










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Salud Animal
versión impresa ISSN 0253-570X
Rev Salud Anim. v.29 n.1 La Habana ene.-abr. 2007
¿CÓMO SUPERAR LA MEMBRANA CELULAR?
HOW TO CROSS THE CELLULAR MEMBRANE?
Marlén González* y A. Vega**
Grupos de Biología Molecular* y Virología**. Dirección Microbiología. Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, La Habana, Cuba. Correo electrónico: marlen@censa.edu.cu
RESUMEN
Con el propósito de poder introducir moléculas en el interior de la célula, superando su barrera natural, se han desarrollado diferentes procedimientos de gran utilidad en los estudios de biología molecular y celular, entre los que podemos citar: Técnicas de permeabilización celular, Técnicas de Microinyección, Técnicas Transfección celular y Técnicas de Transducción. La elección de uno u otro método depende en gran medida del objetivo final del experimento y de la disponibilidad de equipamiento adecuado. En esta revisión se abordan los métodos de laboratorio empleados para la introducción de moléculas en el interior de las células y se discuten las ventajas y desventajas de los mismos.
Palabras clave: permeabilización celular; microinyección; transfección celular; transducción.
ABSTRACT
In order to introduce molecules into cells, different useful procedures have been developed in molecular and cellular biology studies. Among them we have: permeabilization, microinjection, cellular transfection, and transduction methods. The selection of the best method depends on the experiment final purpose and the equipment available. In this review we approach the laboratory methods used to introduce molecules into cells with a special emphasis in their advantages and disadvantages.
Key words: permeabilization; microinjection; cellular transfection; transduction.
INTRODUCCIÓN
En las últimas décadas se ha producido el desarrollo de una de las ramas más fascinantes de la biología: la tecnología del ADN recombinante, lo que ha permitido la manipulación del genoma, aumentando las expectativas en campos como la investigación, el diagnóstico y la terapéutica, lo que hace posible la transferencia genética (61; 62).
En muchas ocasiones, el desarrollo de estos procedimientos es necesario para la introducción de moléculas en el interior de las células, ya sea con el fin de estudiar el efecto que produce dicha molécula sobre la célula, o bien con el fin de la síntesis de otra (2). En este último caso, la célula es empleada como factoría de síntesis de moléculas, lo que ha requerido la optimización de los métodos de transferencia para poder aplicarlos con alta eficiencia y seguridad, así como con la menor dosis de invasividad posible (73).
Con el propósito de introducir moléculas en el interior de la célula, se han desarrollado diferentes procedimientos que permitan superar su barrera natural, la elección del método está en función del objetivo final del experimento y de la disponibilidad de equipamiento adecuado.
En esta revisión se abordan los métodos de laboratorio empleados para la introducción de moléculas en el interior de las células, así como se discute las ventajas y desventajas de los mismos.
Métodos de transferencia de material genético al interior de las células
La posibilidad de introducir ácido nucleico en el interior de las células ha permitido el desarrollo del conocimiento de la regulación génica y la función de las proteínas en células eucariotas, tejidos y organismos. Los primeros estudios realizados por Pagano y Vaheri (53), Graham y Van Der Eb (31), con las técnicas de transfección mediadas por DEAE dextrano y fosfato de calcio, abrieron el camino a los siguientes experimentos donde se necesitaba transferir ADN dentro de células.
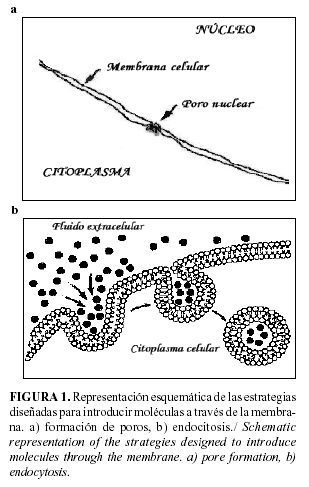
Las células vivas presentan una barrera a la libre difusión de moléculas, la membrana plasmática, la cual evita la pérdida de componentes fundamentales e impide la entrada de moléculas externas, por ello posibilita la existencia de gradientes de concentración de productos a través de la membrana.
Para poder superar esta barrera se han diseñado muchas estrategias experimentales, unas basadas en la formación de poros mas o menos permanentes (Figura 1 a), y las otras en la vehiculización de moléculas mediante el uso de las vías naturales de entrada de macromoléculas; la endocitosis (Figura 1 b) (60).
Entre las estrategias diseñadas podemos señalar las siguientes:
• Técnicas de permeabilización celular
• Técnicas de Microinyección
• Técnicas Transfección celular.
• Técnicas de Transducción
Técnicas de permeabilización celular:
Se trata de técnicas que permiten mantener las células vivas, inmersas en una solución, durante un periodo de tiempo que no suele superar las pocas horas, en condiciones tales que se produzca gran cantidad de orificios en la membrana. Cualquier molécula añadida a la solución que las rodea, penetra en la célula gracias al gradiente de concentración (60).
El medio que rodea la célula está formado por dos componentes importantes:
• Se requiere aportar externamente una fuente de energía (ATP), debido a que la pérdida de los gradientes electroquímicos de membrana impide que la célula regenere el ATP consumido.
• Un extracto citosólico procedente de sistemas homólogos o heterólogos. Su función es reducir en lo posible la pérdida de componentes vitales para el funcionamiento celular.
Estos sistemas de células permeabilizados son ampliamente empleados en estudios de identificación de factores solubles importantes en los procesos celulares, pero presenta dos grandes inconvenientes: la escasa duración del ciclo celular viable, y la pérdida incontrolable de componentes celulares que pueden ser vitales para el funcionamiento celular (a excepción de cuando se emplea el extracto citosólico de la propia célula) (60).
Frecuentemente se utiliza la técnica de electroporación para lograr la permeabilización de la membrana. Se basa en la aplicación de un elevado voltaje a las células durante un periodo de tiempo muy corto. Las células despolarizan sus membranas y se forman pequeños orificios por los que penetran las moléculas que se encuentran alrededor. Los impulsos eléctricos utilizados modifican transitoriamente las propiedades de la membrana celular, resultando en la permeabilización reversible de la célula (49). Pasada la despolarización, muchas células sufren daños irreparables y mueren (en muchos casos más del 90%), pero algunas (5 al 10%) se recuperan e incorporan las moléculas deseadas. La ventaja de esta técnica es que se aplica a millones de células a la vez y habitualmente se obtienen eficiencias de entrada de las moléculas del 100% en las células que sobreviven. Es una técnica que requiere su ajuste para cada tipo celular, a fin de determinar las condiciones óptimas de duración y potencia del pulso. Se busca siempre el mejor equilibrio entre las condiciones que aseguran la entrada en la célula y las que maximizan la viabilidad celular (60).
