










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Salud Animal
versión impresa ISSN 0253-570X
Rev Salud Anim. vol.34 no.2 La Habana mayo-ago. 2012
ARTÍCULO ORIGINAL
Polimorfismo genético en los receptores de la hormona del crecimiento y prolactina en el Siboney de Cuba. Desarrollo de metodologías
Genetic polimorphism in growth hormone receptors and prolactin in Siboney de Cuba. Development of methodologies
A. AcostaI, R. RondaI, F. LópezII, Z. FernandesIII, M.A. Gomes-FilhoIII, O. UffoI*, S.B.P. BarbosaII
ILaboratorio de Genética Molecular (GenMol), Centro Nacional de Sanidad Agropecuaria (CENSA) PO Box 10, San José de las Lajas, CP 32700, Mayabeque, Cuba. Correspondencia del autor: Odalys Uffo Reinosa (Fax: + 5347-868104, Tel: + 5347-863145, Correo electrónico: uffo@censa.edu.cu);
IIPrograma de Gerenciamento de Rebanhos Leiteiros do Nordeste (PROGENE), Departamento de Zootecnia, UFRPE, Pernambuco, Brasil;
IIILaboratório Fisiologia Animal Molecular Aplicada (FAMA), Departamento de Morfologia e Fisiologia Animal, UFRPE, Pernambuco, Brasil
RESUMEN
Se han realizado estudios previos para la determinación del genotipo del SNP en el exón ocho del gen receptor de la hormona del crecimiento (GHR), causante de la sustitución aminoacídica F279Y y el SNP en el exón tres del gen receptor de la prolactina (PRLR) que provoca la sustitución aminoacídica S18N. Se describe que ambos polimorfismos están fuertemente asociados al rendimiento y componentes lácteos. Nuestro objetivo fue describir dos nuevas metodologías para identificar dichos polimorfismos y determinar las frecuencias génicas para ambos loci en la raza Siboney de Cuba (N=130). Las metodologías propuestas fueron la amplificación creando el sitio de restricción (ACRS) en el locus GHR y la reacción en cadena de la polimerasa (PCR) combinada con polimorfismo de longitud de los fragmentos de restricción (RFLP) en el locus PRLR. Se identificaron los tres genotipos en ambos loci, con frecuencias en el locus GHR de 0.517, 0.428 y 0.025 para los genotipos FF, FY y YY respectivamente, mientras que la frecuencia de los genotipos SS, SN y NN fueron de 0.686, 0.24 y 0.074 en el locus PRLR. Ambos loci se encuentran desviados de la condición de equilibrio Hardy-Weinberg (p<0.05).
Palabras clave: bovino, gen GHR, gen PRLR, frecuencia alélica.
ABSTRACT
There are several previous studies on the typing of SNP in exon eight of the growth hormone receptor gene (GHR) causing F279Y amino acid substitution, and SNP in exon three of the prolactin receptor gene (PRLR) causing S18N amino acid substitution. Both polymorphisms have been strongly associated with milk yield and milk components. The aim of the present study was to describe two new methods to identify these polymorphisms and determine gene frequencies for both loci in the breed Siboney de Cuba (N = 130). The genetic polymorphism of the GHR locus was detected by amplification creating restriction sites (ACRS), and the genetic polymorphism of the PRLR locus was detected by polymerase chain reaction (PCR) combined with restriction fragment length polymorphism (RFLP). Three genotypes were identified in both loci, with frequencies in the GHR locus of 0.517, 0.428 and 0.025 for genotypes FF, FY and YY respectively; and the frequency of genotypes SS, NS and NN were 0.686, 0.24 and 0.074 in the PRLR locus. Both loci do not fit Hardy-Weinberg equilibrium in this population (p <0.05).
Key words: cattle, GHR gene, PRLR gene, allele frequency.
INTRODUCCIÓN
Por las diversas funciones que tienen en el organismo, los genes que codifican para la hormona del crecimiento (GH) y la prolactina (PRL) han sido incluidos en numerosos estudios. De la GH se conoce su especial participación en crecimiento pos-natal y producción de leche en mamíferos (1,2). La PRL es un regulador esencial del desarrollo de la glándula mamaria, que actúa en sinergismo con otras hormonas durante la pubertad y la preñes (3,4,5,6). La actividad de estas hormonas es mediada por receptores ubicados en la membrana celular para la GH; es el receptor de la hormona de crecimiento (GHR) quien regula su función (7,8) y la actividad de la PRL está regulada por el receptor de la prolactina (PRLR), que participa en numerosas actividades endocrinas (9,10,11) y es un miembro de la familia de las citoquinas receptoras de superficie de membrana celular (12,13).
