica o duodenal, carcinoma del tejido linfoide asociado a mucosa (MALT) y cáncer gástrico, por lo anterior es considerado el principal agente etiológico para la mayoría de los desórdenes gástricos y fue declarado en 1994, por la Organización Mundial de la Salud, como agente carcinogénico de tipo I para el hombre.3
En estudios realizados in vitro, H. pylori mostró ser susceptible a una amplia gama de antibióticos, pero la mayor parte de ellos pierden su actividad in vivo por las condiciones de pH bajo existentes en el estómago. Esta limitante ha traído como consecuencia el uso de inhibidores de la bomba de protones (omeprazol, lanzoprazol) en la terapia, los cuales facilitan la acción de los antibióticos. Los antibióticos más frecuentemente usados en el tratamiento de la infección con H. pylori son el metronidazol, la amoxicilina, la tetraciclina y la claritromicina. No obstante, se han utilizado otros en menor medida, tal es el caso de la ciprofloxacina, levofloxacina, moxifloxacina, azitromicina, eritromicina, furazolidona y nitrofurantoína.4,5 Actualmente, la terapia que se considera correcta por los grupos más prestigiosos que estudian a H.pylori es la triple terapia (2 antibióticos y 1 inhibidor de la bomba de protones) y en caso de fallo, la cuádruple terapia que incluye, además de la triple medicamentación, sales de bismuto.6 El principal objetivo de esta revisión es actualizar el conocimiento de los mecanismos y formas de detección de la resistencia a los antibióticos empleados en la terapia contra H. pylori.
RESISTENCIA DE H. PYLORI A LOS ANTIBIÓTICOS EMPLEADOS EN LA TERAPIA
Metronidazol
En el tratamiento contra H. pylori, el metronidazol es uno de los antibióticos más utilizados a escala mundial, su empleo en otras afecciones de forma indiscriminada ha traído como resultado el desarrollo de altos niveles de resistencia.7 En la terapia se emplean dosis de 500 mg, 2 veces al día de 1 a 2 sem.8
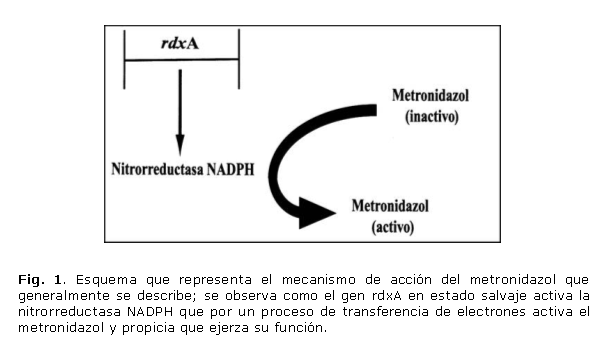
El metronidazol es un antibiótico bactericida que pertenece al grupo de los nitroimidazoles, es un compuesto heterocíclico con un núcleo de 5 átomos de carbono y 1 radical nitro (NO2), que es su grupo funcional. Cuando entran a la célula diana los nitroimidazoles son activados por un proceso de transferencia de electrones, mediado por la enzima NADPH nitrorreductasa, transformándolos en intermediarios imidazoles que causan daños letales a las estructuras celulares y fragmentando al ADN.9,10
Existen 2 genes que, se ha demostrado, están involucrados en la resistencia de H. pylori a este antibiótico, ellos son rdxA y frxA. El gen rdxA, de aproximadamente 630 pb, codifica para una NADPH nitrorreductasa insensible al oxígeno que dona los electrones reduciendo al metronidazol y activándolo.11 Uno de los mecanismos más importantes en la resistencia de H. pylori a metronidazol son las mutaciones que inactivan al gen rdxA. Estas mutaciones pueden presentarse por inserciones de pares de bases, sustituciones que originen codones de parada o simplemente sustituciones que en su conjunto hacen a la proteína disfuncional. No se ha observado que exista alguna zona del gen más importante o con mayor predisposición a sufrir mutaciones. Entre las mutaciones descritas pudieran citarse los cambios de aminoácidos cisteína por tirosina (C19Y) y treonina por lisina (T49K), que por si solos hacen que la enzima sea inactiva y la inserción de una adenina en la posición 141 originándose un codón de parada entre otras muchas reportadas en la literatura.12,13
