










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Salud Animal
versión impresa ISSN 0253-570X
Rev Salud Anim. vol.34 no.1 La Habana ene.-abr. 2012
Artículo original
DETECCIÓN POR PCR DE Anaplasma marginale EN BÚFALOS DE LA REGIÓN OCCIDENTAL DE CUBA
DETECTION BY PCR Anaplasma marginale IN BUFFALOES OF THE WESTERN REGION OF CUBA
Belkis Corona*, D. Obregón**, Siomara Martínez*, Ivette Espinosa*, A. Henrique Fonseca***, E. Roque**
*Centro Nacional de Sanidad Agropecuaria. Apartado 10, San José de las Lajas, Mayabeque, Cuba. Correo electrónico: bcorona@censa.edu.cu. **Universidad Agraria de La Habana. Autopista Nacional y carretera de Tapaste, San José de las Lajas, Mayabeque, Cuba. ***Universidad Federal Rural de Rio de Janeiro, RJ, Brasil
RESUMEN
La especie bubalina, introducida en Cuba, ha mostrando excelentes resultados productivos y reproductivos. La situación de salud en ellos ha estudiado poco, aunque se reconoce su resistencia natural a varias enfermedades informadas en el país, incluidas las hemoparasitosis. Los búfalos también son susceptibles a la infección por Anaplasma marginale, pero la anaplasmosis clínica difícilmente se presenta, permaneciendo como portadores asintomáticos, lo que facilita la transmisión a los bovinos. El presente trabajo tuvo como objetivo determinar la presencia de A. marginale en búfalos de la región occidental de Cuba. Para ello se desarrolló un ensayo de nPCR, a partir de la amplificación del gen msp5 de este hemoparásito. Se analizaron 52 muestras de ADN, extraído de sangre de animales seleccionados al azar en seis granjas del occidente de Cuba, a los que se les realizó Frotis Sanguíneo. Del total de muestras analizadas, 47 muestras (90,4%) resultaron positivas por el nPCR, visualizándose en todos los casos una banda de 345 pb, mientras que sólo cinco animales (9,6%) fueron positivos en el Frotis Sanguíneo. Uno de los fragmentos amplificados fue secuenciado y la secuencia obtenida fue depositada en el GenBank con Número de Acceso JF270381, mostrando un alto por ciento de identidad con secuencias depositadas de Anaplasma marginale de diferentes regiones del mundo.
Palabras clave: Anaplasma marginale; PCRn; msp5.
ABSTRACT
The buffalo species was recently introduced in Cuba, showing excellent productive and reproductive performance. Their health situation has been little studied, although their natural resistance to various diseases reported in the country, including haemoparasitosis has been recognized. Buffaloes are also susceptible to infection with Anaplasma marginale, but hardly present clinical anaplasmosis, remaining asymptomatic carriers, which facilitates transmission to cattle. This study is aimed at determining the presence of A. marginale in buffaloes of the western region of Cuba. For this purpose, an nPCR test, based on msp5 gene amplification of this hemoparasites, was developep. Fifty-two DNA samples, extracted from animal blood, randomly selected from 6 farms in western Cuba, which underwent blood smear were analized. From the total samples analyzed, 47 (90.4%) were positive by nPCR, visualizing in all cases a band of 345 bp, while only 5 animals (9.6%) were positive in blood smear. One of the fragment amplified was sequenced and the sequence obtained was deposited in GenBank under Accession Number JF270381, showing a high percentage of identity with Anaplasma marginale deposited sequences in different regions of the world.
Key words: Anaplasma marginale; nPCR; msp5.
INTRODUCCIÓN
En Latinoamérica la existencia estimada de búfalos asciende a más de 3,8 millones de cabezas (1) y en el mundo supera los 2020 millones (2). La especie bubalina (Bubalus bubalis) fue introducida en Cuba en la década de los 80, (3), como alternativa para la recuperación de leche, carne y su empleo en la tracción animal, garantizando su desarrollo con bajos insumos, adaptándose muy bien a nuestras condiciones de crianza (4) y en el 2009 alcanzaban la cifra de 63 050 cabezas (5).
Hasta la actualidad no se han realizado estudios que aborden la incidencia de las hemoparasitosis en los rebaños bubalinos. Tampoco existen reportes de focos o cuadros clínicos de estas patologías (Informe de Balance, 2009, IMV), predominando el criterio entre productores y personal veterinario, de que los búfalos son refractarios a las hemoparasitosis.
