










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Salud Animal
versión impresa ISSN 0253-570X
Rev Salud Anim. v.33 n.1 La Habana ene.-abr. 2011
Artículo original
ANÁLISIS DE CEPAS DE Anaplasma marginale UTILIZANDO LAS SECUENCIAS REPETIDAS DE LA PROTEÍNA PRINCIPAL DE SUPERFICIE LA (MSP1a)
ANALYSIS OF Anaplasma marginale STRAINS USING THE REPEATED SEQUENCES OF MAJOR SURFACE PROTEIN 1a (MSP1a)
Belkis Corona, Ariadna Rodríguez, Siomara Martínez
Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, La Habana, Cuba. Correo electrónico: bcorona@censa.edu.cu
RESUMEN
Anaplasma marginale es un patógeno del ganado bovino, que causa severas pérdidas económicas en las regiones tropicales y subtropicales. Las proteínas principales de superficie de los cuerpos iniciales de este hemoparásito están involucradas en las interacciones patógeno-hospedero y garrapata-patógeno y han sido utilizadas como marcadores para la caracterización genética de cepas de Anaplasma marginale y estudios filogenéticos. La proteína MSP1a varía entre cepas de regiones geográficas diferentes en el número y secuencias de las regiones repetidas en tandem en la región amino-terminal. En el presente estudio caracterizamos 24 cepas de Anaplasma marginale utilizando la secuencia descrita para la proteína MSP1a. Estos resultados corroboran la heterogeneidad de cepas de Anaplasma marginale en regiones endémicas y que el análisis filogenético de las secuencias repetidas en tandem de esta proteína MSP1a no resulta en clusters relacionados con el origen geográfico de las cepas, pero contribuye a entender la diversidad genética y la evolución de A. marginale.
Palabras clave: Anaplasma marginale; proteínas principales de superficie; MSP1a.
ABSTRACT
Anaplasma marginale is a pathogen for cattle, causing severe economic losses in tropical and subtropical regions. The major surface proteins of initial bodies are involved in pathogen-host and tick-pathogen interactions and have been used as markers for the genetic characterization of Anaplasma marginale strains and for phylogenetic studies. MSP1a protein varies between strains of different geographical regions in the number and sequence of tandem repeated sequences in the amino-terminal region. In this study, 24 Anaplasma marginale strains were characterized using the sequence described for MSP1a protein. These results support the heterogeneity of Anaplasma marginale strains in endemic regions and that the phylogenetic analysis of tandem repeated sequences of this MSP1a protein does not result in clusters related to the geographical origin of strains, but helps to understand the genetic diversity and evolution of Anaplasma marginale.
Key words: Anaplasma marginale; major surface protein; MSP1a.
INTRODUCCIÓN
Anaplasma marginale (Rickettsiales: Anaplasma-taceae) es un patógeno que causa importantes pérdidas económicas en el ganado bovino de regiones tropicales y subtropicales (1).
Cepas de A. marginale de diferentes regiones geográficas han sido identificadas y difieren en su biología, características genéticas y trasmisibilidad por garrapatas (2). La diversidad genética de las cepas de A. marginale ha sido caracterizada utilizando los genes que codifican para las proteínas principales de superficie que están involucradas con células de los hospederos vertebrados e invertebrados (2).
La fracción de membrana externa de A. marginale está compuesta por al menos seis polipéptidos principales de superficie, los cuales incluyen las proteínas principales (MSPs), MSP1a, MSP1b, MSP2, MSP3, MSP4 y MSP5 (3,4,5).
Se conoce que la proteína principal de superficie 1a (MSP1a), codificada por el gen msp1a, está involucrada en las interacciones patógeno-hospedero y garrapata-patógeno (2). Esta proteína es una adhesina para el eritrocito bovino y la célula de garrapata (6, 7) y el dominio de adhesión está contenido en la región variable N-terminal (8). MSP1a ha mostrado estar involucrada en la transmisión de A. marginale por la garrapata Dermacentor spp. (9) y contiene epítopes de células B en los repetidos de la región N-terminal que pueden estar involucrados en la respuesta protectiva contra la infección de A. marginale en bovinos (10).
La proteína MSP1a de A. marginale ha estado bajo selección por presión positiva y en cepas de diferentes regiones geográficas, difiere en el peso molecular debido a la presencia de un número variable de repeticiones en tandem (11,9,2,12).
A pesar del alto grado de variación en la secuencia dentro de áreas endémicas, el análisis de la secuencia de MSP1a de cepas de Norte y Sur América, Italia, Israel y Australia no proporcionaron información filogeográfica (2). Sin embargo el análisis de las secuencias repetidas a una escala global puede proporcionar información filogenética y evolutiva de cepas de Anaplasma marginale (13).