El gen frxA, con una talla similar a rdxA (654pb), codifica para la enzima NADPH flavinoxidorreductasa que tiene un mecanismo similar a rdxA y constituye un importante factor en la activación del metronidazol en el interior celular. La resistencia debida a la disfuncionalidad de este gen se ha observado solo en algunas cepas. En algunos casos frxA promueve por sí solo la aparición de resistencia y en otros se presenta mutado junto a rdxA provocando ambos la resistencia a este antibiótico. El gen de frxA no presenta zonas definidas de mayor probabilidad de mutaciones, o sea, que no se conocen mutaciones que tributen directamente a un fenotipo resistente a metronidazol.14 No obstante, se ha descrito con cierta regularidad la deleción de adenina (A) en la posición 54 y se han observado también importantes cambios como el de arginina por histidina en la posición 16, y las sustituciones de bases adenina por guanina A217G y citosina por timina C215T (fig. 1).15,16
Amoxicilina
Un antibiótico fundamental en la terapia contra H. pylori es la amoxicilina, la cual ha mostrado niveles de resistencia muy bajos en el tratamiento de H. pylori.17-19 El modo de utilización consiste en 1g, 2 veces al día por 1 o 2 sem.8
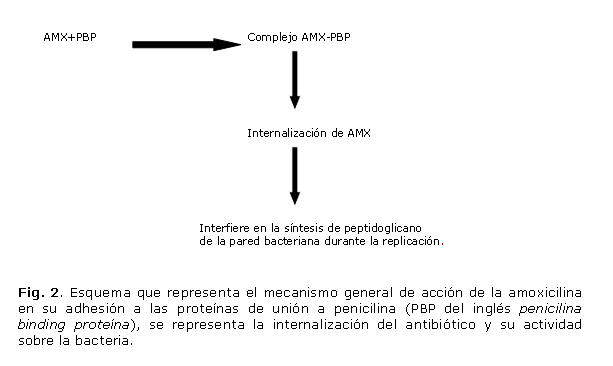
Tiene acción bactericida y actúa frente a microorganismos grampositivos y gramnegativos, inhibiendo la biosíntesis y reparación de la pared bacteriana. Es una penicilina semisintética susceptible a la acción de las betalactamasas. La base principal de su elevada especificidad y potencia bacteriana es una estructura cíclica, el anillo ß-lactámico, el cual debe permanecer intacto para que la penicilina ejerza su acción. La inadecuada formación de la pared bacteriana, en las especies susceptibles, produce un desequilibrio osmótico que afecta especialmente a las bacterias en fase de crecimiento. La amoxicilina presenta algunas ventajas destacables frente a otras aminopenicilinas como son: mejor absorción a partir del tracto alimentario, mejor capacidad para alcanzar concentraciones efectivas en los lugares de acción y mayor capacidad para penetrar en la pared celular bacteriana, atributos que la hacen importante en el tratamiento de H. pylori. Para que este antibiótico cumpla su función es necesaria la presencia de las proteínas de unión a penicilina (PBPs, del inglés penicillin binding proteins). Éstas se encuentran localizadas en la superficie de la membrana bacteriana y es mediante ellas que se internaliza el antibiótico. Una vez que la amoxicilina entra, bloquea la síntesis de la pared por interferencia del anillo ß-lactámico lo que propicia la lisis celular por desequilibrio osmótico.20
El mecanismo de resistencia que H. pylori ha creado contra la amoxicilina consiste en mutaciones en las PBPs. Particularmente, mutaciones en la PBP1A, que presenta mayor afinidad por la amoxicilina que las restantes proteínas de la familia por lo que al mutar se afecta la efectividad del antibiótico. Las mutaciones descritas en este gen se localizan fundamentalmente en el motivo II, cuya secuencia aminoacídica es serina-lisina-asparagina (SKN402-404) y en el motivo III, compuesto por los residuos lisina-treonina-glicina (KTG555-557) o en regiones adyacentes (región C-terminal de la proteína). Esto provoca cambios conformacionales en las PBPs que disminuyen la afinidad de unión a la amoxicilina y por tanto podrían dar lugar a fenotipos resistentes.21 Existen mutaciones que se han reportado en la literatura como las más frecuentes en las cepas resistentes, la mutación universal es la sustitución de la serina de la posición 414 por arginina (S414R) la cual se ha demostrado que por sí sola tributa a un fenotipo resistente. También existen otras mutaciones con alta frecuencia de aparición como las sustituciones de: serina por glicina (S402G), glutamato por alanina (E406A), serina por treonina (S417T), treonina por serina (T555S), asparagina por tirosina (N561Y), serina por arginina (S542R), treonina por isoleucina (T540I) e isoleucina por valina (I562V) que en su conjunto favorecen la aparición de resistencia.20,22 El mecanismo de resistencia de H. pylori a la amoxicilina ha sido bien descrito y demostrado. En la figura 2 se muestra cómo en las cepas resistentes, al imposibilitarse la formación del complejo AMX-PBP1A, se evita la acción bactericida del antibiótico y puede crecer en presencia de este.
