










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Salud Animal
versión impresa ISSN 0253-570X
Rev Salud Anim. v.30 n.2 La Habana Mayo-ago. 2008
Trabajo original
AISLAMIENTO DEL VIRUS DE LA GASTROENTERITIS TRANSMISIBLE DEL CERDO EN CULTIVOS CELULARES
ISOLATION OF SWINE TRANSMISSIBLE GASTROENTERITIS VIRUS IN CELL CULTURES
Edisleidy Rodríguez*, A. Betancourt**, Ana María Acevedo*, Damarys Relova*, J. Ayala* y Maritza Barrera*
*Grupo Virología Animal, Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de Las Lajas, La Habana, Cuba. **Facultad de Medicina Veterinaria. Universidad Agraria de la Habana. (UNAH). Cuba. Correo electrónico: batista@censa.edu.cu
RESUMEN
La gastroenteritis transmisible (TGE) es una enfermedad de los cerdos, infecciosa, aguda y altamente contagiosa; causada por un virus de la familia Coronaviridae, género Coronavirus. En nuestro país, a partir del año 2003 se comenzaron a presentar brotes de síndrome diarreico con las características clínico epizotiológicas de la enfermedad de TGE, pero para lograr la confirmación del diagnóstico era necesario aislar el virus en cultivos celulares a partir de animales enfermos, seguido de una identificación en aquellos cultivos que presentaran efecto citopático, para lo cual era necesario desarrollar una metodología para el aislamiento viral, ya que para este virus ha sido reportado con anterioridad que el aislamiento en células resulta muy difícil de lograr. En esta investigación se desarrolló una metodología para el aislamiento del virus de TGE. Los aislamientos obtenidos durante la evaluación de la técnica fueron identificados como positivos a través de la aplicación de inmunoperoxidasa indirecta sobre las células infectadas con un anticuerpo monoclonal comercial específico anti TGE.
Palabras clave: gastroenteritis transmisible del cerdo; TGE; aislamiento viral
ABSTRACT
Transmissible gastroenteritis (TGE) is an infectious, acute and highly contagious disease in swine. It is caused by a virus belonging to the Coronaviridae family, Coronavirus genus. Some outbreaks starting with diarrheic symptoms with the clinical characteristics of TGE disease have been present in our country since 2003. But for having the confirmation of the diagnostic, it was necessary to isolate the virus in cell cultures from sick animals, followed by an identification in those cell cultures with cytopatic effect. That is why, it was necessary to develop a methodology for the virus isolation, since for this virus, it has been previously reported that isolation in cells is very difficult to obtain. In this research, a methodology was developed for the virus isolation. The isolates obtained during the technique evaluation were identified as positive by indirect immunoperoxidase in cells with an anti TGE specific commercial monoclonal antibody.
Key words: transmissible gastroenteritis virus; TGE; virus isolation
INTRODUCCIÓN
La gastroenteritis transmisible del cerdo (TGE) es una enfermedad de los cerdos, infecciosa, aguda y altamente contagiosa; causada por un virus de la familia Coronaviridae, género Coronavirus (2). La misma se caracteriza por diarreas profusas y vómitos, provocando una alta morbilidad y letalidad en cerditos lactantes (8). Debido a las afectaciones que produce en la industria porcina ha sido considerada una de las enfermedades de declaración obligatoria por la Organización Mundial de Salud Animal (OIE)(4). El virus de TGE está presente en muchos países de Europa, América, Asia y África (7).
De acuerdo a los monitoreos serológicos conducidos en los Estados Unidos de América y Europa existe una alta prevalencia de este virus (6, 9). En Cuba, nunca se había presentado brotes de síndrome diarreico con las características clínico epizotiológicas, tales que sustentarán el emprender estudios serológicos para descartar o no la presencia de este virus u otro miembro de la familia. Sin embargo, a partir de febrero del 2003, se comenzaron a presentar en la provincia de La Habana cuadros de diarrea y vómitos en crías recién nacidas y diarrea, vómitos y agalactia en las puercas madres, con índices de mortalidad cercanos al 100% en las crías. Estos síntomas fueron asociados por las evidencias clínico epizootiológicas con la presencia de TGE. Posteriormente, se tuvieron evidencias por microscopía electrónica y serológicas, pero era necesario realizar la confirmación definitiva de la participación de este virus como agente causal del síndrome diarreico presentado en los cerditos.