Técnicas de Microinyección:
La microinyección es una tecnología que permite la introducción mecánica de soluciones en el interior de la célula mediante una micropipeta que se controla con la ayuda de un micromanipulador, bajo un microscopio de alta resolución (Figura 2).
Es una técnica muy sensible, que requiere una gran especialización del personal que la realiza y un equipo delicado, sofisticado y costoso. Los instrumentos modernos de microinyección, manipulados por personal experto, permiten realizar algunas decenas de inyecciones por hora de trabajo, por lo que resulta ser un sistema tedioso, que permite manipular pocas docenas o escasos cientos de células con gran dificultad (60).
La fertilización por microinyección es una importante innovación surgida en los últimos años, que finalmente ha resultado en un tratamiento viable, incluso, en los casos más difíciles de esterilidad masculina (41).
Existen además sistemas de microinyección biológicos, como los virus, que inoculan su material genético dentro de la célula hospedadora, y es por
ello que son muy usados como vectores sobre sistemas de células de mamíferos (64; 73).
Técnicas de transfección celular:
Las técnicas de transfección son la imagen de las técnicas de transformación en procariotas, se basan en la inserción de material exógeno en el genoma de ese organismo (11; 26).
Estas técnicas han sido desarrolladas específicamente para la introducción de ácidos nucleicos dentro de las células, y han permito ampliar los conocimientos que se tenían sobre replicación, regulación génica y el funcionamiento de las células a nivel molecular, entre otras, que actualmente se aplican en el campo de la genética para la producción de plantas y animales transgénicos (8), líneas celulares modificadas (2; 17), y la construcción de ADN recombinante (27; 29; 80).
La introducción de una cadena de ADN recombinante en las que se ha situado una secuencia codificante, bajo una secuencia de regulación que se desea estudiar, permite medir con facilidad las tasas de expresión genética en diferentes situaciones experimentales. Por otro lado, la introducción de un plásmido con secuencias codificante y reguladora permite la producción de una proteína deseada. En este caso se emplea a la célula hospedadora como una "fábrica" para este producto (60). Es importante la selección de las células que han recibido el material genético incorporado, de las que no. Para ello se puede realizar un cultivo en medio selectivo, puesto que normalmente los plásmidos que se insertan incluyen también marcadores de selección (4; 60).
Existen diversos métodos para introducir y expresar genes en las células. Algunos se utilizan para la expresión transitoria o temporal de los genes. Estos permiten evaluar la transcripción o la replicación de genes transfectados varios días después de la introducción del ADN. Otros métodos permiten la expresión estable o permanente, al menos durante un extenso periodo, es útil en los experimentos donde se requiera la formación de líneas celulares que contengan genes que se integren en el ADN cromosomal (26; 79).
De forma general las técnicas de transfección se pueden dividir en dos grupos: Métodos químicos y Métodos físicos.
Métodos químicos de transfección celular: Son aquellos en los que se forman complejos para que las células sean capaces de aceptar o incorporar una cierta molécula a su citoplasma, por medio del mecanismo normal de endocitosis. Durante años se han desarrollado diversos reactivos de transfección con esta finalidad, todos con características propias. Entre ellos podemos citar el fosfato de calcio (9; 28), el DEAE dextrano (10) y la lipofección (45; 52). Los dos primeros se unen al ADN formando complejos asimilables por la célula, mientras que la lipofección se basa en el recubrimiento de la molécula de ADN en un complejo de lípidos capaces de atravesar la membrana.
Recientemente se han diseñado varios reactivos comerciales que facilitan los métodos químicos de transfección (36), como por ejemplo el juego de reactivos de formulación no liposomal FuGeneTM 6, que se caracteriza por producir altos niveles de transfección para una amplia variedad de líneas celulares eucariotas con mínima citotoxicidad (http:// biochem.boehringer-mannheim.com).
a) Método de fosfato de calcio:
Este procedimiento ha sido empleado por más de 20 años para la transferencia y expresión de información genética en células de mamíferos. Fue descrito por primera vez para la introducción de ADN de adenovirus en células de mamíferos por Graham y Van Der Eb en 1973 (31) y se ha convertido en una de las técnicas más populares para la transfección estable o transitoria de células de mamíferos, siendo estudiada en detalles por varios grupos en el mundo (9; 28).
Se basa en la obtención de un precipitado entre el cloruro de calcio y el ADN en una solución salina de fosfatos, formando unos agregados que son endocitados/fagocitados por las células. Aparentemente el agregado con calcio protege al ADN de la degradación por las nucleasas celulares.
El tamaño y la calidad del precipitado es crítico para el éxito del proceso, que se ve afectado por factores tales como pequeños cambios en el pH de la solución. Otros factores que intervienen en la eficiencia de la transfección son la cantidad de ADN en el precipitado, el tipo de célula a utilizar y el tiempo de exposición de las mismas al precipitado (28).
El método del fosfato de calcio ha sido utilizado en el análisis de la replicación del ADN y la funcionalidad de los promotores en experimentos transitorios y la utilización de líneas celulares establemente transfectadas. Tiene como ventajas que permite introducir ADN en células eucariotas de forma estable o transitoria, requiere de reactivos baratos y de uso frecuente, y un equipamiento tradicional para un laboratorio dedicado al trabajo con líneas celulares. Además, puede ser aplicado a una cantidad relativamente grande de células en placas de cultivo. Sin embargo está limitado por la proporción variable y frecuentemente baja de células que toman el ADN exógeno (28).
b) Método con DEAE dextrano:
El diethyl aminoethyl dextrano (DEAE dextrano) es un polímero catiónico derivado del dextran y su carga positiva le permite unirse a los enlaces aniónicos fosfodiester del ADN. Estos complejos formados entre el ADN y el dextrano mantienen su carga positiva, lo que le permite unirse a la superficie de la célula cargada negativamente y pasa al interior de la célula mediante endocitosis (70).
