










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Salud Animal
versión impresa ISSN 0253-570X
Rev Salud Anim. v.33 n.1 La Habana ene.-abr. 2011
Artículo original
DETECCCIÓN DE Anaplasma marginale EN BOVINOS, MEDIANTE LA AMPLIFICACIÓN POR PCR DEL GEN msp5
DETECTION OF Anaplasma marginale IN BOVINE, USING THE msp5 GENE AMPLIFICATION BY PCR
Belkis Corona, Siomara Martínez
Centro Nacional de Sanidad Agropecuaria. Apartado 10, San José de las Lajas, La Habana, Cuba. Correo electrónico: bcorona@censa.edu.cu
RESUMEN
Anaplasma marginale es una rickettsia del genogrupo II de las Ehrlichias, que parasita los eritrocitos maduros del ganado bovino. Hasta el momento no se cuenta con un método de control eficaz contra la enfermedad, por lo que resulta de gran importancia desarrollar un inmunógeno capaz de prevenir la infección con este patógeno. A esto se le agrega la necesidad de contar con técnicas de diagnóstico más sensibles para ser utilizadas en el movimiento internacional de ganado hacia zonas libres de la enfermedad, que permitan la detección de animales portadores, así como para conocer la prevalencia de la enfermedad en las regiones tropicales y subtropicales. El gen msp5 está representado en el genoma como una simple copia, altamente conservado entre todas las especies de Anaplasma y todas las cepas de A. marginale, por lo que resulta un importante candidato para ser utilizado en el diagnóstico. En el presente trabajo se realizó la amplificación por PCR de dicho gen para la detección de Anaplasma marginale en 113 animales sin síntomas clínicos de la enfermedad, de los cuales 96 resultaron positivos para Anaplasma marginale, resultando el ensayo altamente sensible y específico.
Palabras clave: Anaplasma marginale; diagnóstico; PCR; gen msp5.
ABSTRACT
Anaplasma marginale is a rickettsia of Ehrlichias genogroup II. It parasites mature erythrocytes in bovines. There is not still an effective control method against the disease; thus it is of great importance to develop an immunogene able to prevent infection by this pathogen. There is also a need of having more sensitive diagnostic techniques which allow the detection of carrier animals in order to be used in cattle international movement towards areas free of the disease; as well as to know the prevalence of the disease in tropical and subtropical regions. Gene msp5 is represented in the genome as a simple copy; it is highly preserved among all Anaplasma species and all A. marginale strains; thus it is an important candidate for being used in diagnostic. In this paper, the amplification by PCR of such gene was carried out for the detection of Anaplasma marginale in 113 animals without clinical symptoms, from which 96 were positive to this microorganism, being the assay highly sensitive and specific.
Key words: Anaplasma marginale; diagnosis; PCR; msp5 gene.
INTRODUCCIÓN
Anaplasma marginale es una rickettsia del genogrupo II de las Ehrlichias, que parasita los eritrocitos maduros del ganado bovino. Los animales que sobreviven a la anaplasmosis bovina aguda, después de una infección primaria, permanecen persistentemente infectados de por vida, independientemente de re-exposiciones a la rickettsia (1), sirviendo como reservorio para la transmisión de la enfermedad en el campo (2).
El control de la anaplasmosis bovina requiere de una vacuna efectiva y de la identificación exacta de los animales portadores. Los dos métodos recomendados para la detección de éstos son la identificación del ADN de la rickettsia en sangre, mediante técnicas moleculares y la detección en suero de anticuerpos específicos contra A. marginale (3). Este último se hace difícil por los métodos serológicos de rutina (4), por lo que contar con técnicas de mayor sensibilidad y especificidad, es una necesidad.