Dentro de los métodos de diagnóstico confirmatorio de las enfermedades virales se encuentra el aislamiento viral a partir de cultivos celulares, principalmente debido a que la TGE era una enfermedad exótica para nuestro país. A nivel mundial se informa que este virus es extremadamente difícil de aislar (1). En Cuba no se contaba con experiencia previa de trabajo con este género viral, ni con una metodología precisa para el aislamiento de esta entidad. Por estas razones el objetivo de este trabajo fue evaluar una metodología para el aislamiento del virus de TGE, a partir de cultivo celulares, realizando algunas modificaciones al método normalizado en nuestro laboratorio para otros virus y la metodología descrita en el Manual de la OIE, 2004 (3) para este virus.
MATERIALES Y MÉTODOS
Reproducción experimental: Se inoculó una cerda recién parida y su camada de 3 cerditos por vía oral, con 2 mL de una suspensión de macerado de intestinos obtenidos a partir de crías con signos clínicos y sacrificadas a tal efecto. El inóculo fue tratado con antibióticos y filtrada (0.22µm).
Cultivos Celulares: Se utilizaron cultivos primarios de riñón de cerdo neonato y las líneas celulares: ST (testículo de cerdo) y SK6 (riñón de cerdo) procedentes de la Universidad de Guelph, Canadá y Universidad de Giessen, Alemania respectivamente. Las células se sembraron en placas de 24 pozos Nunc y para su crecimiento se utilizó el medio DMEM (Dulbeco's modified Eagle's medium) (Sigma-Aldrich), suplementado al 10% con suero fetal bovino. Los cultivos se utilizaron de dos formas: a las 24 horas de sembrados cuando la monocapa de células ya estaba completa y en fase exponencial de crecimiento. Como medio de mantenimiento para las células se usó medio DMEM, suplementado con antibióticos (Penicilina 100UI/mL; Estreptomicina 100 µg/mL; Fungizone 2,5 µg/mL)(SIGMA) y HEPES 25mM. Se evaluó el uso en la hora de adsorción de tripsina 10µg/mL (5) y sin tripsina.
Procesamiento de las muestras: Se analizaron las heces fecales diarreicas presentes en los cerditos inoculados a las 24 y 48 horas post inoculación (pi), las cuales fueron homogenizadas 1 gramo en 10 mL de medio DMEM sin suero, suplementado con antibióticos (3 veces la concentración descrita anteriormente), posteriormente se centrifugaron y filtraron a través de una membrana de acelato de cleulosa de 0,45 µm (3). Dentro del procesamiento de las muestras se evaluó la sonicación, para esto se utilizaron muestras en paralelo, unas se sonicaron después del paso de homogenización y otras fueron inoculadas sin sonicar.
Inoculación de los cultivos: Las muestras de heces fecales procesadas fueron inoculadas en tres réplicas cada caso. Tanto en las monocapas con una confluencia al 100% (24 horas) como en los cultivos en fase exponencial de crecimiento, el volumen del inóculo fue 200 µL y el caso específico de la inoculación en monocapa se evaluó la adición de 300 µL de medio de mantenimiento con tripsina o sin tripsina durante el tiempo de adsorción. Posteriormente se retiró el inóculo, se lavó con medio sin tripsina y se añadió 1 mL de medio sin tripsina.
Identificación de los aislamientos: Las células infectadas y no infectadas se fijaron luego de la aparición del efecto citopático con acetona al 30% y se analizaron mediante la técnica de inmunoperoxidasa indirecta (IPI), para detectar la presencia de antígenos específicos del virus de TGE. Para esto se utilizó el anticuerpo monoclonal 1DB12 comercializado por INGENASA y un antisuero policlonal de cerdo obtenido en nuestro laboratorio. Como 2dos anticuerpos se emplearon Anti IgG de ratón (Sigma) (dilución 1:400) para el monoclonal y Proteina A peroxidasa (Sigma) (dilución 1: 2000) en el caso del suero policlonal. Como sustrato cromógeno se utilizó una solución de Aminoetilcarbazol (AEC) (1 mL de 0,4% de AEC en N,N-dimetil-formamida, 19 mL de acetato de sodio 0,05 M pH 5,0) y peróxido de hidrógeno al 0,01%)..