Tetraciclina
La tetraciclina es otro de los antibióticos bacteriostáticos que se usa frecuentemente en la terapia erradicadora, sobre todo en el régimen de segunda línea. Su estructura química lo ubica en el grupo de las tetraciclinas y es sintetizada de forma natural por 3 microorganismos: S. aureofaciens, S. rimosus y S. viridofaciens.23
En la terapia erradicadora, la dosis de tetraciclina prescrita por los médicos es de 500 mg cada 8 h alrededor de 10 días.8
El mecanismo de acción de la tetraciclina consiste en bloquear la síntesis de proteínas en las bacterias. Esto lo realiza uniéndose a la subunidad ribosomal 30S del ribosoma e impidiendo la unión del ARN de transferencia aminoacil (ARNt-aa) a su sitio de unión.24,25 De esta forma, se bloquea el paso de elongación y, en consecuencia, se detiene la síntesis de proteínas. Esto provoca que los microorganismos no puedan multiplicarse, lo cual convierte a este fármaco en un potente bacteriostático.
Desde el año 1950, este antibiótico ha sido ampliamente empleado en la práctica médica y, por este uso extensivo, muchos patógenos bacterianos han desarrollado resistencia a este fármaco.23
En H. Pylori, el mecanismo de resistencia a tetraciclina demostrado es la presencia de mutaciones en el gen rrn 16S que codifica para el ácido ribonucleico ribosomal 16S de la subunidad 30S del ribosoma bacteriano. Las mutaciones se producen fundamentalmente en el triplete de bases adenina, guanina y adenina de las posiciones 926 a la 928 (AGA926-928) del gen rrn 16S, justo en la llamada caja C (fig. 3) y pueden producirse como simple, dobles o hasta triples sustituciones.26
Es un hecho ya demostrado que la concentración del antibiótico que una cepa resistente es capaz de tolerar, depende directamente del número de mutaciones que presenta en el triplete de bases AGA926-928. De esta forma, la triple sustitución de las bases AGA926-928 por timina, timina y citosina, respectivamente (AGA926-928®TTC), permite que las cepas toleren concentraciones del antibiótico de hasta 64 mg/L, las dobles mutaciones permiten tolerar concentraciones de hasta 8 mg/L y la simple sustitución de una de estas bases permite tolerar concentraciones mayores o iguales de 2 mg/L.27,28
En los últimos años se ha estado debatiendo la existencia de otros posibles mecanismos de resistencia a tetraciclina en H. pylori. Son los casos de la acción de la proteína de eflujo TetA (P) y de un mecanismo de reducción de la permeabilidad de la membrana.
En el año 2006, Li y Dannelly evidenciaron, in vitro, el aporte de esta proteína de eflujo TetA (P) en un evento transitorio de resistencia a tetraciclina que depende de la presencia sostenida del antibiótico en H. pylori (resistencia inducible). Estos investigadores realizaron experimentos donde mostraron que las mutaciones en el gen que codifica para esta proteína favorecieron la pérdida de la resistencia inducible a tetraciclina.29 Entre las funciones de esta proteína pudiera estar la expulsión del antibiótico fuera de la célula y la disminución de su concentración intracelular minimizando su efecto.