Trabajos previos han demostrado que la proteína MSP5 de A. marginale resulta de gran utilidad en ensayos de formato ELISA (5,6), ya que elimina los problemas señalados de los antígenos crudos, utilizados en otras técnicas como la aglutinación en tarjeta. El gen msp5 es de gran utilidad para el diagnóstico molecular, mediante PCR, en animales persistentemente infectados (5). Este gen y su producto de expresión son recomendados para el monitoreo de animales durante el movimiento internacional de ganado y en programas de control de la anaplasmosis bovina (7).
Teniendo en cuenta las potencialidades del gen msp5 y la sensibilidad y especificidad que ofrece la PCR, para ser utilizados en el diagnóstico, el objetivo del presente trabajo fue desarrollar un ensayo de PCR a partir de la amplificación del gen msp5, para demostrar la presencia de A. marginale en muestras de bovinos en explotación.
MATERIALES Y MÉTODOS
Muestras trabajadas: Se muestreó un total de 113 animales hembras, procedentes de tres establecimientos ganaderos de la provincia La Habana que realizan las actividades de cría con propósito lechero y se recogieron los datos que se muestran en la Tabla 1. La sangre utilizada para el diagnóstico por frotis sanguíneo se obtuvo mediante punción de una vena de la cara interna de la oreja, previo lavado de la piel, con aguja de calibre 26 X 1/2, desechando las primeras gotas (8). La sangre para el diagnóstico por PCR se obtuvo mediante punción de la vena yugular, utilizando agujas hipodérmicas calibre 40 X 15 y tubos de cristal de fondo plano (Polylabo) conteniendo 10 mg de EDTA como anticoagulante. La sangre se lavó con PBS y se conservó a -20°C en crioviales (Neogene) con 30% de glicerol (9).
Controles positivos: Se seleccionaron cinco bovinos de la raza Siboney, de seis a siete meses de edad, seronegativos a Babesia spp. por inmunofluorescencia indirecta (10) y a A. marginale por aglutinación en placa (11). Estos animales se esplenectomizaron y se mantuvieron en estabulación con pienso y forraje verde. Durante 25 días se les realizó chequeo de hemoparásitos por frotis sanguíneo (8). Posteriormente se les inoculó por vía intravenosa, 20 mL de un estabilizado del aislamiento Habana, preparado según Dalgliesh (12) y se mantuvieron inoculados hasta que alcanzaron un 50% de parasitemia, siguiendo el curso de la infección por tinción de los frotis sanguíneos con colorante de Giemsa (8). Cuando alcanzaron esta parasitemia, se les extrajo sangre, la cual se mezcló con 10 mg de EDTA como anticoagulante y se sometieron a tratamiento.
Controles negativos: Se trabajó con dos bovinos de la raza Siboney de seis a siete meses de edad, seronegativos a Babesia spp. por inmunofluorescencia indirecta (10) y a Anaplasma marginale por aglutinación en placa (11). Se esplenectomizaron y se mantuvieron en estabulación con pienso y forraje verde durante 25 días. Se les realizó chequeo de hemoparásitos y posteriormente se les extrajo sangre, siguiendo la metodología descrita previamente.
Purificación del ADN: El ADN de A. marginale se purificó siguiendo la metodología descrita por Ambrosio y Potgieter (13), a partir de 2 mL de sangre de los animales experimentalmente infectados, de los animales negativos y de los animales de campo. Los ADN de los aislamientos Florida e Idaho fueron gentilmente cedidos por el Dr. Guy H. Palmer (Departamento de Medicina Veterinaria y Parasitología, Washington State University y el ADN de Tripanosoma spp. fue cedido por el Laboratorio de Genética, CINVESTAV, México. La purificación del ADN de B. bovis y B. bigemina se realizó según el protocolo descrito por Fahrimal et al. (14). El ADN bovino se purificó según la metodología descrita por Osta et al. (15).
