










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Salud Animal
versión impresa ISSN 0253-570X
Rev Salud Anim. v.31 n.2 La Habana Mayo-ago. 2009
Artículo original
APLICACIÓN DE TÉCNICAS MOLECULARES EN LA CARACTERIZACIÓN DE AISLADOS DE Escherichia coli PROCEDENTES DE CERDOS CON SÍNDROME DIARREICO EN LA PROVINCIA DE VILLA CLARA
APPLICATION OF MOLECULAR TECHNIQUES IN THE CHARACTERIZATION OF Escherichia coli ISOLATES FROM PIGS WITH DIARRHOEIC SYNDROME IN VILLA CLARA PROVINCE
L. Lazo Pérez*, Ghizlane Dahbi**, M. Blanco Álvarez**, J.E. Blanco Álvarez**, J. Blanco Álvarez**, F. LLorens Blanco*
*Departamento de Medicina Veterinaria y Zootecnia. Facultad de Ciencias Agropecuarias. Universidad Central «Marta Abreu» de Las Villas. 54830 Santa Clara. Villa Clara, Cuba. Fax:(53)-(42)- 281778 (53)-(42)- 281618 Correo electrónico: lazo@agronet.uclv.edu.cu; ** Laboratorio de Referencia de E. coli (LREC). Departamento de Microbiología y Parasitología. Facultad de Veterinaria, 27002 LUGO, España TEL-FAX 982-285936 Correo electrónico: jba@lugo.usc.es
RESUMEN
Escherichia coli es una importante causa de diarrea en lechones recién nacidos y postdestetados, y ocasiona importantes pérdidas económicas en las explotaciones porcinas en todo el mundo. En el presente trabajo se realizó una caracterización de los aislados de Escherichia coli, a partir de muestras provenientes de cerdos con síndrome diarreico en la provincia de Villa Clara, para lo cual se aplicaron diferentes métodos de diagnóstico como la detección de los genes que codifican factores de virulencia, el serotipado y el estudio de los perfiles de electroforesis en campos pulsantes (PFGE). Los resultados mostraron, que en los procesos diarreicos causados por Escherichia coli, que afectan al cerdo durante la etapa neonatal y postdestete en diferentes granjas porcinas de la provincia de Villa Clara, predominan los patotipos O141 H- TSa TSb VT2 y O157 H19 VT2. La relativa baja incidencia de E. coli enterotoxigénico (ECET) y E. coli verotoxigénico (ECVT) en las muestras analizadas, permite inferir, que pudieran estar actuando otros agentes etiológicos en concomitancia con E. coli en los animales afectados. Los aislados de E. coli, a partir de muestras provenientes de cerdos con diarrea en la población estudiada, muestran un restringido número de serogrupos y serotipos, la mayoría de los seropatotipos evaluados presentaron gran diversidad genética, al mostrar diferentes clones con patrones electroforéticos distintos, se evidenció relación clonal entre los aislados de uno de los serotipos más prevalentes detectados en granjas ubicadas en zonas geográficamente distantes.
Palabras clave: diarrea; Escherichia coli; seropatotipos; perfiles de electroforesis en campos pulsantes (PFGE)
ABSTRCT
Escherichia coli is an important cause of diarrhoea in newborn and post weaning pigs, and it is responsible for significant economical losses at large-scale farms worldwide. In the present work, a characterization of Escherichia coli isolates was carried out in samples from pigs with diarrhoea syndrome in Villa Clara province. To accomplish this characterization, different diagnostic methods such as detection of genes encoding virulence factors, serotyped and study on pulsing-field gel electrophoresis (PFGE) were applied. Results showed that in the diarrhoeic processes caused by Escherichia coli affecting pigs during post-birth and post-weaning stages in different swine farms of Villa Clara province, the patotypes O141 H- STaSTb VT2 and O157 H19 VT2 were predominant. The relatively low incidence of enterotoxigenic E. coli (ETEC) and verotoxigenic E. coli (VTEC) in the analyzed samples allowed to infer that other etiological agents could be acting together with the group of E. coli isolate from the affected animals. Escherichia coli isolates from samples of pigs with diarrhoea in the population studied showed a restricted number of serogroups and serotypes and the majority of the seropatotypes evaluated presented a great genetic diversity due to the different clones with distinct electrophoresis patterns showed clone relationship among the isolates of one of the most prevailing detected serotypes in the farms located was evidenced in different geographical zones
Key words: diarrhoea; Escherichia coli; seropatotypes; pulsing-field gel electrophoresis (PFGE)
INTRODUCCIÓN
En Cuba, la colibacilosis es una de las primeras causas de mortalidad infecciosa en cerdos neonatos y jóvenes, con una pérdida anual de hasta un 10%, lo cual afecta la economía nacional en más de un millón de pesos. Muchas de estas pérdidas son provocadas por la debilidad y el incremento de su predisposición a otras enfermedades, además del costo en fármacos y el personal relacionado con el manejo del animal ((1.) Escherichia coli es la mayor causa de infección entérica en cerdos neonatos en Cuba (2). La diarrea postdestete en cerdos, es con frecuencia, el principal problema infeccioso de las granjas a gran escala, y es responsable de pérdidas significativas a nivel mundial (3).