Por otra parte, también se ha expuesto la hipótesis sobre la existencia de una reducción de la permeabilidad de la membrana a tetraciclina. Este mecanismo hipotético permitiría explicar la aparición de aislados de H. pylori resistentes, que no tienen mutaciones en la secuencia AGA926-928 de su rrn 16S, pero muestran menor acumulación intracelular de este fármaco.30
Sin embargo, sólo ha sido demostrada la presencia del gen que codifica para la supuesta proteína de eflujo TetA (P) en todas las cepas de H. pylori, pudiendo o no su acción constituir otro posible mecanismo de resistencia a tetraciclina.
Se estima que la prevalencia de la resistencia a tetraciclina es menor del 1%, excepto en algunos pocos países asiáticos como Corea del Sur17, China31 y en Japón.32
Claritromicina
La claritromicina es el antibiótico menos usado en la terapia anti-H. pylori en Cuba.
Pertenece al grupo de los macrólidos es un derivado semisintético de la eritromicina y constituye la droga de más potencia contra H. pylori. Este fármaco se absorbe mejor en la mucosa gástrica, es más estable en pH ácido y, por tanto, más efectiva contra H. pylori. 33
La claritromicina es prescrita por los médicos en una dosis de 500 mg cada 12 h por 10 d.8
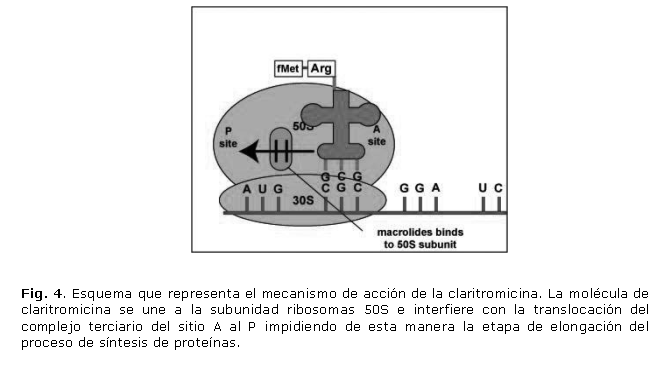
Al igual que la tetraciclina, es un antibiótico bacteriostático que afecta la síntesis de proteínas. La unión del macrólido a la subunidad ribosomal 50S del ribosoma bacteriano impide la translocación del complejo terciario: ARNt-aa, factor de elongación Ef-Tu, molécula de GTP, del sitio A al P del ribosoma, interfiriendo con la etapa de elongación en la síntesis de proteínas (fig. 4). La unión se realiza mediante la formación de puentes de hidrógeno entre diferentes radicales hidroxilo del macrólido y determinadas bases del ARNr 23S.34
La resistencia a claritromicina en H. pylori es el resultado de mutaciones puntuales que se presentan en el gen rrn 23S que codifica para el ARNr 23S de la subunidad ribosomal 50S. Las mutaciones más frecuentes son la sustitución de la adenina por una citosina o guanina en las posición 2142 (A2142C, A2142G) o por una guanina en la posición 2143 (A2143G). 35,36 Como consecuencia se produce una modificación en la estructura del ribosoma que afecta la unión permanente del antibiótico y, por tanto, no se afecta la síntesis de proteínas.36
Estas mutaciones provocan que las cepas resistentes toleren concentraciones crecientes del antibiótico que van desde 1mg/L hasta 64 mg/L. 36,37
La prevalencia de la resistencia a claritromicina varía según su consumo en las diferentes regiones. En Europa y en Estados Unidos de Norteamérica se pueden encontrar valores entre 9 y 18 %7,38 y en Canadá se estiman valores de prevalencia menores del 4 %.7
Los valores más preocupantes de prevalencia se encuentran en América del Sur, específicamente en Perú con 50 %39 y en el sureste asiático con 29 %,40 valores que son el resultado de un mayor consumo de este antibiótico en esos países.