Este método fue empleado originalmente para facilitar la entrada de ARN de poliovirus a células (53) y posteriormente para introducir ADN del virus SV40 (47).
Tiene como ventajas su sencillez, reproducibilidad y bajo costo, además de que se requiere de pocas cantidades de ADN. Entre sus desventajas se puede señalar el número limitado de líneas celulares en las cuales se pueden obtener resultados eficientes y su limitación a transfecciones transientes de ADN en la célula huésped debido a la toxicidad del DEAE.
Este método ha sido utilizado en el estudio de expresión de proteínas recombinantes (10) y la transfección de genomas virales y plásmidos (7). Puede estar interferido por la concentración de DEAE dextrano, el tiempo de exposición, la concentración de ADN, y el número y tipo de células (20).
c) Método de Lipofección:
Se basa en la formación de complejos entre lípidos catiónicos y ADN, llamados liposomas. Dicho complejo tiene afinidad por la membrana y permite la entrada del ADN en el citosol.
La lipofección es una técnica muy efectiva aunque laboriosa. Se emplea en la introducción de ADN recombinante en las células embrionarias, en el proceso de obtención de animales transgénicos y en la terapia génica (52). Es fácilmente reproducible, se describe tanto para expresión transitoria como estable, resultando la expresión estable 5-100 veces más eficiente, que por el resto de los métodos químicos, y en la expresión estable se obtiene 3-20 veces mayor frecuencia de transfección. Tiene la habilidad de transfectar ciertos tipos de células donde no resultan otros métodos químicos (12; 13), capaz de introducir ADN de todas las tallas desde oligonucleótidos de cromosomas de levadura (40), introducción de ARN (46) y proteínas (14). A diferencia de los otros métodos químicos puede ser usado para transferir ADN y ARN in vivo a humanos y animales (18; 45).
En el año 1980, Fraley et al. (21), comenzaron a utilizar liposomas artificiales para introducir ADN en el interior de las células, ya en la actualidad existe un gran número de lípidos que se emplean en este método.
Es esencial optimizar las condiciones específicas de transfección para obtener buenas eficiencias, que suelen ser en buenas condiciones de entre el 70 y el 90% de las células de la placa. Las desventajas del método son, aparte del elevado precio de los lípidos, el hecho de que no todos los lípidos funcionan en todos los tipos celulares, y que hay que optimizar el ensayo para cada tipo celular. Los parámetros a optimizar son la relación de cargas entre el lípido y el ADN, la cantidad de ADN empleado, el tiempo que se exponen las células la complejo y la presencia o ausencia de suero (5).
Métodos físicos de transfección celular: Son aquellos en los que se produce una introducción mecánica del ADN como la microinyección (60), electroporación (33; 48), fusión de protoplastos (58) y bombardeo de microproyectiles recubiertos con ADN (41; 42).
a) Microinyección directa:
Esta técnica resulta ser muy efectiva, para la transfección celular, aunque laboriosa. Básicamente, se usa como un método para establecer líneas celulares que lleven copias integradas del ADN de interés. Es además, el método que se emplea en la introducción del ADN recombinante en las células embrionarias en el proceso de obtención de animales transgénicos (60).
Tiene como ventajas que no se exponen los polinucleótidos a los compartimentos celulares, tales como el endosoma (de bajo pH), es un método simple, económico, y no tóxico para las células, puede ser usado tanto en ensayos de transfección estables como transitorios, lográndose una alta eficiencia, donde una de cada cinco células incorporan el ADN de forma estable. Sin embargo, no puede ser empleado de forma rutinaria para introducir ADN a un gran número de células, como para permitir después análisis bioquímicos, y requiere de equipamientos, infraestructura y personal altamente entrenado (60).
b) Electroporación:
Es una técnica de introducción de ADN a células en cultivo que fue reportada por primera vez, por Wong y Neumann (83), para estudios de transfección de
genes. Se realiza mediante la aplicación de pulsos eléctricos a las células, lo que provoca la apertura de poros en la membrana de estas con la consiguiente penetración del ADN presente en el medio de electroporación. Conlleva a la formación de líneas celulares establemente transformadas con la integración de una sola copia del ADN foráneo (59; 67).
Las ventajas de esta técnica son que permite la transfección de líneas celulares recalcitrantes a otros métodos (se requiere de su optimización para cada tipo celular), se aplica a millones de células a la vez y habitualmente la eficiencia de transfección celular es del 100% de las células que sobreviven al pulso eléctrico.
En términos generales la mayor desventaja de la electroporación es su costo, ya que la mayoría de los equipos comercialmente disponibles son caros, en el orden de los miles de dólares, además de los accesorios como cubetas y adaptadores que también lo son (7).
La eficiencia de la transfección por electroporación está influenciada por una serie de factores, que necesitan ser optimizados para lograr los resultados deseados (7), entre estos se encuentran: la magnitud del campo eléctrico aplicado; la duración del pulso eléctrico; temperatura; concentración, pureza y tipo de ADN; y composición iónica del medio.
Ha sido utilizada para introducir ADN en una amplia variedad de células de animales (83), de plantas (22) y también de bacterias y levaduras (1), en la introducción de ADN foráneo a espermatozoides bovinos para su uso en transgénesis mediada por inseminación artificial (23) y transfección de células embrionarias totipotentes (30). Además, se ha adaptado como una herramienta para un amplio rango de investigaciones biomédicas y terapia humana, fundamentalmente en investigaciones y tratamiento del cáncer (25; 35; 44).
Recientemente, Haleem-Smith et al. (33) optimizaron una nueva variación de esta técnica llamada Nucleofección, donde el ADN plasmídico es transferido directamente en el interior del núcleo celular, independientemente del estado de crecimiento de la célula. Esta técnica muestra una alta eficiencia de transfección y una aceptable supervivencia celular, lo que la hace útil en estudios de expresión de genes en biología cardiovascular, y como paso hacia el desarrollo de las estrategias de terapia del gene (36).
c) Fusión de protoplastos:
Desarrollado por Schaffner en 1980 (66), el método consiste en obtener protoplastos de bacterias que llevan alto número de copias del plásmido de interés y se funde directamente con las células de mamíferos en cultivo.