En el bovino, el GHR está codificado por un único gen que se localiza en el cromosoma 20 (14). Dicho gen está constituido por nueve exones (enumerados del 2-10), siendo la parte que se traduce en proteína y una extensa región no codificante en la región 5´, la que origina diferentes variantes de mRNA (15). El gen que codifica para PRLR también se ubica en el cromosoma 20 en el bovino, a una distancia aproximada de 7Mb del GHR (16) y se describe con un total de 10 exones, de los cuales del 3 al 10 son los exones codificantes (17).
En el gen que codifica para GHR se ha prestado especial atención al polimorfismo F276Y por encontrarse evidencias de alto grado de influencia en el por ciento de proteína y grasa (8) y en el rendimiento en la producción de leche (18,19,20). Por otra parte, el polimorfismo S18N del gen que codifica para PRLR se asocia al rendimiento de la producción láctea y sus componentes (8,21,22).
Para la identificación de estos polimorfismos genéticos se utilizaron metodologías como el alelo discriminante y la extensión de oligonucleótidos en el estudio realizado por Viitala et al. (8) y la secuenciación en los estudios de Blott et al. (18) y Waters et al. (19). El uso de estas metodologías precisa de un equipamiento costoso que encarece la técnica y disminuye las posibilidades de procesamiento de un el elevado número de muestras. Por ello se han realizado investigaciones donde se hace uso de la reacción en cadena de la polimerasa (polymerase chain reaction, PCR) combinada con polimorfismo de longitud de los fragmentos de restricción (restriction fragment length polymorphism, RFLP) en estudios de polimorfismos del locus GHR (23,24).
En tal sentido, se propone el empleo de la metodología amplificación creando el sitio de restricción (amplification created restriction sites ACRS) para identificar el polimorfismo F276Y en el locus GHR y las metodologías PCR-RFLP en la identificación de las variantes polimórficas S18N del locus PRLR. Es por ello que el objetivo del presente trabajo fue describir dos nuevas metodologías para identificar los polimorfismos F276Y del locus GHR y S18N del locus PRLR, así como determinar las frecuencias génicas para ambos loci en la raza Siboney de Cuba. Dichas metodologías podrán ser utilizadas en las evaluaciones genéticas del ganado bovino en general, tomando las regiones estudiadas aquí como genes candidatos a ser incluidos en los programas de selección genética, con particular importancia en la implementación de la selección asistida por marcadores dirigida a la mejora de la calidad de la leche.
MATERIALES Y MÉTODOS
Material genético
Se incluyeron en el estudio un total de 130 vacas de la raza Siboney de Cuba, a las que se les extrajo sangre periférica, con pleno cumplimiento de las normas del comité de ética y bienestar animal. Se emplearon 0.5mL de EDTA 0.5M como anticoagulante y se procedió a la extracción y purificación del ADN mediante el método de precipitación salina descrito por Miller et al. (25). Se determinaron las concentraciones de ADN por espectrofotometría, con lecturas de absorbancia a 260nm y se procedió a realizar las correspondientes diluciones para trabajar con muestras con una concentración de 20pmol/µL.
Genotipado de los receptores de GH y PRL
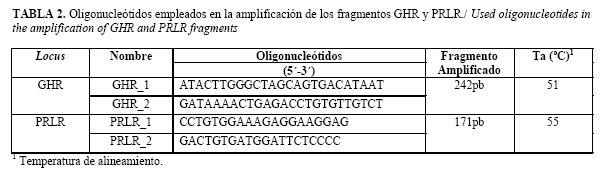
Para el estudio de los polimorfismos que se muestran en la Tabla 1, se desarrolló la metodología de amplificación con creación de sitio de restricción en el locus GHR, donde se colocó el nucleótido A en la posición 23 del GHR_1 en el extremo 3´ y la metodología de polimorfismo de longitud de los fragmentos de restricción para el caso PRLR.