Los trabajos realizados hasta el momento indican la necesidad e importancia de caracterizar la situación epidemiológica de estas infecciones en los rebaños bubalinos (6,7,8,9), en especial, la identificación de animales portadores, por constituir un parámetro epidemiológico importante en el monitoreo y control de estas infecciones (10).
Los búfalos son susceptibles a muchas de las enfermedades y parásitos que afectan al bovino, pero el efecto en ellos es menos serio, en un mismo ecosistema (11). Anaplasma marginale es una rickettsia del genogrupo II de las Ehrlichias, que parasita los eritrocitos maduros del ganado bovino. Los animales que sobreviven a la anaplasmosis bovina aguda, después de una infección primaria, permanecen persistentemente infectados de por vida, independientemente de re-exposiciones a la rickettsia (12), sirviendo como reservorio para la transmisión de la enfermedad en el campo (13).
La enfermedad clínica es más notable en bovinos, pero otros rumiantes que incluyen búfalo de agua, bisonte, antílope africano, y ciervo se pueden convertir en persistentemente infectados con Anaplasma marginale (11).
El control de la anaplasmosis bovina requiere de una vacuna efectiva y de la identificación exacta de los animales portadores. Los dos métodos recomendados para la detección de estos son la identificación del ADN de la rickettsia en sangre, mediante técnicas moleculares y la detección de anticuerpos específicos contra A. marginale en suero (14). Este último se hace difícil por los métodos serológicos de rutina (15), por lo que contar con técnicas de mayor sensibilidad y especificidad, es una necesidad.
El gen msp5 es de gran utilidad para el diagnóstico molecular, mediante PCR, en animales persistentemente infectados (16). Este gen y su producto de expresión son recomendados para el monitoreo de animales durante el movimiento internacional de ganado y en programas de control de la anaplasmosis bovina (17).
Teniendo en cuenta las potencialidades del gen msp5 y la sensibilidad y especificidad que ofrece la PCR, para ser utilizados en el diagnóstico, el objetivo del presente trabajo fue desarrollar un ensayo de PCR a partir de la amplificación del gen msp5, para demostrar la presencia de A. marginale en muestras de búfalos en explotación en la región occidental de Cuba.
MATERIALES Y MÉTODOS
Muestras de sangre: Se tomaron muestras de sangre a 52 búfalos seleccionados al azar, de diferentes categorías, procedentes de varias unidades de producción bufalina, en cuatro provincias del occidente de Cuba (Tabla 1). Ningún caso presentaba manifestaciones clínicas de síndrome hemolítico.

Diagnóstico por frotis sanguíneo: A cada animal se le extrajo sangre periférica, perforando el borde interno de la oreja, previo lavado de la piel, con aguja de calibre 26 X 1/2, desechando las primeras gotas y se realizó frotis sanguíneo delgado. Las extensiones fueron llevadas al laboratorio, fijadas con metanol absoluto (Uni-Chem) y teñidas durante 30 min con Giemsa según Gainer (18). La lectura se hizo con microscopio óptico (Opton), empleando lente de inmersión de 100X.
Colecta de sangre: La sangre para el diagnóstico por PCR se obtuvo mediante punción de la vena yugular, utilizando agujas hipodérmicas calibre 40 X 15 y tubos de cristal de fondo plano (Polylabo) con 10 mg de EDTA como anticoagulante y se conservó -20°C en crioviales (Neogene), sin descongelarse, hasta su uso.
Purificacion del ADN: La extracción de ADN fue realizada según la metodología descrita por Ambrosio y Potgieter (19).
Como control positivo se utilizó el ADN del aislamiento Habana de A. marginale, obtenido a partir de sangre infectada, donada por el Grupo de Parasitología, CENSA. Esta sangre se extrajo de bovinos esplenectomizados infectados experimentalmente y que alcanzaron un 60% de parasitemia. La sangre fue lavada con PBS y conservada en 30% de glicerol a -20oC hasta su uso y el ADN se purificó según la metodología descrita por Ambrosio y Potgieter (19).