RESULTADOS Y DISCUSIÓN
A las 18 horas post-inoculación (pi), se presentó en las crías recién nacidas un cuadro gastroentérico, que fue agravándose hasta llevar a los animales a deshidratación severa y muerte en el término de 5 días pi. La cerda madre presentó vómitos y agalactia, recuperándose a los 5 días pi.
Los cultivos primarios de riñón de cerdo inoculados con las muestras de heces fecales diarreicas obtenidas a las 24 y 48 horas pi de los cerditos enfermos, mostraron la formación de sincitios a las 12 horas de inoculadas las células (Fig. 1). A las 15 a 18 horas después de la inoculación estas aparecieron agrupadas y en la mayoría de los pozos la monocapa fue totalmente destruida a las 24 horas.
En el manual de técnicas diagnósticas y vacunas para animales terrestres (3), se describe una metodología para el aislamiento de este virus en cultivo de células. En este capítulo, se refiere la aparición del efecto citopático entre 3 y 7 días pi de los cultivos primarios, aspecto que debe ser señalado, pues en nuestro caso, este efecto se pudo observar tan temprano como a las 12 horas pi, el cual se completa a las 24 horas pi.
Durante el proceso de evaluación y estandarización de la metodología de aislamiento se obtuvo como resultado el aislamiento viral de las muestras de diarrea de las 48 horas, lo mismo sonicadas que sin sonicar, y en presencia de tripsina en la hora de adsorción. La presencia de tripsina en el medio de mantenimiento fue fundamental para lograr el aislamiento viral, ya que sin tripsina, se aisló el virus solamente de una muestra. Se plantea que la función de la tripsina es facilitar la penetración del virus a través de la membrana. La adición de tripsina al medio para el aislamiento del virus de TGE y otros como el rotavirus ha sido empleada por Paton y Lowings, 1997. En el caso de TGEv existe una metodología recomendada por el manual de la OIE (3) para su aislamiento, en la cual sugieren el empleo de la tripsina, pero no se describen el momento ni las concentraciones que se deben emplear. De manera general, a nivel mundial, los virus que forman este género, han sido reconocidos como muy difíciles de aislar (5, 8, 10).
En los cultivos en fase exponencial de crecimiento se observó un efecto tóxico debido a los lavados y la concentración de antibióticos, sobre todo en presencia de tripsina, por lo que se determinó que el uso de los cultivos en forma de monocapa establecida es indispensable para lograr el aislamiento de este virus.
Se demostró que la incubación para la adsorción del virus a las células, debe realizarse durante 1 hora, ya que con dos horas se logró el aislamiento viral, sin embargo, la monocapa de células presentó zonas destruidas.
La evaluación de las líneas celulares SK6 y ST, evidenció que estas son menos sensibles en el proceso de aislamiento viral que el cultivo primario, ya que solamente una muestra presentó efecto citopático en ambas líneas y el mismo apareció 24 horas después, que en los cultivos primarios donde se evidencia entre las 12 a 24 horas pi.
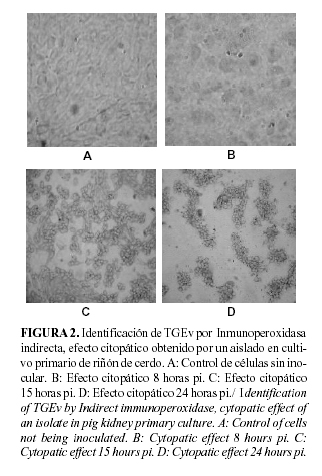
Siguiendo la metodología establecida en nuestro laboratorio, se analizó un grupo de muestras de diferentes provincias del país. Las muestras positivas (Tabla 1), mostraron el efecto citopatogénico caracterizado previamente. El mismo fue identificado como TGEv mediante la reacción positiva coloreada por IPI con el monoclonal comercial (1DB12) y el policlonal de cerdo (Figura 2).