Actualmente, se requiere de un diagnóstico preciso para poder enfrentar los planes de prevención y control de enfermedades emergentes y reemergentes que están incidiendo en la población porcina mundial en los últimos años. Ante esta amenaza, el diagnóstico debe encaminarse a la detección e identificación correcta de los agentes patógenos que prevalecen en las granjas porcinas, por lo que existe una tendencia mundial, a introducir métodos moleculares en el diagnóstico de las enfermedades transmisibles, ya que presentan mayor sensibilidad y se realizan en un menor tiempo que los métodos tradicionales. Las técnicas de biología molecular se han convertido desde mediados de los años ochenta, en herramientas necesarias en los laboratorios de diagnóstico de medicina veterinaria y medicina humana. Estas se basan en el análisis del genoma bacteriano en los tejidos del animal infectado que haya muerto, o en las secreciones de animales vivos enfermos (4).
El diagnóstico de la infección por Escherichia coli enterotoxigénica (ECET) se realiza a partir de la detección de factores de virulencia conocidos, del serogrupo de ECET sospechoso, y la determinación de la resistencia a antibióticos. Actualmente, la técnica más frecuentemente empleada para detectar E. coli diarreagénica es la PCR múltiple (Reacción en Cadena de la Polimerasa), capaz de detectar los genes de virulencia más representativos de ECEP, ECET, ECEI, ECVT, ECEA y ECAD (5). Para los estudios epidemiológicos es muy importante determinar el serotipo O: H de la cepa aislada (6).
La caracterización y clasificación de cepas patógenas de E. coli se puede desarrollar con métodos de biología molecular (7) Algunas de las herramientas de diagnóstico más recientes dentro de estos métodos de tipificación, análisis y caracterización epidemiológica son el polimorfismo de la longitud de los fragmentos de restricción (Restriction Fragment Length Polymorphism (RFLP)) y la electroforesis en campos pulsantes (Pulsed-Field Gel Electroforesis (PFGE), las cuales permiten el análisis del grado de relación genética o variabilidad entre diferentes serotipos de ECET, así como, cepas de algunos serogrupos (8,9,10,11,12,13).
Hay pocos estudios que divulgan las relaciones genéticas de cepas de E. coli aisladas en cerdos con diarrea (14, 4, 15, 16,8). La electroforesis en campos pulsantes ha dado mejores resultados para detectar las conexiones clonales entre cepas relacionadas epidemiológicamente. Esta metodología emplea enzimas de restricción que cortan de manera infrecuente el ADN cromosómico para generar fragmentos muy grandes que son separados por electroforesis. Los fragmentos de ADN generados son mayores de 40 kb y se pueden separar si se alterna cíclicamente la orientación del campo eléctrico durante la electroforesis1. Nos propusimos como objetivo en el presente trabajo, analizar las características principales de los aislados de E. coli procedentes de cerdos con síndrome diarreico en la provincia de Villa Clara.
MATERIALES Y MÉTODOS
Aislamiento e identificación de las cepas1: Las muestras se extrajeron de diez granjas porcinas localizadas en la provincia de Villa Clara, para lo cual se realizó hisopaje rectal a 399 cerdos con diarrea y con características clínicas compatibles con colibacilosis entérica, considerándose como criterio de exclusión aquellos animales que manifestaron diarreas de tipo sanguinolentas o con presencia de membranas difteroides.
Los cerdos muetreados estaban en edades comprendidas entre 0 y 33 días (crías), y entre 33 y 48 días (precebas). El muestreo fue realizado en los meses de enero a diciembre de 2002. Las muestras fueron sembradas en Agar MacConkey Lactosa (ML Oxoid, UK) y posteriormente transferidas a partir del crecimiento confluente a un medio de conservación compuesto por Agar nutritivo (Difco, EE.UU) 11.5 g, Caldo nutritivo (Difco, EE.UU) 4.0 g y Agua destilada 1 L, para su traslado al Laboratorio de Referencia de Escherichia coli (LREC) del Departamento de Microbiología de la Facultad de Medicina Veterinaria de la Universidad de Santiago de Compostela (USC), en España. A partir del medio de conservación se resembró en medio ML para el aislamiento e identificación, realizándose pruebas bioquímicas (TSA, Simón Citrato, Urea e Indol) a una colonia típica para su confirmación.
Reacción en Cadena de la Polimerasa (PCR) para la detección de los genes de virulencia: Se determinó la presencia de genes que codifican los factores de virulencia VT1, VT2, TL, TSa, TSb, eae, y además, los factores de adhesión K88, K99, P987, F17, F18 y F41en las cepas aisladas, mediante la técnica de Reacción en Cadena de la Polimerasa (PCR), para lo cual se aplicaron los procedimientos descritos por Blanco et al. 2004 (18).
Extracción del ADN a partir del crecimiento confluente: Las muestras fueron sembradas en medio MacConkey lactosa (ML) e incubadas (37°C/18 h). Se tomó con el asa, a partir del crecimiento confluente y se suspendió en 200 mL de agua bidestilada estéril contenidos en un tubo Eppendorf. La suspensión fue calentada a 100°C/5 minutos, y después centrifugada (10.000 rpm/5 min). Del sobrenadante se tomaron 7 mL para la amplificación. En los casos que dieron positivo, se realizó similar procedimiento a partir del crecimiento de l0 colonias aisladas, agrupadas en un pool de cinco colonias individuales.