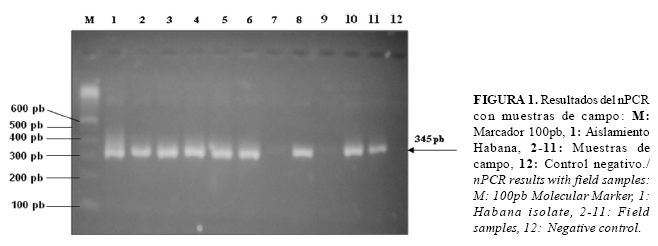
Realización del PCR: Para la realización del nPCR se utilizaron los cebadores descritos por Torioni et al. (16), con los que se amplifican dos fragmentos dentro del gen msp5 de A. marginale. Para el PCR1 los cebadores 1 y 2 (5´GCATAGCCTCCGCGTCTTTC3´; 5´TCCTCGCCTTGCCCCTCAGA3´) amplifican un fragmento de 458 pb, y para el PCR2 los cebadores 3 y 2 (5´TACACGTGCCCTACCGATTA3'; 5´TCCTCGCCTTGCCCCTCAGA3´) amplifican un fragmento de 345 pb dentro del primer fragmento amplificado. La síntesis de los cebadores se realizó en el Centro de Ingeniería Genética y Biotecnología (CIGB), La Habana, Cuba.
Ambos ensayos (PCR1 y PCR2) se realizaron en una mezcla de reacción de 25 µL, según lo descrito por Torioni et al. (16), con modificaciones. La mezcla de reacción contenía 2,5 µL de solución tampón 1X (Promega) (KCl 50mM; Tris-HCI 10mM, pH 8.3); 2.5mM de MgCl2, 200µM de los dNTPs (Promega), 10µM de los cebadores y 1µL de Taq Polimerasa (AmpliCEN). En el PCR1 se utilizaron 5µl de ADN de A. marginale, y en el PCR2 se utilizó 1µL del producto del PCR1. Todos los reactivos se manipularon en flujo laminar (IKEM), utilizando puntas con filtros (Eppendorff) resistentes a aerosoles. Las reacciones se desarrollaron en un Termociclador «Mastercycler Personal» (Eppendorf). Ambos ensayos se realizaron con el siguiente programa de amplificación: un ciclo de 95ºC, durante 3 minutos; 35 ciclos de 95ºC- 30 segundos, 61ºC- 1 minuto, 72ºC- 30 segundos y una extensión final de 10 minutos a 72ºC.
Visualización de los resultados del nPCR: Los resultados del nPCR se aplicaron en geles de agarosa (Sigma) al 2%, en tampón TBE 0.5 X, se corrieron a 100 Volts y 50 mA, en cámara de electroforesis «Maxiphor 2012» (LKB Bromma) durante 35 minutos; el tampón de corrida (TBE 0.5X) contenía Bromuro de Etidio (0.5 µg/mL). Los resultados se visualizaron en un transluminador de luz ultravioleta «Macro Vue» (Pharmacia) y se fotografiaron con cámara digital (Olympus) de 10 megapixel. Se utilizó un marcador de peso molecular de 100pb (Promega).
Análisis de restricción al fragmento amplificado en el nPCR: Se realizó un análisis in silico para determinar las posibles enzimas a emplear, para lo que se utilizó el programa «NEB cutter» (http://www.tols.neb.com/nebcutter), y como molde, la secuencia del gen msp5 del aislamiento «Habana» de A. marginale con número de acceso AY527217. Para confirmar que los fragmentos visualizados en el gel de agarosa, eran realmente de A. marginale, se seleccionaron muestras al azar y se les realizó análisis de restricción con EcoR I (Promega).
Secuenciación del fragmento amplificado: Uno de los fragmentos amplificados fue secuenciado utilizando el Kit Genome Labtm DTCS- Quick start kit, en el Equipo CEQtm 8800 (Beckman Coulter). La secuencia obtenida fue depositada en el GenBank con Número de Acceso JF270381 y se analizó el por ciento de identidad con el programa BlastN del NCBI.
RESULTADOS Y DISCUSIÓN
La primera determinación de A. marginale en sangre de búfalo se realizó a través de la visualización de los frotis sanguíneos, teñidos con Giemsa. Del total de 52 muestras analizadas, solo 5 (9,6%) resultaron positivas, encontrándose escasos eritrocitos parasitados por campo. Con el empleo del nPCR se visualizó la banda de 345 pb en 47 de las muestras analizadas, para un 90,4% (Figura 1, Tabla 2).
Esta diferencia en los resultados obtenidos por ambas técnicas confirma que el frotis sanguíneo no es lo suficientemente sensible para la identificación de animales portadores, ya que esta técnica solo detecta niveles de parasitemia, superiores a 106 EI/ml de sangre (20), mientras que en los animales portadores la parasitemia fluctúa entre 102 y 107 EI/ml de sangre (21), lo que limita que pueda ser usado para establecer un diagnóstico preciso en animales portadores.