Otros antibióticos
La levofloxacina y la ciprofloxacina son antibióticos alternativos que se usan en la terapia contra H. pylori y que han mostrado buena efectividad en el tratamiento. Específicamente, se prescriben en dosis de 250 mg 2 veces al día.41,42 a pacientes alérgicos a amoxicilina o a aquellos que no han erradicado el microorganismo después de la triple terapia.
Ambos antibióticos son bactericidas y pertenecen al grupo de las quinolonas. Su mecanismo de acción es similar y consiste en bloquear la replicación del ADN bacteriano por unión con la enzima ADN girasa.43 La acción fundamental de estas quinolonas ocurre sobre la subunidad A de la ADN girasa (gyr A), en la región determinante de resistencia a quinolonas (QRDR, del inglés quinolone resistance-determining region).44 La resistencia de H. pylori a ciprofloxacina y levofloxacina está dada por mutaciones que aparecen en QRDR. Las que más predominan son aquellas que provocan sustituciones de aspartato por glicina (D91G) y asparagina por lisina (N87K).45
Los niveles de resistencia son muy bajos, en el caso de la ciprofloxacina, que es el más empleado, se han obtenido niveles de resistencia de 5,7 %, fundamentalmente en Europa.45,46 La levofloxacina también ha mostrado valores bajos de resistencia en las terapias en las que ha estado incluido.41
MÉTODOS DE DETECCIÓN DE LA RESISTENCIA A METRONIDAZOL, AMOXICILINA, TETRACICLINA Y CLARITROMICINA
Los métodos de detección de la resistencia se clasifican en fenotípicos o técnicas de cultivo y genotípicos o técnicas moleculares.8 Los métodos fenotípicos más empleados para detectar la presencia de cepas de H. pylori resistentes a estos antibióticos son la dilución en agar y el E-test, que han mostrado ser efectivos en estudios de importantes grupos investigativos en esta temática.33
Los estudios de comparación entre estos métodos respecto a los métodos moleculares han mostrado que sus resultados son definitivo pues se basan en la observación directa del microorganismo y que este sea viable, lo cual asegura la positividad. Algunas desventajas, es que son menos consistentes, más lentos, necesitan suplementos especiales que son costosos, menos sensibles y fracasan en un 10 % de los casos por contaminación o porque la bacteria no crece.8 De ahí, que los métodos moleculares constituyen una interesante y ventajosa alternativa, ya que son rápidos y no requieren la viabilidad de las bacterias, se obtienen resultados reproducibles y fáciles de estandarizar. Estos métodos presentan la dificultad de dar falsos negativos por reactividad cruzada o por la presencia de inhibidores enzimáticos, además de poseer un equipamiento muy costoso.8
Un método novedoso que se está utilizando en el estudio de la resistencia de H. pylori a metronidazol es la detección de la proteína RdxA por Western Blot. En esta técnica se emplean anticuerpos de conejo que reconocen específicamente a la proteína RdxA mutada.47
Una técnica molecular que resulta útil para la detección de las mutaciones es la secuenciación posterior a la amplificación por reacción en cadena de la polimerasa (PCR) de un fragmento del gen donde están las mutaciones que contribuyen a la resistencia. Esta metodología resulta eficiente para cualquiera de estos 4 antibióticos.7
Para la amoxicilina, la tetraciclina y la claritromicina se sigue empleando un método que no por costoso ha dejado de realizarse: el PCR en tiempo real.7 Ésta es una técnica molecular donde al mismo tiempo que se amplifica por PCR el fragmento del gen asociado a la resistencia, se van detectando las mutaciones que puedan estar presentes mediante el uso de una sonda marcada que es complementaria al alelo susceptible. 37,48
Otra técnica molecular que también se ha usado mucho para detectar resistencia a tetraciclina y claritromicina es el PCR-RFLP. Este es un método relativamente simple que se basa en el hecho de que las mutaciones implicadas en la resistencia introducen nuevos sitios de restricción dentro del fragmento amplificado por PCR donde están presentes. Luego, con el uso de enzimas de restricción que cortan en esos sitios, los fragmentos amplificados son escindidos y dan, como resultado, fragmentos de diferente talla molecular. Los fragmentos amplificados de las cepas sensibles no son escindidos por las enzimas por lo que van a poder ser diferenciados de las cepas resistentes.8
Por todo lo planteado en este artículo, se infiere que resulta de vital importancia el estudio de la resistencia a antibióticos de cepas de H. pylori circulantes en una región determinada. Particularmente en Cuba nunca se ha desarrollado una investigación de este tipo, la cual resultaría de gran relevancia por el impacto que pudiera tener en lograr terapias erradicadoras más efectivas.