Amplificación del gen msp5 mediante PCR: La amplificación del gen se realizó mediante PCR, a partir de 50 ng de los ADN purificados, en una mezcla de reacción de 25 mL, contentiva de 2.5 mL de solución tampón 1X (Tris HCl 200 mM, pH 8.8, KCl 10 mM, MgCl2 2.5 mM, Triton X-100 0.1 %, Promega), 20 pmoles de los cebadores: 5´GTGTTCGTTGGGGTGTGATAGATGAG3´ y 5´TAAGAATTAAGCATGTGACCGCTGAC3´, 0.2 mM de los dNTPs (Promega) y 1U de Taq polimerasa 5 U/mL (Promega). Del ADN obtenido de las muestras de campo se utilizaron 5 ul para la realización del PCR. En todos los casos se utilizó el siguiente programa de amplificación: un ciclo de 94°C, 1 minuto; 30 ciclos de 94°C, 1 minuto, 53°C, 1 minuto, 72°C, 1 minuto; un ciclo de 72°C, 10 minutos. Todos los reactivos se manipularon en flujo laminar (BHA 72), utilizando puntas con filtros (Eppendorf) resistentes a los aerosoles.
Las amplificaciones se realizaron en un equipo Mastercycler personal (Eppendorf) y los resultados se visualizaron en geles de agarosa (Sigma) al 0.8 %, en tampón TBE 0.5X a 100 V y 50 mA en TBE 0.5X, en cámara de electroforesis Apparatus GNA-100 (Pharmacia) y fuente Shandom Southern (V500/150), durante una hora. Para la visualización del ADN los geles se tiñeron con bromuro de etidio 0,5 mg/mL, en tampón de corrida durante 30 minutos, se observaron en un transiluminador de luz ultravioleta MacroVue (Pharmacia) y se fotografiaron con cámara digital EPSON 3.3 Mega, PIXELS.
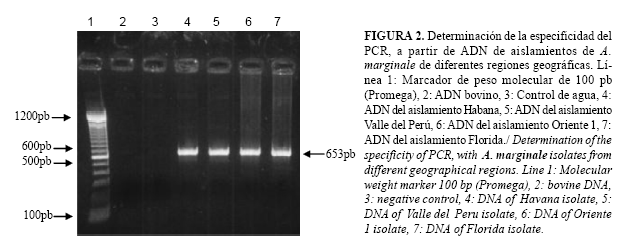
Especificidad del PCR con diferentes aislamientos de A. marginale: Para determinar la especificidad con diferentes aislamientos de A. marginale se utilizaron 50 ng de ADN de los aislamientos cubanos, Habana, Valle del Perú, Oriente 1 y del aislamiento Florida.
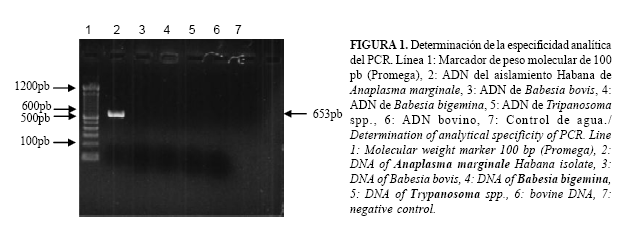
Especificidad analítica del PCR: Para determinar la especificidad con otros hemoparásitos, se realizó la amplificación del gen msp5 a partir del ADN de B. bovis, B. bigemina y Tripanosoma spp. Se realizó además la amplificación a partir de ADN bovino.
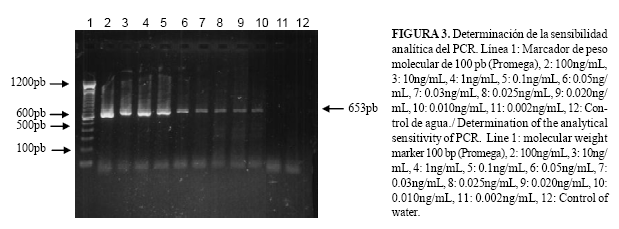
Sensibilidad analítica del PCR: Para determinar la más baja concentración de ADN purificado que puede ser detectado, se trabajó con ADN de un animal inoculado con la cepa Habana de A. marginale, con un 50 % de parasitemia, se realizaron diluciones seriadas desde una concentración de ADN de 100ng/uL hasta 0.002ng/uL. Los resultados se visualizaron en gel de agarosa al 1 % en TBE 0.5X.