Extracción del ADN a partir de colonias aisladas: Fueron sembradas l0 colonias en medio Agar Soya Triptona (TSA) e incubadas (37°C/18 h). Se tomaron cinco colonias (o su equivalente del crecimiento en placa) y fueron suspendidas en 150 ml de agua bidestilada estéril contenidos en un tubo Eppendorf. La suspensión fue calentada a 100°C/5 minutos, y después centrifugada (10.000 rpm/5 min). Del sobrenadante se tomaron 7 ml para la amplificación (en caso de dar positivas a factores de virulencia, se realiza similar procedimiento a partir del crecimiento de las colonias individuales).
Desarrollo de la PCR: La mezcla de reacción se preparó para un volumen final de 30 mL. que contenía 7 mL del ADN extraído de la muestra problema, los cuatro nucleótidos dATP, dGTP, dCTP y dTTP a una concentración final de 0,2 mM de cada uno de ellos, y Tris HCl 10 mM (pH 8.8), MgCl2 1,5 mM, KCl 50 mM y 1 U de taqpolimerasa (BIOTAQTM DNA polymerase, BIOLINE, Reino Unido). Los cebadores se utilizaron en una concentración de 30 a 300 ng / 100 ml dependiendo del gen a detectar (eae, TSa, TL, y K99 30 ng, y VT1, VT2, TSb, K88, P987, F17, F18 y F41, 300 ng).
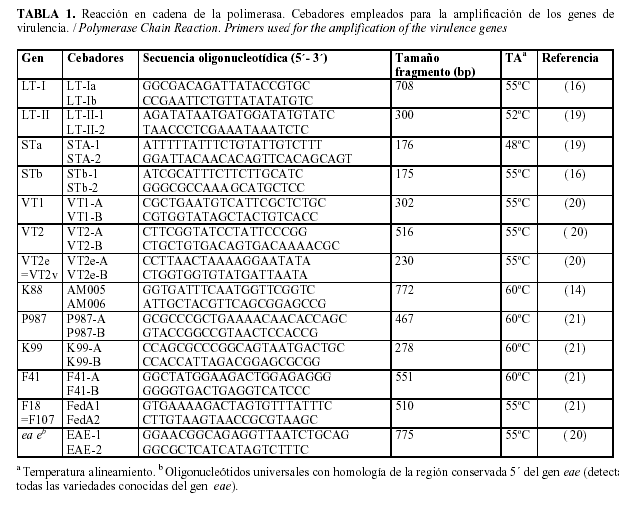
Cebadores: En la Tabla 1 se listan los cebadores utilizados en las pruebas de PCR, sus secuencias nucleotídicas, y el tamaño en pares de bases de los fragmentos amplificados. La mayoría de los pares de cebadores fueron diseñados en el LREC por el Dr. Miguel Blanco.
La amplificación se realizó en un termociclador (modelo PCR EXPRESS, HYBAID). Las mezclas de reacción fueron sometidas a 1 ciclo de desnaturalización inicial (94°C/2 min), seguido de 35 ciclos de 94°C/1 min (desnaturalización), 48ºC a 60ºC dependiendo del gen a detectar (48°C para TSa, 55°C para VT1, VT2, TL, TSb, eae y F18, y 60°C para K88, K99, P987 F17 y F41) / 1 min (para el acoplamiento) y 72ºC / 2 min (extensión). Se realizaron solamente dos amplificaciones múltiples (TL TSb y VT1 VT2).
Los productos amplificados se visualizaron mediante electroforesis en gel de agarosa al 2% (Seakem LE agarose, BMA, EE.UU.) en TBE (89 mM Tris, 89mM ácido bórico, 2.5 mM EDTA), previamente a la electroforesis, a la muestra a cargar en el gel (12 ul) se le adicionó 1 / 5 del volumen de tampón de muestra. Como marcador de pesos moleculares se usó el ADN del bacteriófago øX174 cortado con HaeIII (fragmentos de 1353, 1078, 872, 603, 310, 281, 271, 234, 194, 118 y 72 pares de bases) (Promega). La electroforesis se realizó a 124 vol, 400 mA
(miliamperes), durante 34 minutos. Los geles de agarosa se tiñeron con bromuro de etidio (10 mg/mL) (Merck) y se observaron bajo luz ultravioleta (transiluminador modelo 2010 Macrovue, LKB).
Serotipado: La determinación del antígeno O y H de las cepas fue realizada por medio de una técnica de microaglutinación, siguiendo la metodología originalmente descripta por Guinée et al. 1981(22) y modificada por Blanco et al. 1992 (20). Se emplearon todos los antisueros disponibles O (O1 - O185) y H (H1 - H56) en el LREC. Todos los antisueros fueron obtenidos y absorbidos con los antígenos de reacción cruzada correspondientes para eliminar las aglutininas no específicas. Los antisueros O fueron producidos en el LREC y los antisueros H fueron obtenidos del Statens Serum Institut (Copenhagen, Denmark).
Estudio de los perfiles de electroforesis en campos pulsantes (PFGE) en los seropatotipos de mayor prevalencia en las diferentes unidades del territorio: La electroforesis en campos pulsantes (PFGE) se utilizó con un sistema CHEF MAPPER (Bio-Rad, Hemel Hempstead, United Kingdom) mediante el protocolo standard descrito por la Enternet para PFGE (http://www.foodborne-net.de/content/e25/e70/e580/index_ger.html).
Para la preparación de las células bacterianas. Se suspendieron las bacterias en buffer CSB (Tris 100mM, EDTA 100mM) pH8 con proteinasa K (Roche) 0,5 mg/m. Para el ajuste de la densidad celular se leyó la densidad bacteriana en el espectrofotómetro a 420 nm y se ajustó a una concentración final de 0,8. Se usó CSB pH8 con proteinasa K para diluir.