Los oligonucleótidos empleados en la reacción en cadena de la polimerasa se diseñaron con el empleo del programa Vector NTI AdvanceTM 11.0 (Invitrogen Corporation 2008) (Tabla 2). El volumen final de la reacción fue de 25µL constituida por buffer de reacción 10X (conteniendo 10mM TrisHCl, pH 9.0; 50mM KCl y 0.1% Triton X-100), 50mM MgCl2, 1.25mM dNTPs, 20nmol/µL de oligonucleótido correspondiente y 5U de ADN polimerasa comercial (Fermentas), con 50ng/µL de ADN molde completando el volumen final con agua. El programa de amplificación empleado para ambos locus fue similar, solo varió la temperatura de alineamiento. El mismo constó de un ciclo de 94oC por 5min en la etapa de desnaturalización, seguido por 32 ciclos de 94oC por 1min, 1min de alineamiento y 72oC por 1min, el tiempo final de extensión fue de 5min a 72oC. Se utilizó un termociclador Mastercycler Eppendorf® de 96 pocillos.
Después de proceder a chequear la amplificación por electroforesis en gel de agarosa 2%, utilizando Blue Green Loading Dye I (LGC Biotecnologia) como agente intercalante, se tomaron 10µL del producto amplificado y se procedió a la digestión con las enzimas específicas de la casa comercial Promega, para cada uno de los fragmentos las cuales se muestran en la Tabla 3. La temperatura de incubación fue de 37oC durante 3 horas. Para la comprobación de la digestión se se realizó en gel electroforesis de agarosa al 2%, TBE 0.5X, con tinción de Blue Green Loading Dye I (LGC Biotecnología), empleando 0.3µL por cada 5µL de producto digerido, comparado con un marcador de peso molecular de 100pb (Promega) y observado bajo luz UV en transiluminador (CONSORT).

Procesamiento estadístico
El programa GENEPOP versión 3.4 (26,27) fue empleado para realizar el cálculo de las frecuencias alélicas, genotípicas y la prueba exacta de la desviación de los locus del estado de equilibrio Hardy-Weinberg. El cálculo de la heterocigosidad observada (Ho) y esperada (He) por locus se realizó usando el programa ARLEQUIN (28).
RESULTADOS Y DISCUSIÓN
A partir del empleo de los oligonucleótidos diseñados se obtuvo un fragmento amplificado de 242pb para el locus GHR, permitiéndose la creación de un sitio de restricción reconocido por la enzima SspI que posibilitó identificar las dos variantes alélicas relacionadas con el polimorfismo proteico F279Y (Figura 1). En el locus PRLR, el fragmento amplificado fue de 171pb, donde se incluyó el sitio de restricción reconocido por la enzima DdeI lo que permitió identificar las variantes alélicas relacionadas con el polimorfismo proteico S18N en dicho locus (Figura 2).


El alelo F del locus GHR tuvo una mayor frecuencia, con valores de 0.746 (Tabla 4). El comportamiento de la frecuencia de este alelo es similar en otras razas estudiadas. Por ejemplo en siete razas estudiadas por Fontanesi et al. (23) (Holstein Friesian Italiano, Brown Italiano, Simmental Italiano, Jersey, Rendena, Reggiana y Modenese) se aprecian valores de frecuencias del alelos F que van desde 0.772 a 0.947. Igualmente en los seis grupos de datos estudiados por Blott et al. (18) se aprecia el alelo F con una mayor frecuencia, con valores que van desde 0.825 a 0.94. En cambio, los estudios realizados en Holstein Chino mostraron frecuencias del alelo F inferiores al alelo Y con valores de 0.366 y 0.634, respectivamente (24).
En el locus PRLR el alelo S (Tabla 4) fue el de mayor frecuencia. Los estudios realizados por Lü et al. (29) informan frecuencias del alelo S que van desde 0.596 a 0.802 en las cinco razas nativas Chinas, por lo que la frecuencia encontrada en el Siboney de Cuba se encuentra próxima a estas razas. En el Holstein Chino también se describe el alelo S con una mayor frecuencia, pero su valor está más cercano a la frecuencia del alelo N por el gran número de animales homocigotos para el mismo (22).
Se observaron los tres genotipos en ambos loci. En el caso del locus GHR se han descrito los tres genotipos en la mayoría de las razas estudiadas. Tal es el caso de los seis grupos estudiados por Blott et al. (18), la población Holstein estudiada por Li-Juan et al. (24) y las siete poblaciones estudiadas por Fontanesi et al. (23). En tres de ellas no se observó el genotipo YY (Brown Italiano, Jersey y Modenese). En la mayoría de las razas estudiadas el genotipo con una mayor frecuencia fue FF (18,23), con excepción del comportamiento de este locus en el Holstein Chino donde el genotipo más frecuente fue el YY (24). La heterocigosidad observada es superior a la esperada en el Siboney, observándose un desvío del estado de equilibrio Hardy-Weinberg (p<0.05). Similar estado de desviación del estado de equilibrio Hardy-Weinberg se describe en la raza Holstein Chino (24), mientras que para las siete razas estudiadas por Fontanesi et al. (13) no se observa en ninguna de ellas un desvío significativo de dicho estado de equilibrio.