RESULTADOS Y DISCUSIÓN
El PCR desarrollado resultó específico para A. marginale, (Fig. 1). No hubo amplificación a partir del ADN de otros hemoparásitos que pueden estar presentes en el bovino, ni de ADN bovino. La amplificación del gen fue posible en todos los aislamientos de A. marginale probados, apareciendo una banda de 653pb (Fig. 2).
Se demostró la sensibilidad del PCR utilizando diluciones de ADN de A. marginale, pudiendo detectarse hasta 0.01 ng/mL de ADN genómico de este hemoparásito (Fig. 3). Teniendo como base lo planteado por Eriks et al. (1), quienes determinaron que 0.01 ng de ADN es equivalente a aproximadamente 6000 organismos y conociéndose que cada eritrocito parasitado contiene un promedio de 6-12 organismos de A. marginale (16), entonces esto es equivalente a 1000 eritrocitos parasitados por mL de sangre bovina y un 0.000025% de parasitemia.
Con relación al frotis sanguíneo, nuestro ensayo resulta altamente sensible, si tenemos en cuenta que la tinción de los frotis de sangre con Giemsa sólo permite detectar niveles de parasitemia del orden del 0.1- 2% (17).
La amplificación del ADN del parásito tiene ventajas sobre otros métodos para la detección en animales persistentemente infectados con A. marginale, porque el parásito es difícil de detectar por los métodos de diagnóstico convencional (17) cuando el animal tiene bajos niveles de parasitemia. Herrero et al. (18), realizaron un estudio por PCR para la detección de A. marginale en Costa Rica, planteando que el PCR puede detectar bajos niveles de parasitemia en los animales persistentemente infectados, que no pueden ser detectados por los métodos convencionales.
Figueroa et al. (19), realizaron un PCR múltiple donde analizan los productos por hibridación con una sonda no radioactiva y encontraron que la sensibilidad fue de 0.00001% para B. bovis y B. bigemina y 0.0001% para A. marginale. Este sistema se utilizó posteriormente para estudios epidemiológicos en México y reportó en 420 muestras evaluadas una prevalencia de 59.6 % para A. marginale (20). Otros autores han utilizado el PCR para determinar la presencia de A. marginale en la hemolinfa (21) y en la saliva (22) de D. andersoni, demostrando así, tanto la función del vector como la vía de transmisión, al hallar el parásito primero en la hemolinfa y luego en la saliva de la garrapata.
La utilización del PCR en el diagnóstico de A. marginale permite avances significativos no solo en el diagnóstico de la parasitosis, permitiendo diseñar métodos de diagnóstico altamente eficientes (17), sino también en estudios epidemiológicos (20, 23) y en investigaciones de transmisión de especies de Anaplasma por rumiantes salvajes, como posibles reservorios de la anaplasmosis (24).
Al evaluar las muestras provenientes de los animales experimentalmente infectados todas resultaron positivas por PCR, amplificando la banda de 653pb, lo que denota la presencia de A. marginale en estos (datos no mostrados).
De un total de 113 animales estudiados, 22 resultaron positivos por tinción con Giemsa a los frotis de sangre, para un 19.46% y un total de 96 (84.95 %), resultaron positivos por PCR (Tabla 2) (Fig. 4). Estos animales sin síntomas clínicos de la enfermedad, donde fue posible detectar el microorganismo, son animales que se encuentran persistentemente infectados. En estos, las bandas amplificadas en el PCR fueron de diferente intensidad entre animales de un mismo grupo lo que puede estar relacionado con los niveles de parasitemia presentes y la habilidad para transmitir la enfermedad, lo que tiene un importante significado epidemiológico (1,22).