REFERENCIAS BIBLIOGRÁFICAS
1. Marshall BJ, Warren JR. Unidentified curved bacilli in the stomach of patient with gastritis and peptic ulceration. Lancet. 1984;1:1311-5.
2. Ahuja V, Sharma MP. High recurrence rate of Helicobacter pylori infection in developing countries. Gastroenterology. 2002;123:653-4.
3. World Health Organization. IARC monographs on the evaluation of carcinogenic risk to human. 1994;61:177-240.
4. Gisbert JP, Pajares JM. Helicobacter pylori "rescue" therapy after failure of two eradication treatments. Helicobacter. 2005;10:363-72.
5. Campo SMA, Zullo A, Hasan C, Moroni S. Antibiotic treatment strategies for Helicobacter pylori. Rec Pat Anti-Infective Drug Disc. 2007;2:1-72.
6. Vakil N. Eradication Therapy for Helicobacter pylori. Gastroenterology. 2007;133:1001.
7. Megraud F, Lehours P. Helicobacter pylori Detection and Antimicrobial Susceptibility Testing. Clin Microbiol Rev. 2007;20:280-322.
8. Guerrits MM, van Vliet A, Kuipers EJ, Kusters JG. Helicobacter pylori and antimicrobial resistance: molecular mechanisms and clinical implications. Lancet Infect Dis. 2006;6:1-10.
9. Sisson G, Jeong JY, Goodwin A, Bryden L, Rossler N, Lim-Morrison S et al. Metronidazole activation is mutagenic and causes DNA fragmentation in Helicobacter pylori and in Escherichia coli containing a cloned H. pylori RdxA(+) (Nitroreductase) gene. J Bacteriol. 2000;182:5091-6.
10. van der Wouden EJ, Thijs JC, Kusters JG, van Zwet AA, Kleibeuker JH. Mechanism and clinical significance of metronidazole resistance in Helicobacter pylori. Scand J Gastroenterol. 2001;36 Suppl 234:10-14.
11. Kaakoush NO, Mendz GL. Helicobacter pylori disulphide reductases: role in metronidazole reduction. FEMS Immunol Med Microbiol. 2005;44:137-42.
12. Solca NM, Bernasconi MV, Piffaretti JC. Mechanism of metronidazole resistance in Helicobacter pylori: comparison of the rdxA gene sequences in 30 strains. Antimicrob Agents Chemother. 2000;44:2207-10.
13. Paul R, Postius S, Melchers K, Schafer KP. Mutations of the Helicobacter pylori genes rdxA and pbp1 cause resistance against metronidazole and amoxicillin. Antimicrob Agents Chemother. 2001;45:962-5.
14. Chisholm SA , Owen RJ. Frameshift mutations in frxA occur frequently and do not provide a reliable marker for metronidazole resistance in UK isolates of Helicobacter pylori. J Med Microbiol. 2004;53:135-40.
15. Chisholm SA, Owen RJ. Mutations in Helicobacter pylori rdxA gene sequences may not contribute to metronidazole resistance. J Antimicrob Chemother. 2003;51:995-9.
16. Matteo MJ, Perez CV, Domingo MR, Olmos M, Sanchez C, Catalano M. DNA sequence analysis of rdxA and frxA from paired metronidazole-sensitive and -resistant Helicobacter pylori isolates obtained from patients with heteroresistance. Int J Antimicrob Agents. 2006;27:152-8.