El genotipo SS es el de mayor frecuencia encontrado en el locus PRLR para el Siboney. De forma similar se comporta la raza estudiada por Zhang et al. (22), donde la heterocigosidad observada es menor que la esperada con una desviación significativa de la condición de equilibrio Hardy-Weinberg (p<0.05). En las razas estudiadas por Lü et al. (29) se observó en una de ellas (Qinchuan) una desviación muy significativa del estado de equilibrio Hardy-Weinberg para este locus (p<0.01). En el Holstein Chino también se observó este locus fuera del estado de equilibrio Hardy-Weinberg (22).
Las metodologías desarrolladas sirven de base para la utilización en los protocolos de tipificación para la determinación de la estructura genética de poblaciones bovinas, específicamente para los dos loci estudiados y a su vez, posibilitan la inclusión de ambos receptores como indicadores de selección en los programas de mejoramiento genético, específicamente dirigidos a la mejora de la calidad de la leche en bovinos.
CONCLUSIONES
Se dispone de dos metodologías que permiten identificar los polimorfismos F276Y del locus GHR y S18N del locus PRLR, cuya información pudiese ser utilizada en programas de selección asistida por marcadores, las cuales permitieron identificar las dos variantes alélicas en los loci estudiados y a su vez determinar que ambos loci se encuentran desviados de la condición de equilibrio Hardy-Weinberg.
AGRADECIMIENTOS
Los autores quieren agradecer el soporte financiero brindado por el convenio CAPES/MES (Brasil/Cuba), Brasil para la realización de esta investigación. También agradecemos las facilidades brindadas por la Dirección Nacional de Genética y la Empresa Pecuaria Genética Valles de Perú para la obtención de muestras del rebaño Siboney de Cuba.
REFERENCIAS
1. Etherton TD. Somatotropic function: The somatomedin hypothesis revisited. J Anim Sci. 2004;82:239-244.
2. Etherton TD, Bauman DE. Biology of somatotropin in growth and lactation of domestic animals. Physiol Reviews. 1998;78:745-761.
3. Clevenger CV, Furth PA, Hankinson SE, Schuler LA. The role of prolactin in mammary carcinoma. Endocr Rev. 2003;24:1-27.
4. Kelly PA, Bachelot A, Kedzia C, Hennighausen L, Ormandy CJ, Kopchick JJ, et al. The role of prolactin and growth hormone in mammary gland development. Molec Cell Endocrin. 2002;197(1-2):127-131.
5. Auchtung T. Effects of photoperiod during the dry period on prolactin, prolactin receptor, and milk production of dairy cows. J Dairy Sci. 2005;88(1):121-127.
6. Lee SA, Lee SA, Haiman CA, Burtt NP, Pooler LC, Cheng I, et al. A comprehensive analysis of common genetic variation in prolactin (PRL) and PRL receptor (PRLR) genes in relation to plasma prolactin levels and breast cancer risk: the Multiethnic Cohort. BMC Medical Genetics. 2007;8:72.
7. Kopchick JJ, Andry JM. Growth hormone (GH), GH receptor, and signal transduction. Molec Genetics Metab. 2000;71:293-314.
8. Viitala S, Szyda J, Blott S, Schulman N, Lidauer M, Maki-Tanila A, et al. The Role of the Bovine Growth Hormone Receptor and Prolactin Receptor Genes in Milk, Fat and Protein Production in Finnish Ayrshire. Dairy Cattle Genetics. 2006;173:2151-2164.
9. Kelly PA, Binart N, Lucas B, Bouchard B, Goffin V. Implications of multiple phenotypes observed in prolactin receptor knockout mice. Frontiers neuroendocrinol. 2001;22(2):140-145.
10.Kelly PA, Binart N, Freemark M, Lucas B, Goffin V, Bouchard B. Prolactin receptor signal transduction pathways and actions determined in prolactin receptor knockout mice. Biochem So Transac. 2001;29(2):48-51.
11.Vaclavicek A, Hemminki K, Bartram CR, Wagner K, Wappenschmidt B, Meindl A, et al. Association of prolactin and its receptor gene regions with familial breast cancer. J Clinic Endocrin & Metab. 2006;91(4):1513.