El análisis in silico permitió identificar varias enzimas de restricción, cuyos sitios de corte están presentes en la secuencia nucleotídica del fragmento de msp5 amplificado por el nPCR (Figura 2). Se seleccionó la enzima de restricción EcoR1, cuyo sitio de corte se encuentra en la posición 152 del fragmento de 345 pb del gen msp5, amplificado en el nPCR.
Como resultado del análisis de restricción se obtuvieron bandas con tallas de 193pb y 152pb respectivamente en 25 muestras, seleccionadas al azar, del total de muestras positivas. En la Figura 3 podemos observar que las bandas obtenidas mostraron patrones de peso según lo esperado, siendo otra evidencia de la especificidad del ensayo.
La identificación de Anaplasma marginale fue confirmada con la secuenciación de uno de los fragmentos de 345 pb amplificado. La secuencia, depositada en el GenBank con número de acceso JF270381, se comparó con secuencias previamente informadas para A. marginale, A. centrale, A. ovis y A. phagocytophilum (Fig. 4), mostrando entre un 94%-99% de identidad con cepas de A. marginale, un 88% de identidad para A. centrale, un 86% para A. ovis y un 69% para A. phagocytophilum. En la comparación con secuencias de cepas de A. marginale mostró los más altos por cientos de identidad con la cepa Habana, aislada de bovinos (99%) y las cepas de Estados Unidos (97%).
Strik et al. (22) encontraron alta conservación del gen msp5 entre aislamientos de A. pahogocytophilum de diferentes regiones de Estados Unidos y Europa, reafirmando además que el gen msp5 de Anaplasma sp. y su ortólogo map2 de Ehrlichia spp. son altamente conservados dentro los géneros Anaplasma y Ehrlichia, aspecto planteado por otros autores (23,24).
Por otra parte, Dreher et al. (25) informaron un 64% de similitud en la secuencia deducida de aminoácidos de la proteína MSP5 de A. phagocytophilum y la del aislamiento Habana de Anaplasma marginale, con la que nuestra secuencia nucleotídica tiene un 99% de identidad. En nuestro caso existe un 69% de identidad de la secuencia nucleotídica del gen msp5 con este aislamiento de A. phagocytophilum para el que se informó reactividad inmunológica cruzada entre la proteína MSP5 de Anaplasma marginale y la Anaplasma phagocytophilum, recomendando el uso del PCR para el caso de muestras que resulten positivas por IFA y cELISA.
Molad et al. (26) encontraron un 86.8% de identidad entre la secuencia nucleotídica del gen msp5 de la cepa vacunal de A. centrale y una cepa de Anaplasma marginale, aislada de bovino, lo que coincide con nuestros resultados.
Con estos resultados se verificó que el ensayo de nPCR constituye una herramienta diagnóstica de precisión para la identificación de animales portadores de la infección por Anaplasma marginale, coincidiendo con los resultados de Torioni et al. (16) quienes lo emplearon en muestras de sangre de bovinos. Los resultados obtenidos confirman que el ensayo puede ser empleado en otras especies, coincidiendo con lo planteado por de la Fuente et al. (27,28) al realizar la detección de Anaplasma marginale en muestras de sangre de Bisonte Americano (Bison bison) y ciervo ibérico, respectivamente. Los resultados obtenidos por estos autores demostraron que el ciervo ibérico estaba naturalmente infectado con Anaplasma, y que por lo tanto podrían servir como reservorio del patógeno, identificando a Hyalomma marginatum y Rhipicephalus bursa como vectores biológicos potenciales para A. marginale, y que pueden efectuar la transmisión de A. marginale entre el ciervo y la población bovina.
El hallazgo de Anaplasma marginale infectando los búfalos en el occidente de Cuba constituye un resultado novedoso, siendo el primer informe de este parásito en la especie en el país. Estos resultados coinciden con los obtenidos por Jacobo et al. (29), Rajput et al. (11) y Gomes et al. (30), quienes detectaron la infección con Anaplasma marginale y la enfermedad en la forma crónica o asintomática en búfalos de otras regiones del mundo.
Los resultados de la distribución de casos según las diferentes provincias, aun cuando no se trata de un estudio epidemiológico, permiten suponer que la infección está ampliamente distribuida en todos los rebaños de la región, como ocurre en otros países, como Argentina y Colombia (7).