Los geles fueron preparados con Seakem gold agarose (FMC) al 1% en buffer TE (Tris 10mM, EDTA 1mM) pH 8. Se atemperó en baño a 50-56ºC. Se mezcló en la proporción 1:1 la agarosa con la suspensión bacteriana, se repartió en los moldes y se dejó solidificar el gel a 4ºC. Para la lisis de las células en los moldes de agarosa se colocaron los moldes ya solidificados en 2 mL de buffer de lisis (Tris 50mM, EDTA 50mM, sarkosyl 1%) pH8 con 0,1 mg/mL de proteinasa K, añadida justo antes de utilizarlo. Se incubó 2 h mínimo en un baño con agitación a 54ºC, o durante 4 h sin agitación. Se lavaron los moldes dos veces a 50ºC en baño con agitación en 5 mL mínimo de agua destilada estéril. Se lavaron a continuación los moldes cinco veces a 50ºC en baño con agitación en 5 mL mínimo de buffer TE pH8. Se guardó a 4ºC en buffer TE pH 8 fresco hasta el día siguiente. Para la digestión de los moldes de agarosa se cortó un tercio del molde de agarosa, se añadió entre 0,2-0,8 U/µL del enzima de restricción XbaI (Roche) y se incubó a 37ºC durante 4 h como mínimo o toda la noche. Los otros dos tercios del molde se guardaron en TE fresco a 4ºC hasta un año. Se preparó un gel de Seakem gold agarose al 1,2% (FMC) en buffer TBE 0,5X (Tris 44 mM, ácido bórico 44 mM, EDTA 1 mM). Se colocaron los moldes en los pocillos y se selló con agarosa al 1% en buffer TBE 0,5X. En las posiciones 1, 8, 14 y 20 se colocó la cepa control (Salmonella branderup).
Para la electroforesis se empleó el buffer TBE 0,5X. Esta se ajustó a 14ºC. y se fijaron las condiciones de electroforesis (Pulso inicial: 15 s, Pulso final: 50 s, Rampa: lineal, Tª: 14ºC, Tiempo: 22h, Voltaje: 6v/cm). Los geles se tiñeron en buffer TBE O, 5X (con bromuro de etidio (10microgramos/ml) durante media hora aproximadamente. A continuación se lavaron durante media hora en agua destilada y se fotografiaron. Para comparar los pulsotipos PFGE, se analizaron los ficheros TIFF con el empleo de un software BioNumerics (Applied Maths, Sint-Martns-Latem, belgium). La relación clonal entre las cepas se determinó calculando el porcentaje de similitud estimado con el coeficiente de Dice, considerándose que las cepas estaban relacionadas genéticamente si el mismo era menor o igual que uno, o sea, el criterio de discriminación entre los clones fue considerado si existían diferencias de al menos un fragmento de restricción en los patrones electroforéticos. El análisis de los racimos del índice de similitud de Dice se basó en el método de estimación del peso de los pares de grupo usando la media aritmético (UPGMA) y fue hecho para generar un dendograma describiendo la relación entre los pulsotipos.
RESULTADOS Y DISCUSIÓN
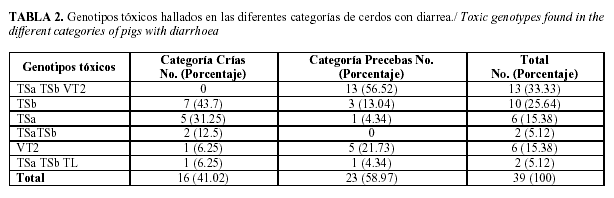
De los 399 cerdos con diarrea (209 lechones lactantes y 190 destetados) que fueron analizados en las diez granjas porcinas, solo 39 (9.77%) resultaron positivos a aislados de E. coli portadores de genes que codifican factores de virulencia, de los cuales 16 (41.02% del total de animales positivos) eran lechones lactantes y 23 (58.97%) lechones postdestetados. Como puede apreciarse en la Tabla 2, las cepas toxigénicas predominantes fueron las productoras de las enterotoxinas TSaTSbVT2 (33.33%), TSb (25.64%), TSa (15.38%) del total de las cepas positivas, TSa TSb (5.12%), y VT2 (15.38%) y TSaTSbTL (5.12%). En las muestras positivas de los lechones lactantes la enterotoxina TSb se halló en un 43.7%, TSa en un 31.25%, TSaTSb en un 12.5 %, VT2 y TSaTSbTL 6.25% respectivamente. En las muestras positivas de los lechones postdestetados, las enterotoxinas TSaTSbVT2 (56.52%), VT2 (21.73%), TSb (13.04%), TSa y TSaTSbTL (4.34%) respectivamente.
La relativa baja incidencia de E. coli con valor diagnóstico en las muestras analizadas a partir de cerdos con diarrea, nos permiten inferir que en los procesos diarreicos que padecen los cerdos de las categorías susceptibles pudieran reflejarse en otros agentes etiológicos como virus y protozoos, que pudieran tener una función importante como agentes primarios de estos procesos, aspecto que ha sido señalado por autores (23), además de el conjunto de factores predisponentes relacionados con el manejo y la alimentación que pueden favorecer la presentación de procesos diarreicos, elementos que son muy comunes en las condiciones de producción en Cuba (24).