12.Bazan JF. A novel family of growth factor receptors: a common binding domain in the growthhormone, prolactin, erythropoietin and IL-6 receptors, and the p75 IL-2 receptor beta-chain. Biochem Biophys Res Commun. 1989;164:788.
13.Fleenor D, Arumugam R, Freemark M. Growth hormone and prolactin receptors in adipogenesis: STAT-5 activation, suppressors of cytokine signaling, and regulation of insulin-like growth factor I. Hormone Res Pediatrics. 2006;66(3):101-110.
14.Moody DE, Pomp D, Barendse W, Womack JE. Assignment of the growth hormone receptor gene to bovine chromosome 20 using linkage analysis and somatic cell mapping. Anim Genet. 1995;26:341-343.
15.Jiang H, Lucy MC. Variants of the 5'-untranslated region of the bovine growth hormone receptor mRNA: isolation, expression and effects on translational efficiency. Gene. 2001;265:45-53.
16.Turner LB, Harrison BE, Bunch RJ, Porto Neto LR, Li Y, Barendse W. A genome-wide association study of tick burden and milk composition in cattle. Animal Prod Sci. 2010;50(4):235-245.
17.Lü A, Hu X, Chen H, Dong Y, Zhang Y, Wang X. Novel SNPs of the Bovine PRLR Gene Associated with Milk Production Traits. Biochem Genetics. 2011;29(3-4):177-189.
18.Blott S, Kim JJ, Moisio S, Schmidt-Kuntzel A, Cornet A, Berzi P, et al. Molecular dissection of a quantitative trait locus: a phenylalanine-to-tyrosine substitution in the transmembrane domain of the bovine growth hormone receptor is associated with a major effect on milk yield and composition. Genetics. 2003;163(1):253-266.
19.Waters SM, McCabe MS, Howard DJ, Gibson L, Magee DA, MacHughn DE, et al. Associations between newly discovered polymorphisms in the Bos taurusgrowth hormone receptor gene and performance traits in Holstein-Friesian dairy cattle. Animal Genetics. 2011;42(1):39-44.
20.Rahmatalla SA, Müller U, Strucken EM,Reissmann M, Brockmann GA. The F279Y polymorphism of the GHR gene and its relation to milk production and somatic cell score in German Holstein dairy cattle. J Appl Genetics. 2011;52(4):459-465.
21.Scotti E, Fontanesi L, Russo V. Mutations in the bovine prolactin receptor (PRLR) gene: allele and haplotype frequencies in the Reggiana cattle breed. Italian J Anim Sci. 2007;6(1S):207.
22.Zhang J, Zan L, Fang P, Zhang F, Shen G, Tian W. Genetic variation of PRLR gene and association with milk performance traits in dairy cattle. Canadian J Anim Sci. 2008;88(1):33-39.
23.Fontanesi L, Scotti E, Tazzoli M, Beretti F, Dall´Olio S, Davoli R, et al. Investigation of allele frequencies of the growth hormone receptor (GHR) F279Y mutation in dairy and dual purpose cattle breeds. Italian J Anim Sci. 2007;6(4):415-420.
24.Li-Juan W, Qiu-Ling L, Quang-fa W, Hong-Mei W, Jian-Bin L, Yun-Dong G, et al. CRS-PCR polymorphisms of the GHR gene and its relationship with milk production traits in Chinese Holstein cows. Chinese J Agricult Biotechnol. 2009;6(03):215-219.
25.Miller SA, Dykes DD, Polesky HF. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acid Res. 1988;16:12-15.
26.Raymond M, Rousset F. Genepop (Version-1.2) - Population-Genetics Software for Exact Tests and Ecumenicism. J Heredity. 1995;86(3):248-249.
27.Rousset F. Genepop'007: a complete reimplementation of the Genepop software for Windows and Linux. Mol Ecol Resources. 2008;8:103-106.
28.Excoffier L, Laval G, Schneider A. Arlequin ver. 3.0: An integrated software package for population genetics data analysis. Evolutionary Bioinformatics. Online. 2005;1:47-50.
29.Lü A, Hu X, Chen H, Dong Y, Pang Y. Single nucleotide polymorphisms of the prolactin receptor (PRLR) gene and its association with growth traits in chinese cattle. Molec Biol Report. 2011;38(1):261-266.
Recibido: 23-5-2012.
Aceptado: 25-6-2012.

{kind=link}
{kind=link}
{kind=link}