Los animales que resultaron positivos por frotis se encontraban, al parecer, en un pico de la rickettsemia, que hace que los niveles del parásito aumenten y puedan ser visualizados al microscopio (17).
El estado de portador de anaplasmosis, es un fenómeno que no está suficientemente entendido. Es posible que bajo determinados niveles de parasitemia animales individuales no sean capaces de trasmitir la anaplasmosis, sin embargo, existen factores externos e internos que pueden ser capaces de alterar los niveles de parasitemia de un individuo. Por ejemplo, la inmunosupresión de los animales infectados, el estrés ambiental o nutricional (25) y la infección con otras enfermedades puede provocar una recrudescencia de la anaplasmosis subclínica. Estos factores pueden contribuir con las fluctuaciones en los diferentes niveles de parasitemia de los animales dentro de un mismo grupo. Eriks et al. (1), determinaron que el número de eritrocitos infectados presente en los animales persistentemente infectados variaba, con niveles de parasitemia entre 0.0025 % y 0.000025 % y que dentro de un mismo animal estos valores fluctuaban en el tiempo. Con el sistema utilizado en el presente estudio, se puede detectar hasta 0.000025% de parasitemia, en animales sin síntomas clínicos de la enfermedad, sin embargo, se puede aumentar la sensibilidad del sistema, con la utilización de PCR dobles. Esto permitirá detectar los animales portadores con más bajos niveles de parasitemia, pues según Torioni y Echaide (26) en los bovinos que se recuperan de la fase aguda de la enfermedad y que permanecen persistentemente infectados, se producen ciclos continuos de rickettsemia con intervalos aproximados de 5 semanas, con valores entre 0.1% y 0.0000001% ó menores, de eritrocitos parasitados. Estos ciclos están asociados a variaciones antigénicas que ocurren en las moléculas de la superficie de A. marginale, que caracterizan a nuevas poblaciones del parásito resistentes a la respuesta inmune generada por la anterior, como un mecanismo para evadir el sistema inmune (26,27). Las combinaciones genéticas para generar estos nuevos epítopes, resultan más complejas en el transcurso de la parasitemia persistente, en respuesta a la presión de selección ejercida por la respuesta inmune (28).
La presencia de un número elevado de animales positivos a A. marginale, sin síntomas clínicos de la enfermedad, en estas unidades de la provincia La Habana puede estar dado por los métodos de control de ixódidos utilizados, que combinan la utilización de la vacuna Gavac, con baños garrapaticidas programados, según las características de las unidades, los cuales garantizan que los animales posean un nivel moderado de garrapatas, suficiente para asegurar la infección temprana de los terneros, los que poseen una resistencia innata, aún en ausencia de anticuerpos calostrales hasta cerca de los nueve meses de edad. Durante esta edad los animales adquieren la infección sin presentar signos aparentes de enfermedad y la inmunidad resultante, una vez establecida, es mantenida en el ganado adulto de por vida, mediante re-infecciones, las que son inaparentes clínicamente, por lo que se asume que un animal que se recobra de la enfermedad se convierte en portador (29,30,31).
En términos experimentales, el estado de portador significa que los eritrocitos infectados con A. marginale continúan circulando, a niveles no detectables, por largos períodos de tiempo, quizás durante toda la vida del animal, siendo relativamente inmunes a infecciones futuras. Este estado se caracteriza por una actividad de anticuerpos generalmente constante, y la detección no frecuente, en el microscopio, de eritrocitos infectados (29). El desarrollo de técnicas moleculares, especialmente el PCR, ha hecho posible la detección de bajos niveles de infección en animales portadores (32,33).
EL uso del gen msp5 en el ensayo de PCR propuesto, permitirá contar con una técnica sensible y específica para la detección de A. marginale en animales sin síntomas clínicos de la enfermedad, lo que constituye una poderosa herramienta para la importación y exportación de ganado. Así, Palmer y McElwain, (7), proponen el gen msp5 y la proteína para ser utilizados en el desarrollo de técnicas de detección, en el movimiento internacional de ganado.