A pesar de no ser de los métodos más eficientes para las líneas celulares normalmente usadas en los laboratorios, se ha utilizado tanto en transfecciones transitorias como estables, fundamentalmente, para aquellas líneas celulares en las cuales el proceso de endocitosis ocurre de forma ineficiente.
La gran ventaja del método radica en su eficiencia para ciertos tipos celulares, mientras que su desventaja es la gran cantidad de manipulaciones que conlleva, el tiempo de trabajo y que normalmente no son posibles las cotransfecciones, por lo que el gen de interés tiene que estar siempre contenido en un plásmido junto con el marcador de selección (65).
d) Bombardeo de microproyectiles recubiertos con ADN:
Este método de transfección celular consiste en la introducción del ADN adherido a micropartículas que se disparan a alta velocidad sobre las células (84). Ha sido utilizado de forma satisfactoria para introducir ácido nucleico a las células en cultivo de igual forma que ocurre in vivo (38).
El primer reporte de transfección de células de mamíferos empleando microproyectiles fue publicado por Zelenin et al. (86), al introducir el gen de la resistencia a neomicina en fibroblastos NIH/3T3. Otros reportes han aparecido en los últimos años que incluyen la transfección de ovocitos de peces (39), células humanas, fibroblastos de pollo, miotúbulos esqueléticos, células epiteliales y miocardiocitos (37). Recientemente, también se adaptó la metodología para células en suspensión, y se ha logrado transfectar células linfoides humanas (43).
Este se ha mostrado como un método efectivo de transferir genes tanto in vitro como in vivo (41; 42). Sin embargo, características de rigidez de los diferentes tejidos, la procedencia del ADN extraño, y la capacidad de transcripción intrínseca conducen a grandes variaciones en la eficiencia de la expresión de los genes en conjunto. Esto podría sugerir aplicaciones limitadas de esta tecnología en la terapia génica, pero sería útil para investigaciones en la expresión de construcciones de ADN en tejidos específicos.
Finalmente, un desarrollo factible de este método puede ser aplicado en su uso directo como parte de un protocolo de vacunación, como demuestran las investigaciones de Ulmer et al. (78), al realizar la inyección directa en el músculo, del gen que codifica la proteína de matriz del virus influenza A.
A pesar de lo avances alcanzados, la biobalística no es utilizada como una técnica de rutina, por su carestía, las dificultades para su aplicación y la existencia de otros métodos mucho mas simples y costeables.
e) Fotoporación laser-asistido:
Recientemente, se ha reportado el método de fotoporación láser-asistido (50; 71; 72) utilizando fuentes como la agitación continua de los lásers del argón-ion que funcionan a 488 nm (54; 68; 69), y más recientemente con los lasers del titanio-zafiro del femtosecond (76; 77; 85).
Consiste en la utilización de un rayo láser firmemente enfocado (con un volumen focal localizado de ~10-19 m3), que perfora la membrana celular, permitiendo que las moléculas se incorporen a la célula. La fluidez de la membrana facilita el cierre subsiguiente de la perforación.
A pesar de que las pruebas con los métodos de fotoporación han resultado satisfactorias, los sistemas del láser empleados son relativamente incómodos e implican típicamente procesos de dos fotones, los cuales requieren la colocación extremadamente exacta del foco del láser en la membrana celular. Tales técnicas han demostrado ser relativamente difíciles de alcanzar.
Como alternativa, más directa los procesos de un fotón han recibido menos atención en la literatura debido a los efectos potencialmente perjudiciales sobre la célula apuntada. Realizando la técnica con suficiente cuidado y control del rayo láser incidente, se ha demostrado un avance dominante en este campo, empleando un sistema altamente compacto y barato del láser. Al formar los poros en la membrana celular utilizando la vía de un solo fotón se logra una alta eficiencia y se evitan los daños celulares. Las células individuales transfectadas pueden mantener su capacidad de multiplicación en forma de clones que son utilizados para crear líneas celulares estables (69).
Una variante más reciente de este método es la transfección de células por el método de fotoporación con un láser de diodo violeta (57). Esta variante es fácil de poner en ejecución, particularmente si existe un microscopio óptico disponible, pues la fuente de láser es barata, compacta y portátil, en comparación con las fuentes previamente usadas para el fotoporador de láser. Las células se pueden seleccionar visualmente apuntado por el fotoporador, y utilizando valores bajos de energía en un corto periodo de tiempo, no causan daños irreversibles de la membrana de la célula.
Los genes reporteros en las transfecciones celulares
Cada uno de los procedimientos, analizados hasta el momento, tienen su propio espectro de ventajas y desventajas por lo que se deben seleccionar teniendo en cuenta la línea celular a utilizar, el objetivo final de la transfección, el gen a introducir, la economía del laboratorio y disponibilidad del equipamiento adecuado (45; 82).
Los métodos de transfección celular transitorios deben ser optimizados para cada línea celular y la vía más simple es utilizando un gen reportero o sea, secuencias que codifican para proteínas de simple detección, cuyo producto es fácilmente detectable por métodos analíticos no complejos y poco costosos, y permiten la cuantificación indirecta de la fortaleza de los promotores, así como la eficiencia de los sistemas de transfección.
En 1982, Gorman et al. (29), iniciaron los informes del concepto de gen reportero con el gen bacteriano de la cloranfenicol acetil transferasa (CAT). Los mas utilizados han sido el gen Lac Z de E. coli, que codifica para la b galactosidasa bacteriana (3; 19; 24), el gen de la cloranfenicol acetil transferasa (18), la hormona de crecimiento humana (34), el gen de la luciferasa de las luciérnagas (74), y los genes que codifican para las proteínas fluorescentes verde y azul de la medusa (33; 41; 48).
La proteína verde fluorescente (GFP) se ha convertido en una herramienta invaluable en las investigaciones contemporáneas de la biología celular, ya que su fluorescencia intrínseca puede ser visualizada en las células vivas. GFP se ha usado generalmente en dos vías: en una la expresión de un gen puede ser monitoreado fácilmente en la célula bajo condiciones variables, por la introducción de un transgen en el cual la secuencia del gen de interés está ligada al gen estructural del la GFP el que actúa como reportero (67); en la otra vía el gen estructural GPF se fusiona al gen de interés produciendo la fusión de las proteínas que están marcadas por la GFP, lo que permite evaluar las interacciones interproteínas (6; 81).