17. Kim JJ, Reddy R, Lee M, Kim JG, El Zaatari FAK, Osato MS et al. Analysis of metronidazole, clarithromycine and tetracycline resistance of Helicobacter pylori isolates from Korea. J Antimicrob Chemother. 2001;47:459-61.
18. Mégraud F. Surveillance de la résistance de Helicobacter pylori aux antibiotiques. In: Surveillance nationale des maladies infectieuses 1998-2000.St Maurice, France:Institut de Veille Sanitaire,2003:327-9.
19. Toracchio S, Marzio L. Primary and secondary antibiotic resistance of Helicobacter pylori strains isolated in central Italy during the years 1998-2002. Dig Liver Dis. 2003;35:541-5.
20. Ortiz AP, Reis FC, Ferraz LFC, Gerrits MM, Mendonca S, Kusters JG et al. Differentially expresed genes in responses to amoxicillin in Helicobacter pylori analyzed by RNA arbitrarily primed PCR. FEMS Immunol Med Microbiol. 2006;50:226-30.
21. Gerrits MM, Godoy AP, Kuipers EJ, Ribeiro ML, Stoof J, Mendonca S et al. Multiple mutations in or adjacent to the conserved penicillin-binding protein motifs of the penicillin-binding protein 1A confer amoxicillin resistance to Helicobacter pylori. Helicobacter. 2006;11:181-7.
22. Gerrits MM, Schuijffel D, van Zwet AA, Kuipers EJ, Vandenbroucke-Grauls CM, Kusters JG. Alterations in penicillin-binding protein 1A confer resistance to beta-lactam antibiotics in Helicobacter pylori. Antimicrob Agents Chemother. 2002;46:2229-33.
23. Chopra I, Roberts M. Tetracycline Antibiotics: Mode of Action, Applications, Molecular Biology, and Epidemiology of Bacterial Resistance. Microb Molec Biol Reviews. 2001; 65:232-60.
24. Schnappinger D, Hillen W. Tetracyclines: antibiotic action, uptake and resistance mechanisms. Arch Microb. 1996;165:359-69.
25. Brodersen DE, Clemons WM, Carter AP, Morgan-Warren RJ, Wimberly BT, Ramakrishnan V. The structural basis for the action of the antibiotics tetracycline, pactamycin and hygromycin B on the 30S ribosoma subunit. Cell. 2000;103:1143-54.
26. Trieber CA,Taylor DE. Mutations in the 16S rRNA genes of Helicobacter pylori mediate resistance to tetracycline. J Bacteriol. 2002;184:2131-40.
27. Dailidiene D, Bertoli MT, Miciuleviciene J, Mukhopadhyay AK, Dailide G, Pascasio MA et al. Emergence of tetracycline resistance in Helicobacter pylori: multiple mutational changes in 16S ribosomal DNA and other genetic loci. Antimicrob Agents Chemother. 2002;46:3940-6.
28. Gerrits MM, Berning M, van Vliet AH, Kuipers EJ, Kusters JG. Effects of 16S rRNA gene mutations on tetracycline resistance in Helicobacter pylori. Antimicrob Agents Chemother. 2003;47:2984-6.
29. Li Y, Dannelly HK. Inactivation of the putative tetracycline resistance gene HP1165 in Helicobacter pylori led to loss of inducible tetracycline resistance. Arch Microbiol. 2006;185:255-62.
30. Wu JY, Kim JJ, Reddy R, Wang WM, Graham DY, Kwon DH. Tetracycline-resistant clinical Helicobacter pylori isolates with and without mutations in 16S rRNA-encoding genes. Antimicrob Agents Chemother. 2005;49:578-83.
31. Wu H, Shi XD, Wang HT, Liu JX. Resistance of Helicobacter pylori to metronidazole, tetracycline and amoxycillin. J Antimicrob Chemother. 2000;46:121-3.
32. Kwon DH, Kim JJ, Lee M, Yamaoka Y, Kato M, Osato MS et al. Isolation and characterization of tetracycline-resistant clinical isolates of Helicobacter pylori. Antimicrob Agents Chemother. 2000;44:3203-5.