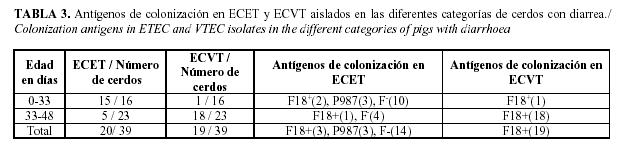
Como puede apreciarse en la (Tabla 3), los fenotipos tóxicos de ECET fueron hallados con mayor frecuencia en los lechones con edades comprendidas entre 0 y 33 días, mientras que ECVT se hallaron en mayor cuantía en los cerdos de 33 a 48 días de edad, resultando la mayoría de estos F18+. El 51.2 % del total de las muestras positivas resultaron cepas de ECVT y estas fueron aisladas en el 78.26% de cerdos destetados con edades comprendidas entre 33 y 48 días, mientras que el 48.71% del total de las muestras positivas resultaron ECET y fueron aisladas en el 87.5% de los cerditos lactantes (menores de 33 días), contra un 21.7% de las precebas.
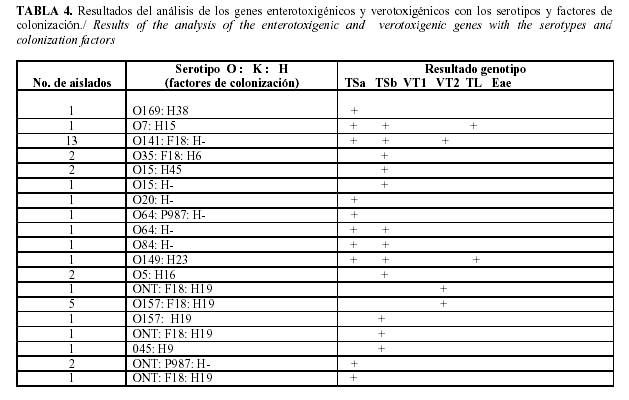
Se obtuvo un total de 39 aislados de E. coli que pertenecieron a los serotipos: O5:H16 (2 aislados), O7:H15 (1aislado), O15:H45 (2 aislados), O15:H- (1 aislado, O20:H- (1 aislado, O35:H6 (2 aislados), O45:H9 (1 aislado), O64:H- (2 aislados), O84:H- (1 aislado), O141:H- (13 aislados), O149:H23 (1 aislados, O157:H19 (6 aislados), O169:H38 (1 aislado), ONT:H19 (3 aislados), ONT: H- (2 aislados) (Tabla 4).
Los resultados obtenidos en este estudio indican que los serogrupos predominantes en el territorio son el O141 y el O157, los serotipos O141 H- y O157 H19, y los seropatotipos (combinación de serotipos y genes de virulencia) más frecuentes fueron el O141: H- TSa TSb VT2e F18 (13 aislados) y el O157:H19 VT2e F18 (5 aislados). Diferentes investigadores en diversas partes del mundo han hallado los serogrupos que nosotros hemos encontrado en este estudio. Otros autores (25), hallaron que los serotipos O8, O147, O149, O157 eran los más comunes en Norte América, todos productores de TL y TSb. Otros investigadores (26), encontraron que los grupos más comunes hallados en Australia son O9, O20 y O101. En estudios realizados en el Reino Unido (27), se demostró que los serotipos más comunes asociados a la diarrea en cerdos en esta región son O8, O138, O147, O149 y O157.
En la diarrea postdestete los principales grupos O más comúnmente involucrados en varias partes del mundo son O8, O138, O139, O141, O147, O149 y O157, de los cuales los que se reportan con mayor incidencia son O8, O141, O149 (28). Parte de estos resultados, coinciden con otros autores (15) quienes aislaron E. coli de los serogrupos O157 con las características F18 TSa TSb y O141 con las características F18 TSa TSb en cerdos con diarrea postdestete y enfermedad edemática, pero no portaban genes que codificaban para la verotoxina VT2 como fue hallado en este estudio.
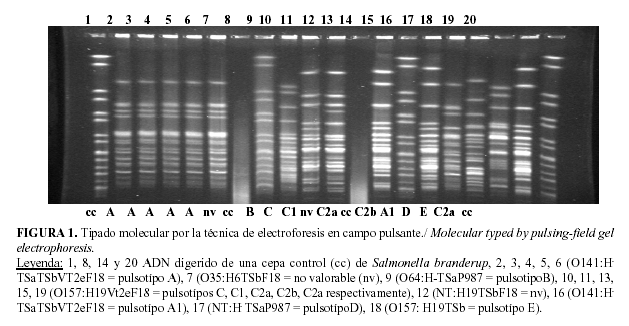
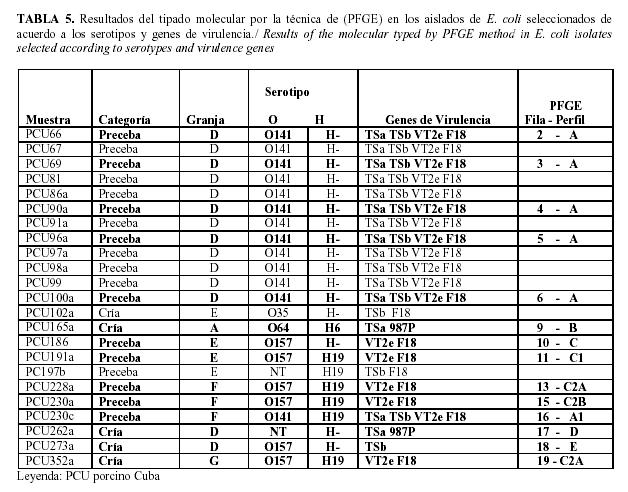
Al realizar la electroforesis en campos pulsantes, del ADN genómico de las cepas escogidas, digerido por la enzima de restricción XbaI (Figura 1) se puso de manifiesto, que todos los aislados del seropatotipo O141: H- TSa TSb VT2e F18 procedentes de cerdos afectados en la granja D, presentaron el mismo pulsotipo (A), por lo que pertenecen al mismo clon de virulencia.