Otro grupo de marcadores o reporteros son aquellos que codifican para la resistencia a una droga. Mediante la selección de colonias resistentes a dicha droga se logran aislar clones independientes que han integrado este marcador de selección. Si bien estos marcadores permiten seleccionar aquellas células vivas que expresen el transgen, es preciso que la célula sobreviva en un ambiente tóxico por un periodo de tiempo generalmente largo. Por ello, estos marcadores son empleados sólo para transfecciones estables. El más comúnmente usado es el gen de la xantina-guanina fosforribosil transferasa (XGPRT, gpt) (4; 15).
• Técnicas de transducción
Las técnicas de transducción consisten en procesos en los cuales las células diana son infectadas con un virus que porta en su genoma la secuencia de interés (51). La introducción de ADN en las células mediante infección es un proceso más complejo que la transfección, precisando de un mayor número de pasos, y requiriendo además un mayor tiempo. No es de desdeñar el problema de seguridad biológica que implica la manipulación de ciertos virus. Además, la eficiencia de infección puede ser muy variable, extrema en células permisivas o muy baja en células resistentes (45).
Los vectores virales están hechos al menos de dos componentes, el genoma vírico modificado y la estructura del virión que lo rodea. La mayoría de los vectores de la generación presente comprenden partículas víricas basadas en gran manera en el tipo de estructura del virus sin cultivar. Esta estructura envuelve y protege al ácido nucleico viral y provee los medios para unirse y entrar en las células dianas. De todas formas, el ácido nucleico del virus en un vector diseñado para la terapia génica es transformado de muchas formas. Los objetivos de estos cambios son discapacitar el crecimiento del virus en las células diana, mientras se mantiene su capacidad para desarrollarse en forma de vector en el envoltorio asequible o células auxiliares; dar espacio en el genoma vírico para la inserción de secuencias de ADN exógeno e incorporar nuevas secuencias que codifiquen y capaciten la expresión apropiada del gen de interés. De esta forma, podemos considerar que los ácidos nucleicos del vector comprenden dos elementos: esas secuencias víricas esenciales que permanecen y la unidad de transcripción para el gen exógeno (61; 81).
Debido a que cualquier secuencia procedente del virus que codifica, podría ser expresada, incluso por un vector viral discapacitado, todas las secuencias víricas no esenciales deberían ser eliminadas. Así, idealmente, un vector contendrá sólo aquellas secuencias de ácido nucleico de virus necesariamente requeridas en cis. De todas formas, todas las demás funciones virales son necesarias para permitir la replicación del genoma del vector para los propósitos de producción y provisionamiento de las proteínas necesarias para ensamblar el virión. Idealmente un envase específico o célula auxiliar debería ser construido para que exprese aquellas funciones víricas capaces de funcionar en trans. En la práctica, la expresión de las proteínas estructurales del virus y de las proteínas implicadas en la replicación, a menudo resultan tóxicas para una línea celular, particularmente si el ciclo de vida del virus incluye una ayuda, entonces la infección productiva producirá la muerte celular más que una infección crónica. Así, la línea celular de envoltura apropiada requiere desarrollar muchos vectores idealizados que no existen (61).
Aunque casi cualquier genoma viral puede ser manipulado para llegar a ser recombinante, y adquirir la capacidad de expresar antígenos foráneos en la célula huésped, no todos los virus poseen la misma eficiencia para lograrlo. Muchas especies virales han sido utilizadas como sistemas de expresión de genes foráneos, siendo unas más eficientes que otras.
En la actualidad, ha sido común la utilización de vectores virales, por su capacidad de infectar un amplio rango de células y aceptar un amplio rango de regiones foráneas. Dentro de estos se encuentran los poxvirus, que incorporan el gen en una región no esencial de su genoma (27; 55; 80); los baculovirus, que infectan células de insectos (24; 32); los retrovirus que pueden expresar de forma permanente un gen foráneo en las células (16); y los adenovirus con sus versátiles aplicaciones, son generados con una mínima manipulación enzimática usando recombinación homóloga en bacterias y en células eucarióticas (5; 75); los lentivirus infectan muchas especies de mamíferos, incluyendo las vacas, caballos, ovejas, gatos, y primates, pueden infectar a las células que no se dividen como los macrófagos y en su genoma codifican proteínas regulatorias de la transcripción viral y el transporte del RNA viral (63).
El virus puede ser utilizado de forma simple como vector, por ejemplo el virus recombinante Vaccinia-rabia (56); además puede actuar como vector y vacuna, por ejemplo el virus capripox recombinante que expresa antígenos del virus de la peste de pequeños rumiantes y protege al animal contra las dos enfermedades (4). Entre estos, poxvirus y adenovirus son los más atractivos y eficientes vectores para ser usados con fines de vacunación (61; 79; 80).
Podemos concluir que el método ideal para la introducción de moléculas en el interior de las células debe caracterizarse por ser fácil de realizar, de bajo costo y lo suficientemente versátil como para ser utilizado en cualquier tipo de célula. Por lo que la selección de uno u otro método depende de varios factores, ya sea de las características de la línea celular, del objetivo que se persigue, la molécula a introducir, la economía del laboratorio y la disponibilidad de equipamiento adecuado.
En nuestro laboratorio se han utilizado frecuentemente los métodos químicos de transfección celular. Se optimizaron las condiciones de transfección de fribroblastos de embrión de pollo utilizando la metodología del precipitado de ADN-fosfato de calcio, para futuros ensayos de expresión temporal de genes en este tipo de células (28). Este método ha sido utilizado en la obtención de cepas virales recombinantes con el objetivo de su utilización como candidatos vacunales (27). Por otra parte se utilizó el método de transfección con DEAE dextrano en la evaluación de la expresión de genes de Anaplasma marginale (10) en un vector de expresión eucariota con vistas a su posterior utilización en nuevas estrategias de vacunación.
REFERENCIAS
1. Andreason GL, Evans GA. Introduction and expression of DNA molecules in eukariotic cell by electroporation. BioTechniques. 1988;6:50-53.
2. Arias AA, Dinauer MC, Ding J, Matute JD, Patiño PJ. Expresión y actividad de posibles polimorfismos provenientes de individuos normales en la proteína de 67 kd del sistema NADPH oxidasa utilizando el sistema COSphox. Biomédica.2004;24:262-72.