La infección de Anaplasma marginale en búfalos ya fue informada en Argentina por Jacobo et al. (7; 8), pero no se evaluó su impacto sanitario en la especie. Sin embargo, Jacobo et al. (29), infieren que esto podría constituirse en todo un problema sanitario para esta especie y particularmente para los bovinos que son sus hospederos primarios, oficiando el búfalo en este caso, como fuente de infección, manteniendo el agente infeccioso y comportándose como reservorio en zonas enzoóticas.
Gomes et al. (30), encontraron que los búfalos mostraron altos niveles de anticuerpos durante las primeras 24 horas después de succionar calostro, indicando la presencia de inmunidad calostral pasiva, por lo que concluyeron que los búfalos son capaces de desarrollar una respuesta inmune humoral específica contra Anaplasma marginale, siendo considerados portadores de este parásito.
Fyumagwa et al. (31) encontraron 13 especies diferentes de garrapatas en búfalos de la región de Ngorongoro Cráter y de éstas en 4 especies se detectó la presencia de Anaplasma marginale. Khan et al. (32) estudiaron la presencia de hemoparásitos en sangre de bovinos y búfalos y encontraron que entre los animales infectados la mayoría presentaba Anaplasma marginale y en el caso de los búfalos el 80% de los animales estudiados estaban infectados.
Los resultados obtenidos son de gran importancia para los programas de control de la anaplasmosis que se desarrollan en nuestro país y en la región del Caribe, si tenemos en cuenta que los búfalos, pueden constituir importantes reservorios para la trasmisión de la enfermedad al bovino.
REFERENCIAS
1. FAO. Principales países productores de ganado bufalino. Disponible en: http://www.fao.org/ag/AGa/AGAP/WAR/warally/v1650b/v16500bOc.htm. 2000.
2. Anuario. Producción bufalina. Dirección de ganadería. Área bufalinos. Disponible en: http://www.Sagpya.mencor.ar. 2008.
3. Fundora O, González ME. Performance of primiparous buffaloes and their progeny. Proc. VI World Buffalo Congress. Maracaibo, Venezuela. 2001;137.
4. Mitat A. Búfalos de agua en Cuba. Origen y evolución. Rev ACPA. 2009;3:45.
5. Simon L, Galloso M. Presencia y perspectivas de los búfalos en Cuba. Pastos y Forrajes [online]. 2011;34(1):3-20.
6. Banerjee DP, Momim RR, Samanta RA. Cross transmisión of Babesia bigemina from cattle (Bos taurus x Bos indicus) to buffalo (Bubalus bubalis). In: Wolrd Buffalo Congress, 1998, New Delhi. Council of Agricultural Research, p. 329.
7. Jacobo RA, Seggiaro S, Cipolini MF, Storani CA, Martínez DE, De Sa Da Silva A, et al. Primera comunicación en Argentina del Complejo tristeza del bovino en búfalos. Resumen de IIª Simposio de Búfalos de las Américas. Corrientes (Argentina), 2004.
8. Jacobo RA, Cipolini MF, Storani CA, Martínez DE, Martínez EI. Infección con el Complejo tristeza del bovino en búfalos. Rev Medicina Veterinaria. 2004;85(5):203-204.
9. Ferreri L, Benitez D, Dominguez M, Rodríguez A, Asenzo G, Mesplet M, et al. Water Buffalos as Carriers of Babesia bovis in Argentina. N.Y. Acad. Sci. Anim Biodiv and Emerging Diseases. 2008;1149:149-151.
10.Almeria S, Castella J, Ferrer D, Ortuño A, Estrada-Peña A, Gutierrez JF. Bovine piroplasms in Minorca (Belearic Islands Spain): a comparison of PCR-based and light microscopy detection. Vet Parasitol. 2001;99:249-259.
11.Rajput I, Hu S, Arijo AG, Habib M, Khalid M. Comparative study of Anaplasma parasites in tick carrying buffaloes and cattle. J Zhejiang Univ SCI, 6B. 2004;1057-1062.
12.Eriks IS, Palmer GH, McGuire TC, Barbet AF. Detection and quantification of Anaplasma marginale in carrier by using a nucleic acid probe. J Clin Microbiol. 1989;27:279-284.
13.Eriks IS, Stiller D, Palmer GH. Impact of persistent Anaplasma marginale rickettsemia on tick infection and transmission. J Clin Microbiol. 1993;31:2091-2096.
14.Knowles DP, Torioni ES, Palmer GH, McGuire TC, Stiller D, McElwain TF. Antibody against an Anaplasma marginale MSP5 epitope common to tick and erythrocyte stages identifies persistently infected cattle. J Clin Micribiol. 1996;34:2225-2230.