El aislado PCU230c que presentó el mismo seropatotipo pero que fue aislado en otra granja (F), generó un pulsotipo (A1) muy similar al de los aislados de la granja D, lo que demuestra que están estrechamente relacionados. Los aislados del otro seropatotipo predomiante (O157: H19 VT2e F18) presentaron cinco perfiles de PFGE (o pulsotipos) cuatro de ellos muy similares o relacionados entre sí (C, C1, C2a, C2b) pero fueron hallados en granjas diferentes, C y C1 (en granja E), C2a (en granja F y G) y C2b (en granja F).
Por otra parte, el aislado PCU273a del seropatotipo O157: H19 TSb, hallado en la granja D mostró un perfil electroforético muy diferente (pulsotipo E) a los aislados O157: H19 VT2e F18 hallados en la granja E, F y G respectivamente, lo cual demuestra que no pertenecen a un mismo clon de virulencia y que a la vez son muy diferentes a los encontrados en los aislados del seropatotipo O141: H- TSa TSb VT2e F18 hallados en la granja D y F.
De acuerdo con los resultados obtenidos, en el estudio de la diversidad genética realizado, con 14 aislados de 16 seropatotipos más prevalentes, reveló 14 patrones de restricción distintos de PFGE enmarcados en 8 grupos (Tabla 5).
Los aislados del mismo seropatotipo fueron colocados juntos en el dendograma y fue observado un alto grado de polimorfismo entre aislados del serotipo O157:H19 (Figura 2).
Este aspecto también ha sido observado por otros estudios, con 79 aislados de E. coli de cerdos con PWD (serogrupos O8, O138, O141, O149, y O157) y 18 aislados del serotipo O149:K91:F4 (K88) en cerdos lactantes, donde fueron hallados 57 tipos electroforéticos (ETs), que mostraron una heterogeneidad genética considerable entre aislados de PWD del mismo serogrupo (29). Estos autores, encontraron además, una diversidad genética muy alta entre aislados de los serogrupos O8 y O138, mientras que la mayoría de los aislados de los serogrupos O141 y O149 tuvieron una relación más cercana.
En otro estudio, fueron analizados 43 aislados de E. coli de cerdos destetados que padecían enfermedad edemática o diarrea en Hungría. Los 43 aislados que pertenecían a los serogrupos O5, O21, O109, O138, O139, O147, O141, O157, demostraron 18 tipos electroforéticos distintos (8) y una homogeneidad genética fuerte que fue detectada entre O157 y los aislados O138; mientras que los otros serogrupos demostraron diversidad genética considerable, especialmente los aislados O139.
Por otra parte, fueron analizados 82 aislados de E. coli de cerdos con PWD, hallados en granjas separadas geográficamente en la parte occidental de Polonia. Los 82 aislados que pertenecían a cuatro serogrupos (O138, O139, O141, y O149) demostraron 13 patrones de PFGE diversos, y aunque fue observado un alto grado de polimorfismo, los que pertenecían al mismo serogrupo demostraron una relación cercana (10). Así, los 25 aislados del serotipo O149:K91 generaron solamente dos tipos de PFGE.
Al respecto, en Checoslovaquia, fue observado un total de 36 patrones de restricción PFGE diferentes en 46 ECET y ECVT aislados en cerdos postdestetados con diarrea. Los aislados del mismo seropatotipo, mostraron un alto grado de polimorfismo. Además, 13 patrones PFGE distintos resultaron de 14 O149:H10 aislados analizados (3). Similares resultados han sido encontrados en España en los aislados de los serotipos más frecuentes (O157: H- TL TSb F4/K88) (21).
El estudio de la diversidad genética en los aislados, permitió evaluar por primera vez en Cuba y en la provincia, la relación clonal entre E. coli patógenas aisladas en diferentes granjas, lo cual nos sirve de base para iniciar estudios de epidemiología molecular que nos permitan comparar los nuevos aislamientos en las diferentes granjas, el posible origen y evolución de los seropatotipos de mayor prevalencia en la provincia.
Resultan interesantes los hallazgos encontrados, relacionados con la presencia de diversos patrones electroforéticos o pulsotipos, en uno de los serotipos más frecuentes aislados en los cerdos con diarrea en
la provincia, lo cual demuestra que son necesarios otros estudios para conocer la prevalencia de algunos de estos clones, analizar su evolución y determinar su relación con la patogenicidad en las diferentes granjas donde aparezcan. El presente estudio constituye un punto de partida para poder evaluar la diversidad genética y el grado de relación clonal de los aislados de E. coli productores de diarrea en cerdos. El análisis de los clones nos permite determinar si los aislados son de reciente introducción o similares a los de mayor prevalencia que circulan en las granjas analizadas. También nos permite analizar la evolución o distribución espacial como aspecto importante a valorar en el poder de difusión de la enfermedad y el conjunto de factores que favorecen la transmisibilidad entre las granjas.