3. Aspden K, Passmore J, Tiedt F, Williamson A. Evaluation of lumpy skin disease virus, a capripoxvirus, as a replication-deficient vaccine vector. J Gen Virol. 2003;84:1985-1996.
4. Berhe G, Minet C, Le Goff C, Barrett T, Ngangnou A, Grillet C, et al. Development of a dual recombinant vaccine to protect small ruminants against peste-des-petits-ruminants virus and capripoxvirus infections. J Virol. 2003;77:1571-1577.
5. Bertelmann E, Ritter T, Vogt K, Reszka R, Hartmann C, Pleyer U. Efficiency of cytokine gene transfer in corneal endothelial cells and organ-cultured corneas mediated by liposomal vehicles and recombinant adenovirus. Ophthalmic Res. 2003;35 (2):117-124 (DOI: 10.1159/000069126).
6. Brackenbury B. Green fluorescent protein. Presented at the ASCB Symposium at the National Association of Biology Teacher Meeting.2002 http://www.ascb.org/teachers/green.html
7. Castro FO, Portelles Y. Transfección de adn a células de mamíferos. Biotecnología Aplicada. 1997;14(3):149-161.
8. Chang K, Qian J, Jiang M, Liu Y, Wu M, Chen Ch. Effective generation of transgenic pigs and mice by linker based sperm-mediated gene transfer. Biotechnology 2002;2:5 http://www.biomedcentral.com/1472-6750/2/5
9. Chowdhury EH, Kunou M, Nagaoka M, Kundu AK, Hoshiba T, Akaike T. High-efficiency gene delivery for expression in mammalian cells by nanoprecipitates of Ca-Mg phosphate. Gene. 2004;341:77-82.
10.Corona Belkis, Camacho Minerva, González Marlén, Martínez Siomara. Clonaje del gen msp1ß de Anaplasma marginale en un vector de expresión eucariota. Rev Salud Anim. 2001;23(1):69-72.
11.Cruz JJ. Terapia Génica: Definición. TERAPIA GENICA. FEFOC.2005 http://www.fefoc.org/educacion/rincon/terapiagenica.jsp
12.Daldrup-Link HE, Rudelius M, Oostendorp RA, Settles M, Piontek G, Metz S, et al. Targeting of hematopoietic progenitor cells with MR contrast agents. Radiology. 2003;228(3):760-767.
13.Daldrup-Link HE, Rudelius M, Piontek G, Metz S, Brauer R, Debus G. Migration of iron oxide-labeled human hematopoietic progenitor cells in a mouse model: in vivo monitoring with 1.5-T MR imaging equipment. Radiology. 2005;234(1):197-205.
14.Debs RJ, Freedman LP, Edmunds S, Gaensler KL, Duzgunes N, Yamamoto KR. Regulation of gene expression in vivo by liposome-mediated delivery of a purified transcription factor. J Biol Chem. 1990;265(18):10189-92.
15.Diallo A, Minet C, Berhe G, Le Goff C, Blank DN, Fleming M. et al. Goat immune response to capripox vaccine expressing the hemagglutinin protein of peste des petits rumiants. Ann N Y Acad Sci. 2002;969:88-91.
16.Eglitia MA, Anderson WF. Retroviral vectors for introduction of genes into mammalian cells. Biotechniques. 1988;6:608-614.
17.Faussner A, Bauer A, Kalatskaya I, Jochum M, Fritz H. Expression levels strongly affect ligand-induced sequestration of B2 bradykinin receptors in transfected cells. Am J Physiol Heart Circ Physiol. 2003;284(6):H1892-8.
18.Felgner PL, Tsai YJ, Sukhu L, Wheeler CJ, Manthorpe M, Marshall J. Improved cationic lipid formulations for in vivo gene therapy. Ann New York Acad Sci. 1995;772(1):126-139.
19.Fernández A, Gónnzález Marlén, Vega A, Espinosa Ivette. Niveles altos de expresión de b-galactosidasa bajo el control del promotor del citomelagovirus (pCMV) en cultivos secundarios de fibroblasto de embrión de pollo. Rev Salud Anim. 2000;22(2): 124-126.
20.Fernández A, Vega A, Coroas Laura, Cuello Sandra Influencia de la concentración de DEAE-Dextrano en la transfección de fibroblastos de embrión de pollo. Rev Salud Anim. 1996;18(3):181-183.
21.Fraley R, Subramani S, Berg P, Papahadjopolous D. Introduction of liposome-encapsulated SV40 DNA into cells. J Biol Chem. 1980;255:10431-35.
22.Fromm M, Taylor LP, Walbot V. Expression of genes transferred into monocot and dicot plant cells by electroporation. Proc Natl Acad Sci. 1985;82:5824-31.
23.Gagne M, Pothier F, Sirard H Electroporation of bovine spermatozoa to carry foreign DNA in oocytes. Molec Reprod Develop. 1991;29:6-15.
24.Gao R, McCormock CJ, Arthur MJ, Ruddell R, Oakley F, Smart DE, et al. High efficiency gene transfer into cultured primary rat and human hepatic stellate cells using baculovirus vectors. Liver. 2002;22(1):15-22.
25.Gershan JA, Johnson BD, Weber J, Schauer DW, Behnke S, Burns K, et al. Immediate transfection of patient-derived leukemia: a novel source for generating cell-based vaccines. Genet Vaccines Ther. 3: 4. Published on line 2005 June 21. doi: 10.1186/1479-0556-3-4. Copyright © 2005 Gershan et al; licensee BioMed Central Ltd.
26.González Marlén, Coroas Laura. Expresión de genes recombinantes en células de mamíferos. Reseña Bibliográfica. Rev. Salud Anim. 2002;24(1):1-10.
27.González Marlén, Coroas Laura, Fernández A, Noda Julia, Perera Carmen L, Ramos Edenis.et al. Obtención de una cepa viral recombinante Fowlpox-Gumboro. Rev. Salud Anim. 2003;25(1):20-26.
28.González Marlén, Espinosa Ivette, Vega A, Fernández A. Optimización de los parámetros que influyen en la transfección de fibroblastos de embrión de pollo mediante la metodología de la formación de precipitado ADN-fosfato de calcio. Rev. Salud Anim. 2000;22(2):89-92.