El presente estudio tiene como objetivo realizar una caracterización de cepas de diferentes regiones geográficas utilizando las secuencias de la proteína msp1a de cepas de A. marginale aisladas de bovino.
MATERIALES Y MÉTODOS
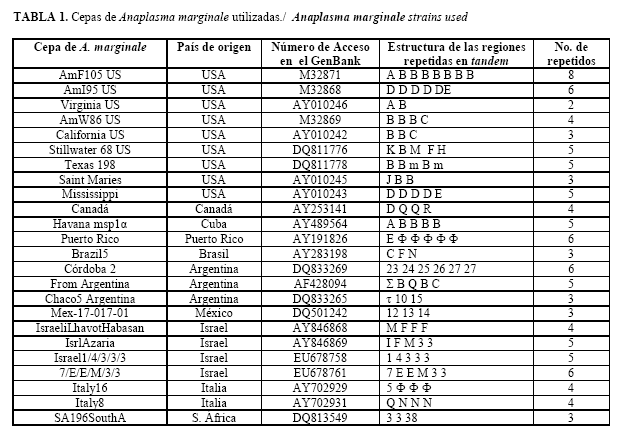
Secuenciación del gen msp1a del aislamiento Habana de A. marginale: La secuencia del gen msp1a del aislamiento Habana de A. marginale clonado en pmal (datos no publicados), se realizó por la compañía francesa Genoma Express, utilizando los siguientes cebadores (Tabla 1):
5'GGAAGATCTTCCATGTCAGCAGAGTATG3' y 5'GGAAGATCTTCCTTACGCCGCCGCCTG3'. El análisis y alineamiento de la secuencia se realizó utilizando el programa DNAsis. Para confirmar la identidad de la secuencia, esta fue analizada en el Blastn y Blastx, respectivamente (14,15) (http://www.ncbi.nlm. nih.gov/). La secuencia fue depositada en el GenBank con número de acceso AY489564.
Cepas de A. marginale: Las cepas utilizadas en este estudio y el número de acceso en el Genbank están listados en la Tabla 1. Todas las cepas fueron obtenidas de bovinos naturalmente infectados.
Análisis de secuencias: El alineamiento múltiple de las secuencias fue realizado utilizando el programa ClustalW v1.83 (16), en la plataforma Web: http://www.genome.jp empleando los parámetros sugeridos. Tanto para los pairwise-aligments como para los alineamientos múltiples se utilizó la matriz de peso BLOSUM62. Para la confección del árbol filogenético se utilizó el método de NJ (Neirghbour Joining) descrito por Saitou y Nei (17). Para confeccionar el árbol se calcularon las distancias (porcentaje de divergencia) entre todos los pares de secuencias del alineamiento múltiple y posteriormente se aplicó el método NJ a la matriz de distancias.
RESULTADOS Y DISCUSIÓN
El gen msp1a de la cepa Habana de Anaplasma marginale es un amplicón de 2043 pb, cuando se utilizan los cebadores descritos para la amplificación de dicho gen. El mismo mostró cinco repetidos, de ellos un repetido del tipo A y cuatro repetidos del tipo B, según las secuencias de los repetidos, descritas por Palmer et al. (18).
En el presente estudio se analizaron 24 cepas de países de América del Norte, América del Sur, Europa, Asia y África (Fig. 2 y Fig. 3a, 3b, 3c), identificándose en estas cepas 34 secuencias repetidas diferentes en MSP1a (Fig. 1). El análisis de dichas secuencias corrobora la heterogeneidad genética de las cepas de A. marginale.

Las relaciones filogenéticas entre la cepa cubana y las cepas de Anaplasma marginale de otras regiones geográficas (América del Norte y América del Sur, Asia, África y Europa) fueron inferidas utilizando la secuencia de los repetidos de la proteína MSP1a, depositadas en el GenBank. En la Figura 3a, 3b y 3c, podemos observar los cambios en la composición de aminoácidos en las secuencias repetidas en tandem, lo que da lugar a la gran variedad de repetidos que existen.
El análisis de la secuencia de MSP1a en cepas de diferentes regiones geográficas no proporciona información filogeográfica debido al alto grado de variación en la secuencia de la proteína MSP1a dentro de un área endémica (2). Sin embargo, el análisis filogenético de toda la región variable de la proteína MSP1a, en las diferentes cepas podría proporcionar información filogenética sobre cepas de Anaplasma marginale.