15.Nakamura Y, Shimizu S, Minami T, Ito S. Enzime-linked immunosorbent assay using solubilised antigen for detection of antibodies to Anaplasma marginale. Trop Anim Health Prod. 1989;20:259-266.
16.Torioni ES, Knowles DP, McGuire TC, Palmer GH, Suarez CE, McElwain TF. Detection of cattle naturally infected with Anaplasma marginale in an endemic region using nested PCR and recombinant MSP5-cELISA. J Clin Microbiol. 1998;36:777-782.
17.Palmer GH, McElwain TF. Molecular bases for vaccine development against anaplasmosis and babesiosis. Vet Parasit. 1995;57:233-253.
18.Gainer JH. Demonstration of Anaplasma marginale with the fluorescent dye, acridine orange; comparisons with the complement-fixation test and Wright stain. Am J Vet Res. 1961;22:882-886.
19.Ambrosio RE, Potgieter FT. The genome of Anaplasma: DNA composition and DNA/DNA hybridization. J Vet Res. 1987;54:53-65.
20.Gale RC, Dimmock CM, Gartside M, Leatch G. Anaplasma marginale detection of carrier cattle by PCR. Int J Parasitol. 1996;26:1103-1109.
21.Palmer GH, Rurangirwa FR, Kocan KM, Brown WC. Molecular basis for vaccine development against ehrlichial pathogen Anaplasma marginale. Parasitol Today. 1999;7:281-286.
22.Strik NI, Alleman AR, Barbet AF, Sorenson HL, Wamsley HL, Gaschen FP, et al. Characterization of Anaplasma phagocytophilum Major Surface Protein 5 and the Extent of Its Cross-Reactivity with A. marginale. Clinic and Vaccine Immunol. 2007;14(3):262-268.
23.Alleman AR, McSherry LJ, Barbet AF, Breitschwerdt EB, Sorenson HL, Bowie MJ, et al. Recombinant major antigenic protein 2 of Ehrlichia canis: a potential diagnostic tool. J Clin Microbiol. 2001;39:2494-2499.
24.Knowles TT, Alleman AR, Sorenson HL, Marciano DC, Breitschwerdt EB, Harrus S, et al. Characterization of the major antigenic protein 2 of Ehrlichia canis and E. chaffeensis and its application for the serodiagnosis of ehrlichiosis. Clin Diagn Lab Immunol. 2003;10:520-524.
25.Dreher UM, de la Fuente J, Hofmann-Lehmann R, Meli ML, Pusterla N, Kocan KM, et al. Serologic Cross-Reactivity between Anaplasma marginale and Anaplasma phagocytophilum. Clin Diagn Lab Immunol. 2005;12(10):1177-1183.
26.Molad T, Brayton KA, Palmer GH, Michaeli S, Shkap V. Molecular conservation of MSP4 and MSP5 in Anaplasma marginale and A. centrale vaccine strain. Vet Microbiol. 2004;100:55-64.
27.de la Fuente J, Vicente J, Höflea U, Ruiz-Fons F, Fernández de Mera IG, Van Den Bussche RA, et al. Anaplasma infection in free-ranging Iberian red deer in the region of Castilla-La Mancha, Spain. Vet Microbiol. 2004;100:163-173.
28.de la Fuente J, Golsteyn Thomas EJ, Van Den Bussche RA, Tanaka EE, Druhan SE, Kocan KM. Characterization of Anaplasma marginale isolated from North American bison. Appl Environ Microbiol. 2003;69:5001-5005.
29.Jacobo RA, Cipolini MF, Storani CA, Martínez DE, Martínez EI. Anaplasmosis en búfalos: diagnóstico clínico y de laboratorio, Resultados preliminares. 2005.
30.Gomes RA, Machado R, Starke-Buzetti WA, Bonesso MA. Respuosta immune-humoral de búfalos (Bubalus bubalis) contra Anaplasma marginale (THEILER, 1910). Rev Bras Parasitol Vet. 2008;17(2):73-80.
31.Fyumagwa RD, Simmler P, Meli ML, Hare R, Hofmann-Lehmann R, Lutz H. Prevalence of Anaplasma marginale in different tick species from Ngorongoro Crater, Tanzania. Vet Parasitol. 2009;161:154-157.
32.Khan MQ, Zahoor A, Jahangir M, Ashraf Mirza M. Prevalence of blood parasites in cattle and buffaloes. Pakistan Vet J. 2004;24(4):193-194.
(Recibido 19-9-2011; Aceptado 3-1-2012)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}