29.Gorman CM, Moffat LF, Howard BH. Recombinant genomes which express chloramphenicol acetyltransferase in mammalian cells. Mol Cell Biol. 1982;2:1044-1051.
30.Gossler A, Doetschman T, Korn R, Serfling E, Kenler R. Transgenesis by means of blastocysts-derived embryonic stem cell lines. Proc Natl Acad Sci. 1986;83:9065-9069.
31.Graham FL, Van Der Eb AJ. A new technique for the assay of infectivity of human adenovirus 5 DNA. Virology. 1973;52:456-461.
32.Gray CE, Coates CJ. High-level gene expression in Aedes albopictus cells using a baculovirus Hr3 enhancer and IE1 transactivator BMC. Mol Biol. 2004;5:1-8 (http://www.biomedcentral.com/ content/pdf/1471-2199-5-8.pdf).
33.HaleemSmith H, Derfoul A, Okafor C, Tuli R, Olsen D, Hall DJ, et al. Optimization of high-efficiency transfection of adult human mesenchymal stem cells in vitro. Mol Biotechnol. 2005;30(1):9-20.
34.Hernández O, Guillen I, Estrada MP, Cabrera E, Pimentel R, Pina JC, et al. Characterization of transgenic tilapia lines with different ectopic expression of tilapia growth hormone. Mol Mar Biol Biotechnol. 1997;6(4):364-375.
35.Hui SW. The application of electroporation to transfect hematopoietic cells and to deliver drugs and vaccines transcutaneously for cancer treatment. Technol Cancer Res Treat. 2002;1(5):373-384.
36.Iversen N, Birkenes B, Torsdalen K, Djurovic S. Electroporation by nucleofector is the best nonviral transfection technique in human endothelial and smooth muscle cells. Genetic Vaccines and Therapy. 3:2. doi: 10.2005;1186/1479-0556-3-2.
37.Johnston SA. Bioballistic transformation: microbes to mice. Nature. 1990;346:776-777.
38.Klein TM, Wolf ED, Wu , Sanford JC. High velocity microprojectiles for delivering nucleic acids into living cells. Nature. 1987;327:70-73.
39.Kolesnikov VA, Alimov AA, Barmintsev AA, Benyumov AO, Zelenina IA, Kraasnov AM et al. High velocity mechanical injection for foreign DNA into fish eggs. Genetika (Moskva). 1990;26:2122-2126.
40.Lamb BT, Gearhart JD. YAC transgenics and the study of genetics and human disease. Curr Opin Genet Dev. 1995;5:342-348.
41.Lauritzen HP, Reynet C, Schjerling P, Ralston E, Thomas S, Galbo H, et al. Gene gun bombardment-mediated expression and translo-cation of EGFP-tagged GLUT4 in skeletal muscle fibres in vivo. Pflugers Arch. 2002;444(6):710-721.
42.Leclerq S, Harms JS, Rosinha GM, Azevedo V, Oliveira SC. Induction of a Th1-type of immune response but not protective immunity by intramuscular DNA immunization with Brucella abortus GroEL heat-shock gene. J Med Microbiol. 2002;51:20-26.
43.Li C-Ch, Ruscetli FW, Rice NR, Chen E, Yang NS, Mikovits J, et al. Differential expression of Rel family members in human T-cell leukemia type-I infected cells. J Virol. 1993;67:4205-4213.
44.Li L, Shivakumar R, Feller S, Allen C, Weiss JM, Dzekunov S, Singh V et al. Highly Efficient, Large Volume Flow Electroporation. Technology in Cancer Research & Treatment. 2002;1(5):341-349.
45.Lundqvist A, Noffz G, Pavlenko M, Saeboe-Larssen S, Fong T, Maitland N, et al. Nonviral and viral gene transfer into different subsets of human dendritic cells yield comparable efficiency of transfection. J Immunother. 2002;25(6):445-454.
46.Malone R, Felgner PL, Verma I. Lipofectin-mediated RNA transfection. Proc Natl Acad Sci. 1989;86:6077-6081.
47.McCutchan JH, Pagano JS. Enhancement of the infectivity of simian virus 40 deoxyribonucleic acid with diethyl aminoethyl-dextran. J Natl Cancer Inst. 1968;41:351-359.
48.Micka B, Trojaneck B, Niemitz S, Lefterova P, Kruopis S, Huhn D, et al. Comparison of non-viral transfection methods in melanoma cell primary cultures. Cytokine. 2000;12(6):828-833.
49.Mir LM. Protocolo europeo de investigación en electroquimioterapia. Oncología. 2004;27(10):49-51. ISSN 0378-4835.
50.Mohanty SK, Sharma M, Gupta PK. Laser-assisted microinjection into targeted animal cells. Biotechnol Lett. 2003;25:895-899.
51.Nakamura Y, Sakudo A, Saeki K, Kaneko T, Matsumoto Y, Toniolo A, et al. Transfection of prion protein gene suppresses coxsackievirus B3 replication in prion protein gene-deficient cells. J Gen Virol. 2003;84:3495-3502.
52.Nakanishi K, Uenoyama M, Tomita N, Morishita R, Kaneda Y, Ogihara T, et al. Gene transfer of human hepatocyte growth factor into rat skin wounds mediated by liposomes coated with the sendai virus (Hemagglutinating Virus of Japan). Am J Pathol. 2002;161:1761-1772.
53.Pagano S, Vaheri A. Enhancement of infectivity of poliovirus RNA with diethylaminoethyl-dextran (DEAE-D). Arch Gesamte Virusforsch. 1965;17(3):456-464.
54.Palumbo G, Caruso M, Crescenzi E, Tecce MF, Roberti G, Colasanti A. Targeted gene transfer in eucaryotic cells by dye-assisted laser optoporation. J. Photochem Photobiol B-Biol. 1996;36:41-46.
55.Pancholi P, Perkus M, Tricoche N, Liu Q, Prince AM. DNA immunization with Hepatitis C virus (hcv) polycistronic genes or immunization by hcv dna priming-recombinant canarypox virus boosting induces immune responses and protection from recombinant hcv-vaccinia virus infection in hla-a2.1-transgenic mice. J Virol. 2003;77(1):382-390.
56.Paoletti E. Applications of pox virus vectors to vaccination: An update. Proc Nat Acad Sci. 1996;93:11349-11353.