Con el análisis realizado (Fig. 2), no se observó una segregación filogeográfica definitiva. La cepa Habana aparece muy cerca de las cepas Florida y Virginia de Estados Unidos, con las que comparte el mismo tipo de repetidos y con la cepa Argentina con la que solo comparte el repetido B; esto reafirma la hiPótesis planteada por de la Fuente et al. (19,20), de que por presión positiva hayan sufrido modificaciones que resultaron en un incremento o disminución del número de repetidos en el fragmento variable. Sin embargo está distante de otras cepas de América, las cuales a su vez, están separadas o cercanas a cepas de Europa y Asia.
Los resultados obtenidos en el presente estudio soportan la hipótesis planteada por de la Fuente et al. (13), que plantean que la heterogeneidad genética observada entre cepas de A. marginale dentro de un área endémica puede ser explicada por el movimiento de ganado y el mantenimiento de diferentes genotipos por los eventos de transmisión independiente, debido a la exclusión de la infección de A. marginale en bovinos y garrapatas, los cuales comúnmente resultan en el establecimiento de solo un genotipo por animal (2). Sin embargo, cuando genotipos distantemente relacionados existen en la misma región, es posible la infección de un simple hospedero con múltiples cepas de A. marginale (21). Otro elemento a considerar es la influencia que la garrapata Boophilus microplus, como vector trans-estadial de la rickettsia (22,23), pueda tener dentro de la generación de diversidad en esta rickettsia, ya que aunque se sabe que MSP1a es un ligando para células de algunas garrapatas, se desconoce la función de esta proteína en relación con su trasmisibilidad por garrapatas del género Boophilus.
La presencia de diferentes genotipos de MSP1a en diferentes países (20,24, 25,26) sugiere que las secuencias de MSP1a, aunque se conservan durante la multiplicación del parásito en el hospedero bovino y después de la transmisión por garrapatas (18, 27), están cambiando rápidamente, dando como resultado variaciones en el genotipo dentro de la población de Anaplasma marginale. Esta heterogenicidad entre las secuencias puede probablemente ser explicada como un resultado de la posición de las secuencias repetidas en tandem de A. marginale dentro de la región que muestra alta mutabilidad (27).
Nuestros resultados coinciden con los obtenidos por Vidotto et al. (28), que con su estudio demuestran que la secuencia del gen msp1á y de la proteína MSP1a no proporcionan una resolución filogeográfica distintiva por su alta variabilidad, lo que coincide además con los resultados obtenidos por de la Fuente et al. (20, 8, 24). En contraste se ha descrito que la secuencia del gen msp4 puede proporcionar información filogeográfica útil (24).
El resultado informado en este estudio proporciona información sobre la evolución de cepas de Anaplasma marginale en áreas endémicas, utilizando las secuencias repetidas de la proteína MSP1a y una vez más queda demostrada la heterogeneidad genética de las cepas de A. marginale.
REFERENCIAS
1. Kocan KM, de la Fuente J, Blouin EF, Garcia-Garcia JC. Anaplasma marginale (Rickettsiales: Anaplsmataceae): recent advances in defining host-pathogen adaptations of a tick borne rickettsia. Parasitology. 2004;129:285-300.
2. de la Fuente J, Rubial P, Mtshali MS, Naranjo V, Shuqing L, Mangold AJ, et al. Analysis of world strains of Anaplasma marginale using major surface protein 1a repeat sequence. Vet Microbiol. 2005;119:382-390.
3. Palmer GH, McGuire TC. Immune serum against Anaplasma marginale initial bodies neutralizes infectivity for cattle. Infect Immun. 1984;13:1010-1015.
4. Tebele N, McGuire TC. Induction of protective immunity using Anaplasma marginale initial body membranes. Infect Immun. 1991;59:3199-3204.
5. Visser ES, McGuire C, Palmer GH, Davis WC, Shkap V, Pipano E, et al. The Anaplasma marginale msp5 genes encoded 19 kDa protein conserved in all recognized Anaplasma species. Infect Immun. 1992;60:5139-5144.
6. McGarey DJ, Allred DR. Characterization of hemaglutinating components of the Anaplasma marginale initial body surface and identification of possible adhesins. Infect Immun. 1994;62:4587-4593.
7. de la Fuente J, Garcia-Garcia JC, Blouin EF, Kocan KM. Major surface protein 1a effects tick infection and transmission of the ehrlichial pathogen Anaplasma marginale. Int J Parasitol. 2001;31:1705-1714.
8. de la Fuente J, Garcia-Garcia JC, Blouin EF, Kocan KM. Characterization of the functional domain of major surface protein 1a evolved in adhesion of the rickettsia Anaplasma marginale to host cells. Vet Microbiol. 2003;91:265-283.
9. de la Fuente J, Van Den Bussche RA, Kocan KM. Molecular phylogeny and biogeography of North American isolates of Anaplasma marginale (Rickettsiaceae: Ehrlichieae). Vet Parasitol. 2001;97:65-76.