33. Megraud F. H. pylori antibiotic resistance: prevalence, importance, and advances in testing. Gut. 2004;53:1374-84.
34. Peters DH, Clissold SP. Clarithromycin. A review of its antimicrobial activity, pharmacokinetic properties and therapeutic potential. Drug. 1992;44:117-64.
35. Versalovic J, Shortridge D, Kibler K, Griffy MV, Beyer J, Flamm RK et al. Mutations in 23S rRNA are associated with clarithromycin resistance in Helicobacter pylori. Antimicrob Agents Chemother. 1996;40:477-80.
36. Occhialini A, Urdaci M, Doucet-Populaire F, Bebear CM, Lamouliatte H, Megraud F. Macrolide resistance in Helicobacter pylori: rapid detection of point mutations and assays of macrolide binding to ribosomes. Antimicrob Agents Chemother. 1997;41:2724-8.
37. Oleastro M, Menard A, Santos A, Lamouliatte H, Monteiro L, Barthelemy P et al. Real-time PCR assay for rapid and accurate detection of point mutations conferring resistance to clarithromycin in Helicobacter pylori. J Clin Microbiol. 2003;41:397-402.
38. Glupczynski Y, Megraud F, Lopez-Brea M, Andersen LP. European multicenter survey of in vitro antimicrobial resistance in Helicobacter pylori. Eur J Clin Microbiol Infect Dis. 2001;20:820-3.
39. Vasquez A, Valdez Y, Gilman RH, McDonald JJ, Westblom TU, Berg D et al. Metronidazole and clarithromycin resistance in Helicobacter pylori determined by measuring MICs of antimicrobial agents in color indicator egg yolk agar in a miniwell format. The Gastrointestinal Physiology Working Group of Universidad Peruana Cayetano Heredia and the Johns Hopkins University. J Clin Microbiol. 1996;34:1232-4.
40. Kato S, Fujimura S, Udagawa H, Shimizu T, Maisawa S, Ozawa K et al. Antibiotic resistance of Helicobacter pylori strains in Japanese children. J Clin Microbiol. 2002;40: 649-53.
41. Cattoir V, Nectoux J, Lascols C, Deforges L, Delchier J, Megraud F, et al. Update on fluoroquinolone resistance in Helicobacter pylori: new mutations leading to resistance and first description of gyrA polymorphism associated with hypersusceptibility. J Antimicrobiol Agents. 2007;29:389-96.
42. Rispo A, Di Girolamo E, Cazzolino A, Bozzi R, Morante A, Pasquale L. Levofloxacin in first-line treatment of Helicobacter pylori infection. Helicobacter. 2007;12:364-5.
43. Rubinstein E. History of quinolones and Their Side Effects. Chemotherapy. 2001;47:3-8.
44. Bogaerts P, Berhin C, Nizet H, Glupczynski Y. Prevalence and mechanisms of resistance to fluoroquinolones in Helicobacter pylori strains from patients living in Belgium. Helicobacter. 2006;11:441-5.
45. Glocker E, Stueger H, Kist M. Quinolone resistance in Helicobacter pylori isolates in Germany. Antimicrob Agents Chemother. 2007;51(1):346-9.
46. Boyanova L, Koumanova R, Gergova G, Popota M, Mitov I, Kovacheva Y, et al. Prevalence of resistant Helicobacter pylori isolates in Bulgarian children. J Med Microbiol. 2002;51:786-90.
47. Megraud F. Basis for the management of drug-resistant Helicobacter pylori infection. Drugs. 2004;64(17):1893-904.
48. Glocker E, Berning M, Gerrits MM, Kusters JG, Kist M. Real-time PCR screening for 16S rRNA mutations associated with resistance to tetracycline in Helicobacter pylori. Antimicrob Agents Chemother. 2005;49:3166-70.
Recibido: 24 de junio de 2008.
Aprobado: 17 de noviembre de 2008.
Lic. Mayrín Hernández Power. Centro Nacional de Investigaciones Científicas. Departamento de Microbiología e Inmunología. Avenida 25 y 158 No.15202, Cubanacán, Playa, Ciudad de La Habana, Cuba. Apartado Postal 6414.


{kind=link}
{kind=link}
{kind=link}
{kind=link}