REFERENCIAS
1. Eriks IS, Palmer GH, McGuire TC, Barbet AF. Detection and quantification of Anaplasma marginale in carrier by using a nucleic acid probe. Clin Microbiol. 1989;27:279-284.
2. Eriks IS, Stiller D, Palmer GH. Impact of persistent Anaplasma marginale rickettsemia on tick infection and transmission. J Clin Microbiol. 1993;31:2091-2096.
3. Knowles DP, Torioni ES, Palmer GH, McGuire TC, Stiller D, McElwain TF. Antibody against an Anaplasma marginale MSP5 epitope common to tick and erythrocyte stages identifies persistently infected cattle. Clin Micribiol. 1996;34:2225-2230.
4. Nakamura Y, Shimizu S, Minami T, Ito S. Enzime-linked immunosorbent assay using solubilised antigen for detection of antibodies to Anaplasma marginale. Trop Anim Hlth Prod. 1989;20:259-266.
5. Torioni ES, Knowles DP, McGuire TC, Palmer GH, Suarez CE, McElwain TF. Detection of cattle naturally infected with Anaplasma marginale in an endemic region using nested PCR and recombinant MSP5-cELISA. Clin Microbiol. 1998;36:777-782.
6. Ngeranwa JJN, Shompole SP, Venter EH, Wambugu A, Crafford JE, Penzohorn BL. Detection of Anaplasma antibodies in wildlife and domestic species in wildlife-livestock interface areas of Kenya by major surface protein 5 competitive inhibition enzyme-linked immunosorbent assay. J Vet Res. 2008;75:199-205.
7. Palmer GH, McElwain TF. Molecular bases for vaccine development against anaplasmosis and babesiosis. Vet Parasit. 1995;57:233-253.
8. Gainer JH. Demonstration of Anaplasma marginale with the fluorescent dye, acridine orange; comparisons with the complement-fixation test and Wright stain. Am J Vet Res. 1961;22:882-886.
9. Alonso M, Blandino T. Anaplasmosis bovina. Sociedad Cubana de Parasitología. Ediciones del Consejo Científico Veterinario de Cuba. 1988; 2-19.
10. Rodríguez J, Alonso M, Blandino T, Abreu R, Gómez E. Manual de Técnicas Parasitológicas. Ciudad de la Habana, Ediciones ENPES, 1987.
11.Teclaw RF, García Z, Romo Z, Wagner GC. Incidente of babesiosis and anaplasmosis infection in cattle sampled monthly in the Mexican states of Nuevo Leon and San Luis. Prev Vet Med. 1985;3:427-435.
12.Dalgliesh RJ. Consideraciones actuales sobre las babesiosis y anaplasmosis del Ganado bovino. Veterinaria. 1984; No. 13.
13.Ambrosio RE, Potgieter FT. The genome of Anaplasma: DNA composition and DNA/DNA hybridization. J Vet Res. 1987;54:53-65.
14.Fahrimal Y, Goff WL, Jasmer DP. Detection of Babesia bovis carrier cattle by using polymerase chain reaction amplification of parasite DNA. J Clin Microbiol. 1992;30:1374-1379.
15.Osta R. Caracterización genética de proteínas lácteas y sexaje de embriones en ganado vacuno mediante la aplicación de la biotecnología al análisis del DNA. Tesis Doctoral, Facultad de Veterinaria.
16.Goff W, Barbet AF, Stiller D, Palmer GH, Knowles D, Kocan K, et al. Detection of Anaplasma marginale infected tick vectors by using a cloned DNA probe. Proc Natl Acad Sci. 1998;85:919-92.
17.Melman S, Hart Y, Giardina S. El uso de la biología molecular en el diagnóstico de Anaplasma marginale. 2004; (http://investigación.unefm.edu.ve).