57.Paterson L, Agate B, Comrie M, Ferguson R, Lake TK, Morris JE, et al. Photoporation and cell transfection using a violet diode laser. Optics express. 2005;13(2):595-600.
58.Rassoulzadegan M, Binetruy B, Cuzin F. High frequency of gene transfer after fusion between bacteria and eukaryotic cells. Nature. 1982;295:257-259.
59.Ray FA. Electroporation of plasmid DNA into normal human fibroblasts. In: Nickoloff, JA, editor. Animal Cell Electroporation and electrofusion protocols. Meth Molecular Biol. 1995;48:133-140.
60.Reina M. Técnicas de introducción de moléculas en las células. Universidad de Barcelona, Departamento de Biología Celular, http://bio.ub.es/biocel/bc/sub/ transfeccion.htm. Material docente diseñado y elaborado por Manuel Reina. Última actualización: 21/10/2003.
61.Rocha D, Costa B,Vieira A, Bruña-Romero O. Recombinant viruses as tools to induce protective cellular immunity against infectious diseases Carolina. Int Microbiol. 2004;7(2):83-94.
62.Rozalén J, Ceña V, Jordán J. Terapia Génica. Vectores de expresión. OFFARM. 2003;22(8):102-108.
63.Rubinson DA, Dillon CP, Kwiatkowski AV, Sievers C, Yang L, Kopinja J, et al. A lentivirus-based system to functionally silence genes in primary mammalian cells, stem cells and transgenic mice by RNA interference. Nature Genet. 2003;33:401-406.
64.Salaün P, Boulben S, Mulner-Lorillon O, Bellé R, Sonenberg N, Morales J. Embryonic-stage-dependent changes in the level of eIF4E-binding proteins during early development of sea urchin embryos. J Cell Sci. 2005;118:1385-1394.
65.Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: a laboratory manual, 2nd edn. Cold Spring Harbor, NY: Cold Spring Harbor Laboratry Press. 1989.
66.Schaffner W. Direct transfer of cloned genes from bacteria to mammalian cells. Proc Natl Acad Sci. 1980;77(4):2163-2167.
67.Schakowski F, Buttgereit P, Mazur M, Märten A, Schöttker B, Gorschlüter M. Novel non-viral method for transfection of primary leukemia cells and cell lines. Gen Vacci Ther. 2004;2(1):1-11. http://www.gvt-journal.com/content/2/1/1
68.Schneckenburger H, Hendinger A, Sailer R, Strauss WS, Schmitt M. Laser-assisted optoporation of single cells. J Biomed Opt. 2002;7(3):410-416.
69.Schneckenburger H, Wagner M, Kretzschmar M, Strauss WS, Sailer R. Laser-assisted fluorescence microscopy for measuring cell membrane dynamics. Photochem Photobiol Sci. 2004;3(8):817-22.
70.Selden RF, Burke-Howie K, Rowe ME, Goodman HM, Moore DD. Human growth hormone as a reporter gene in regulation studies employing transient gene expression. Mol Cell Biol. 1983;6:3173-3179.
71.Shirahata Y, Ohkohchi N, Itagak H, Satomi S. New technique for gene transfection using laser irradiation. J Invest Med. 2001;49:184-190.
72.Soughayer JS, Krasieva T, Jacobson SC, Ramsey JM, Tromberg B, Allbritton NL. Characterization of cellular optoporation with distance. Anal Chem 2000;72:1342-1347.
73.Talavera A, Sánchez H. Nuevos sistemas virales en terapia génica. Artículo de revisión. Virología. 2004;10(1):1-16.
74.Taniyama Y, Tachibana K, Hiraoka K, Namba Y, Yamasaki K, Hashiya N. Local delivery of plasmid DNA into rat carotid artery using ultrasound. Basic Science Reports Circulation. 2002;105:1233. (doi:10.1161/hc1002. 105228).
75.Tarturro F. Recombinant adenovirus-mediated cytotixic gene therapy of lymphoproliferative disorders: is CAR important for the vector to ride. Gene Ther. 2003;10(2):100-104.
76.Tirlapur UK, Konig K. Femtosecond near-infrared laser pulses as a versatile non- invasive tool for intra-tissue nanoprocessing in plants without compromising viability. Plant J. 2002;31:365-374.
77.Tirlapur UK, Konig K. Targeted transfection by femtosecond laser. Nature. 2002;418:290-291.
78.Ulmer JB, Donnelly JJ, Parker SE, Rhodes GH, Felgner PL, Dwarki VJ et al Heterologous protection against influenza by injection of DNA encoding a viral protein. Science. 1993;259(5102):1745-1749.
79.Vázquez D. Los poxvirus como vectores vivos: una promesa en la vacunación Biotecnología Aplicada. 1998;15(1):1-14.
80.Vázquez D, González S, Duarte C. Effect of promoters on cellular immune response induced by recombinant fowlpox virus expressing multi-epitope polypeptides from HIV-1 Biotechnol Appl Biochem. 2002;36:171-179.
81.Wallace DB, Viljoen GJ Immune responses to recombinants of the South African vaccine strain of lumpy skin disease virus generated by using thymidine kinase gene insertion. Vaccine. 2005;23(23):3061-3067.
82.Walters DK, Jelinek DF. The effectiveness of double-stranded short inhibitory RNAs (siRNAs) may depend on the method of transfection. Antisense Nucleic Acid Drug Dev. 2002;12(6):411-418.
83.Wong TK, Neumann E. Electric field mediated gene transfer. Biochem Biophys Res Commun. 1982;107:584-587.
84.Ye GN, Daniell H, Sanford JC. Optimization of delivery of foreign DNA into higher-plant chloroplasts. Plant Molecular Biology. 1990;15:809-819.
85.Zeira E, Manevitch A, Khatchatouriants A, Pappo O, Hyam E, Darash-Yahana M, et al. Femtosecond Infrared Laser - An Efficient and Safe in Vivo Gene Delivery System for Prolonged Expression. Mol Therapy. 2003;8:342-350.
86.Zelenin AV, Titomirov AV, Kolesnikov VA. Genetic transformation of mouse cultured cells with the help of high-velocity mechanical DNA injection. FEBS Lett. 1989;244:65-66.
(Recibido 17-11-2005; Aceptado 5-6-2006)


{kind=link}
{kind=link}