La metodología en resumen que se estableció luego de este trabajo fue la siguiente:
Muestras:
• Raspado de mucosa + contenido intestinal o diarrea de cerditos de 24 a 48 horas de comenzados los síntomas, homogenizadas al 10% en medio DMEM con antibiótico 3 X. Filtrar a través de una membrana de acetato de celulosa de 0.45 µm (Centrisart, Sartoius) y tratar con suero fetal bovino inactivado.
Inoculación:
- Cultivos primarios de riñón de cerdo recién nacido en monocapas establecidas de 24 a 48 horas en placas de 24 pozos a 37ºC; atmósfera de CO2 5%.
- Retirar el medio de cultivo y lavar con medio sin suero.
- Diluir la muestra 1: 1,5 con medio de mantenimiento conteniendo tripsina 10 µg/mL.
- Volumen del inóculo/pozo: 0,5 mL.
- 1 hora de incubación a 37ºC; CO2 5%
- Retirar el inóculo.
- Lavar 1 vez con medio sin tripsina.
- Agregar 1 mL de medio sin tripsina.
- Observación diaria del ECP.
- Recoger el sobrenadante de los pocillos con ECP y congelar y descongelar a -80oC.
La aplicación de estos resultados permitirá implantar el servicio de diagnóstico de este virus en el grupo de Virología Animal del CENSA. Esto nos brinda la posibilidad de aumentar la eficiencia de nuestro diagnóstico, en el esclarecimiento de la etiología de un síndrome diarreico en porcinos, donde varios agentes infecciosos o no, pueden estar involucrados. Como todo diagnóstico rápido y oportuno, permite que se puedan tomar las medidas contra epizoóticas inmediatas, para el control de la enfermedad y minimizar las pérdidas debido a su diseminación a animales susceptibles. Aspecto que incidirá indirectamente sobre la reducción del número de focos de esta enfermedad a nivel nacional.
REFERENCIAS
1. Chen R, Huang W, Lin Z, Zhou Z, Yu H, Zhu D. Development of a novel real-time RT-PCR assay with LUX primer for the detection of swine transmissible gastroenteritis virus. J. Virol. Met.2004;122: 57-61.
2. ICTV : International Committee on Taxonomy of Viruses. [On line]. Disponible en: http://www.ncbi.nlm.nih.gov/ICTV.2004; Consulta 26 de julio de 2006.
3. OIE : Manual of Standard Diagnostic Tests and Vaccines, Part 2 Section 2.6 chapter 2.6.4.[online] 2004 Updated: 23.07.2006 http://www.oie.int/eng/normes/mmanual/A_00085.htm
4. OIE : Animal Diseases Data. Diseases Notifiable to the OIE. [On line].2005; Disponible en: http://www.oie.int/eng/maladies/en_classification.htm
5. Paton D. Lowings, P. Discrimination between transmissible gastroenteritis virus isolates. Arch Virol. 1997;142(8)1703-1711.
6. Pensaert M, Cox E, Van Deun K, Callebaut P. A seroepidemiological study of porcine respiratory coronavirus in Belgian swine. Vet Q. 1993;15: 16-20.
7. Rodríguez E, M Barrera, Betancourt , A. Gastroenteritis Transmisible del Cerdo: Un reto de la Industria Porcina. Rev. Elect. Vet. REDVET. 2005;Vol. VI. Nº 7. disponible en http://vet.org./revistas/redvet
8. Sestak K, Saif LJ. Porcine coronavirus. In Trends in emerging viral infection of swine. Iowa State Press, Ames. 2002;Pag 321-330 [consulted 15 February 2006].
9. US Department of Agricultura (USDA) : Swine 95 study. Part III: 1990-1995 changes in the US pork industry. 1997;555 South Howes, Fort Collins, CO 80521.
10.Jerez JA, Gregori F, Brandão PE, Rosales CAR, Ito HI, Buzinaro M, Sakai T. Isolation of bovine coronavirus (BCoV) in monolayers of HmLu-1 cells. Braz J Microbiol. 2005;36: 3.
(Recibido 5-9-2007; Aceptado 8-2-2008)


{kind=link}
{kind=link}
{kind=link}