18.Herrero MV, Pérez E, Goff WL, Torioni ES, Knowles DP, McElwain TF, et al. Prospective study for the detection of Anaplasma marginale Theiler, 1911 (Rickettsiales: Anaplasmataceae) in Costa Rica. Ann NY Acad Sci. 1998;849:226-233.
19.Figueroa JV, Chieves LP, Johnson GS, Buening GM. Multiplex polymerase chain reaction based assay for the detection of Babesia bigemina, Babesia bovis and Anaplasma marginale DNA in bovine blood. Vet Parasitol. 1993;50:69-81.
20.Figueroa JV, Álvarez JA, Ramos JA, Vega CA, Buening GM. Use of multiplex polymerase chain reaction-based assay to conduct epidemiological studies on bovine hemoparasites in Mexico. Rev Elev Med Vet Pays Trop. 1993;46(1-2):71-75.
21.Stich RW, Bantle JA, Kocan KC, Fekete A. Detection of Anaplasma marginale (Rickettsiales: Anaplasmataceae) in hemolymph of Dermacentor anderosni (Acari: Ixodidae) with the polymerase chain reaction. J Med Entomol. 1993;30:781-788.
22.Stich RW, Sauer JR, Bantle JA, Kocan KC. Detection of Anaplasma marginale (Rickettsiales: Anaplasmataceae) in secretagogue-induced secretions of Dermacentor andersoni (Acari: Ixodidae) with the polymerase chain reaction. J Med Entomol. 1993;30:789-794.
23.Figueroa JV, Alvarez JA, Ramos JA, Rojas EE, Santiago C, Mosqueda JJ, et al. Bovine babesiosis and anaplasmosis follow-up on cattle relocated in an endemic area for hemoparasitic diseases. Ann NY Acad Sci. 1998;849:1-10.
24.Lew AE, Bock RE, Minchin CM, Masaka S. A msp1a polimerase chain reaction assay for spesific detection and differentiation of Anaplasma marginale isolates. Vet Microbiol. 2002;86:325-335.
25.Corrier DE, Wagner GG, Adams LG. Recrudescence of Anaplasma marginale- induced immunosuppression with cyclophosphamide. Am J Vet Res. 1981;42:19-21.
26.Torioni ES, Echaide IE. (2003). Anaplasmosis de los bovinos. 2003. (http://www.veterinario.sursf.com.ar/muestrapublicacion.php).
27.Palmer GH, Futse JE, Leverich CK, Knowles DP, Rurangirwa FR, Brayton KA. Selection for Simple Major Surface Protein 2 Variants during Anaplasma marginale Transmission to Immunologically Naive Animals. Infect Immun. 2007;75:1502-1506.
28.Futse JE, Brayton KA, Knowles DP, Palmer GH. Structural basis for segmental gene conversion in generation of Anaplasma marginale outer membrane protein variants. Molecular Microbiol. 2005;57(1):212-221.
29.Wagner G. Immunoglobulin responses of cattle associated with recrudescent Anaplasma marginale infections. Refuah Vet. 1981;38:37-44.
30.Benavides OE. Consideraciones con relación a la Epizootiología de Anaplasmosis y Babesisosis en los bovinos. Revista ACOVEZ. 1995;9(31):4-11.
31.Cetra B, Ramírez LM, Vangini V. INTA, Noticias y Comentarios. 2000; No. 339.
32.Lew A, Jorgensen W. Molecular approaches to detect and study the organisms causing bovine tick borne diseases: babesiosis and anaplasmosis. African J Biotechnol. 2005;4(4):292-302.
33.Rodríguez DS, García OM, Jiménez OR, Vega CA. Molecular epidemiology of bovine Anaplasmosis with a particular focus in México. Infection, Genetics and Evolution. 2009;9:1092-1101.
(Recibido 14-6-2010; Aceptado 9-10-2010)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}