10.Garcia-Garcia JC, de la Fuente J, Bell-Eunice J, Blouin EF, Kocan KM. Glycosylation of Anaplasma marginale major surface protein 1a and its putative role in adhesion to tick cells. Infect Imm. 2004;72(5):3022-3030.
11.Allred DR, McGuire TC, Palmer GH, Leib SR, Harkins TM, McElwain TF, et al. Molecular basis for surface antigen size polymorphisms and conservation of a neutralization-sensitive epitope in Anaplasma marginale. Proc Natl Acad Sci. USA. 1990;87:3220-3224.
12.Corona B, Minet C, Albina E, Vega A, Martínez S. Sequence of msp1á gene of Anaplasma marginale Havana isolate and expression in eukaryotic cells. Span J Agric Res. 2005;3(3):275-280.
13.de la Fuente J, Rubial P, Mtshali MS, Naranjo V, Shuqing L, Mangold AJ, et al. Analysis of world strains of Anaplasma marginale using major surface protein 1a repeat sequences. Vet Microbiol. 2007;119:382-390.
14. Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment searh tool. J Mol Biol. 1990;215:403-410.
15.Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W, et al. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 1997;25:3389-3402.
16.Jeanmougin F, Thompson JD, Gouy M, Higgins DG, Gibson TJ. Multiple sequence alignment with Clustal X. Trends Biochem Sci. 1998;23:403-405.
17.Saitou N, Nei M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987;4:406-425.
18.Palmer GH, Rurangirwa FR, McElwain TF. Strain composition of the ehrlichia Anaplasma marginale within persistently infected cattle, a mammalian reservoir for tick transmission. J Clin Microbiol. 2001;39:631-635.
19.de la Fuente J, Garcia-Garcia JC, Blouin EF, Saliki JT, Kocan KM. Infection of tick cells and bovine erythrocytes with one genotype of the intracellular ehrlichia Anaplasma marginale excludes infection with other genotypes. Clin Diagn Lab Immunol. 2001;9:658-668.
20.de la Fuente J, Garcia-Garcia JC, Blouin EF, Rodriguez SD, Garcia A, Kocan KM. Evolution and function of tandem repeats in the major surface protein 1a of the ehrlichial pathogen Anaplasma marginale. Anim Health Res Rev. 2002;2:163-173.
21.Palmer GH, Knowles DP Jr, Rodríguez JL, Gnad DP, Hollis LC, Marston T, et al. Stochastic transmission of multiple genotypically distinct Anaplasma marginale strains in a herd with high prevalence of Anaplasma infection. J Clin Microbiol. 2004;42:5381-5384.
22.Aguirre DH, Gaido AB, Vinabal AE, de Echaide ST, Guglielmone AA. Transmission of Anaplasma marginale with adult Boophilus microplus ticks fed as nymphs on calves with different levels of rickettsaemia. Parasite. 1994;1(4):405-407.
23.Scoles GA, Ueti MW, Noh SM, Knowles DP, Palmer GH. Conservation of transmission phenotype of Anaplasma marginale (Rickettsiales: Anaplasmataceae) strains among Dermacentor and Rhipicephalus ticks (Acari: Ixodidae). J Med Entomol. 2007;44(3):484-491.
24.de la Fuente J, Garcia-Garcia JC, Barbet AF, Blouin EF, Kocan KM. Adhesion of outer membrane proteins containing tandem repeats of Anaplasma and Ehrlichia species (Rickettsiales: Anaplasmataceae) to tick cells. Vet Microbiol. 2004;98:313-322.
25.Ferreira AM, Suzart S, Vidotto O, Knowles DP, Vidotto MC. Use of the repetitive DNA elements to define genetic relationships among Anaplasma marginale isolates. FEMS Microbiol Lett. 2001;197:139-143.
26.Lew AE, Bock RE, Minchin CM, Masaka S. A msp1 alpha polymerase chain reaction assay for specific detection and differentiation of Anaplasma marginale isolates. Vet Microbiol. 2002;86:325-335.
27.Bowie MV, de la Fuente J, Tocan KM, Blouin EF, Barbet AF. Conservation of major surface protein 1 genes of the ehrlichial pathogen Anaplasma marginale during cyclic transmission between ticks and cattle. Gene. 2002;282:95-102.
28.Vidotto MC, Kano FS, Gregori F, Headley SA, Vidotto O. Phylogenetic analysis of Anaplasma marginale strains from Parana´ state, Brazil, using the msp1a and msp4 genes. J Vet Med. Series B. 2006;53:404-411.
(Recibido 12-10-2009; Aceptado 15-7-2010)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}