En este estudio se pone de manifiesto la relación clonal de algunos seropatotipos aislados en granjas ubicadas en diferentes regiones del territorio, aspecto que consideramos que pudiera deberse a la transportación de cerdos de diferentes zonas geográficas dentro del territorio, hacia un mismo matadero, en vehículos que no cumplen las normas de bioseguridad del transporte. Los resultados indican que la población de cerdos estudiada, como se describió en otros países ECET y ECVT aisladas de cerdos con diarrea, pertenecen a un restringido número de serogrupos y serotipos. Aparentemente dos seropatotipos son predominantes en la diarrea postdestete: O141: H-: TSa/TSb/VT2e/F18 y O157:H19:VT2e/F18. Sin embargo, antes de dar conclusiones definitivas sobre los más importantes seropatotipos implicados en la diarrea porcina en Villa Clara, es necesario incluir en estudios futuros un mayor número de muestras; no obstante, teniendo en cuenta que los aislados proceden de diez granjas representativas con alta prevalencia de diarrea en la provincia, los resultados obtenidos son confiables.
Estos resultados constituyen una importante fuente de información que sirve de base no solo para los estudios epidemiológicos, sino también, para la elaboración de vacunas autóctonas, a partir de un cepario, que pudiera conservar los aislados con las características deseadas, para un candidato vacunal a partir de cepas que circulan en el territorio, lo cual pudiera contribuir al éxito del conjunto de medidas contraepizoóticas preventivas y recuperativas que establece el programa de lucha y control de esta entidad, establecido por el IMV, el que considera como uno de sus objetivos la elaboración de vacunas autóctonas, para aquellos lugares que así lo requieran.
CONCLUSIONES
En los procesos diarreicos causados por Escherichia coli que afectan al cerdo durante la etapa neonatal y postdestete en diferentes granjas porcinas de la provincia de Villa Clara, aparentemente predominan los patotipos O141 H- TSa TSb VT2 y O157 H19 VT2.
La relativa baja incidencia de E. coli enterotoxigénico (ECET) y E. coli verotoxigénico (ECVT) en las muestras analizadas, permite inferir, que pudieran estar actuando otros agentes etiológicos en concomitancia con E. coli en los animales afectados.
Los aislados de E. coli a partir de muestras provenientes de cerdos con diarrea en la población estudiada, muestran un restringido número de serogrupos y serotipos y la mayoría de los seropatotipos evaluados presentaron gran diversidad genética, al mostrar diferentes clones con patrones electroforéticos distintos, lo que evidenció relación clonal entre los aislados en uno de los serotipos más prevalentes detectados en granjas ubicadas de zonas geográficamente distantes.
AGRADECIMIENTOS
Al Dr. C Jorge Blanco Álvarez, por su colaboración con los medios y equipamiento necesarios para la investigación en el Laboratorio de Referencia de E. coli (LREC) del Departamento de Microbiología de la Facultad de Veterinaria en la Universidad de Santiago de Compostela (USC).
Al Dr. C Edilbert Van Drissche del Laboratorio de Proteínas del Instituto de Biología Molecular de la Universidad libre de Bruselas, por facilitarme la participación en un proyecto de investigación internacional y facilitado el financiamiento de estancias en el extranjero para realizar estos experimentos.
REFERENCIAS
1. Wong I, Bover E, Ramos M, González N, Expósito M, Segura R et al. Eficacia en condiciones de campo de una vacuna recombinante contra la colibacilosis porcina. Biotecnologia Aplicada. 1996;13:16-19.
2. Castro MD, Campal A, Arteaga N, Miranda A, Junco J, León et al. Immunodetection of colonizing antigens in enterotoxigenic Escherichia coli cells isolated from piglets with diarrhoea in Cuba. 6to Congreso Latinoamericano de Inmunología y 3er Congreso Cubano de Inmunología. La Habana. Cuba. Diciembre 9-13 de 2002.
3. VuKhac H, Holada E, Pilipcinec E, Blanco M, Blanco JE, Mora A et al. Serotypes, virulence genes, and PFGE profiles of Escherichia coli isolated from pigs with postweaning diarrhoea in Slovakia. Vet Rec. 2006;20:2-10.
4. Mogollón GJD, Rincón MMA, Arbeláez G, Ruiz S, de La Rosa S, Lara AM. Aplicación de las técnicas moleculares para el diagnóstico de las enfermedades porcinas en Colombia. Informa. 2002;29(1):30-36.
5. Le Guennec, J. Diagnóstico de las diarreas neonatales II. [en línea]. [Fecha de consulta 29/3/05]. Disponible en: www.3tres3. com E. coli diarrea neonatal. Diagnóstico. Htm. 2005.
6. Blanco J, Blanco M, Blanco JE, Mora A, Alonso MP. et al. Enterobacterias: características generales. Género Escherichia. En: Manual de Microbiología Veterinaria. Vadillo S., Píriz S & Mateos E. Eds McGraw-Hill Interamericana, Madrid. 2002;301-325.
7. Rodríguez GA. Principales características y diagnóstico de los grupos patógenos de Escherichia coli. Revista electrónica. Salud pública de México. 44 (5). [en línea]. [Fecha de consulta 17/8/05]. Disponible en: http://www.scielosp.org/scielo.php/script_sci_serial /pid_0036-3634/lng_en/nrm_iso, 2002.
8. Osek J. Genetic diversity among Escherichia coli O141: K 91 strains isolated from pigs with diarrhoea determined by randomlys amplified polymorphic DNA analysis. Res Vet Sci. 1999;67:197-198.
9. Jacek O. Clonal analysis of Escherichia coli strains isolated from pigs with post-weaning diarrhoea by pulsed-field gel electrophoresis. FEMS Microbiology Letters. 2000;186:327-331.
10.Preston MA, Johnson W, Khakhria R, Borczyk A. Epidemiologic subtyping of Escherichia coli serogroup O157 strains isolated in Ontario by phage typing and pulsed-field gel electrophoresis. J Clin Microbiol. 2000; 38:2366-2368.
11.Guinée PA, Jansen, WH, Wadstrom T, Sellwood, R. Escherichia coli associated with neonatal diarrhoea in piglets and calves. Curr. Top. Vet Anima Sci. 1981;13:126-162.
12.Dobrint U, Agerer F, Michaelis K, Janka A, Buchrieser C, Samuelson M et al. Analysis of genome plasticity in pathogenic and commensal Escherichia coli isolates by use of DNA arrays. J Bacteriol. 2003;185:1831-1840.
13.Mora A, Blanco M, Blanco JE, Dhabi G, Alonso MP, Stirrat A et al. Epidemiologic subtyping of Escherichia coli O157:H7 strains isolated in Spain by phage typing and pulsed-field gel electrophoresis. 5th International Symposium on "Shiga Toxin (Verocytotoxin)-Producing Escherichia coli infections". Edinburgh. Final programme and Book of Abstracts, 2003 ;P195 abstract, p. 177.
14.Franklin MA, Francis DH, Baker D, Mathew AG. A PCR-based method of detection and differentiation of K88 adhesive Escherichia coli. J Vet Diagn Invest.1996; 8:460-463.
15.Nagy B, Wilson RA, Whittam TS. Genety diversity among Escherichia coli isolates carrying F18 genes from pigs with porcine postweaning diarrhea and edema disease. J Clin Microbiol 1999;37:1642-1645.
16.Blanco M, Blanco JE, González EA, Mora A, Jansen W, Gomes TAT et al. Genes coding for enterotoxins and verotoxins in porcine enteropathogenic Escherichia coli strains belonging to different O: K: H serotypes. Relationship with toxic phenotypes. J Clin Microbiol. 1997;35 (11):2958-2963.
17.Alexander TJL. Neonatal diarrhea in pigs. En: Gyles CL, ed. Escherichia coli in domestic animals and humans. Wallinglord, United Kingdom: CAB International 1994. Pp. 153-155.
18.Blanco M, Podola NL, Krüger A, Sanz ME, Blanco JE, González EA et al. Virulence genes and intimin types of Shiga-toxin-producing Escherichia coli isolated from cattle and beef products in Argentina. Int Microbio. 2004;7:269-276.
19.Penteado AS, Ugrinovich LA, Blanco J, Blanco M, Blanco JE, Mora A et al. Serobiotypes and virulence genes of Escherichia coli strains isolated from diarrheic and healthy rabbits in Brazil. Vet Microbiol. 2002;89:41-51.
20.Hampson DJ, Woodwar HM. Connaughton ID: Genetic analysis of porcine postweaning diarrhea. Epidemiol Infect. 1993;110:575-581.
21.Blanco AM, Lazo PL, Blanco AJE, Dhabi G, Mora A, López C et al. Serotypes, virulence genes and PFGE patterns of enteropathogenic Escherichia coli isolated from Cuban pigs with diarrhoea. International Microbiology 2006;9(1):53-60.
22.Eklund M, Scheutz F, Siitonen A. Clinical isolates of non-O157 Shiga toxin-producing Escherichia coli: serotypes, virulence characteristics, and molecular profiles of strains of the same serotype. J Clin Microbiol. 2001; 39:2829-2834.
23.Nagy B, Nagy G, Meder M, Mocsari E. Enterotoxigenic E. coli, rotavirus, porcine epidemic diarrhea virus, adhenovirus and calici-like virus in porcine postweaning diarrhoea in Hungary. Acta Vet Hung. 1996;44:9-19.
24.Lazo PL, Hernández LL, Llorens BF, Molina SR, García FE, Cruz ME et al. Impacto sanitario de la colibacilosis entérica porcina en la provincia de Villa Clara: evaluación de sustancias bloqueadoras de la colonización intestinal. Memorias del XVIII Congreso Panamericano de Ciencias Veterinarias. La Habana. Noviembre de 2002.
25.Harel J, Lapointe H, Fallara A, Lortie L, Poulin Mb et al . Detection of genes for fimbrial antigens and enterotoxins associated with E. coli serogroups isolated from pigs with diarrhea. J Clin Microbiol. 1991;29:745-752.
26.Woodward JM, Connaughton ID, Fahy VA. Clonal analysis of Escherichia coli of serogroups O9, O20, and O101 isolated from Australian pigs with neonatal diarrhoea. J Clin Microbiol. 1993;31:1185-118.
(Recibido 14-2-2007; Aceptado 5-1-2009)
1 Blanco, J. (2008): Comunicación personal. Laboratorio de Referencia de Escherichia coli. Facultad de Medicina Veterinaria. Universidad de Santiago de Compostela. España.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}