










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Salud Animal
versión impresa ISSN 0253-570X
Rev Salud Anim. vol.34 no.2 La Habana mayo-ago. 2012
COMUNICACIÓN CORTA
Aislamiento de virus influenza porcina en Cuba
Isolation of swine influenza virus in Cuba
A. Vega, Carmen L. Perera, Heidy Díaz de Arce, María T. Frías, L.J. Pérez
Departamento de Virología, Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, Mayabeque, Cuba. Correo electrónico: armando@censa.edu.cu
RESUMEN
Con el fin de realizar el aislamiento del virus de la influenza, porcina se seleccionaron 10 muestras de tejido pulmonar de un total de 49 que resultaron positivas por RT-PCR en tiempo real. El aislamiento se realizó en embriones de pollo con resultados positivos en 4 de las 10 muestras inoculadas. De estas cuatro muestras tres fueron identificadas como virus de influenza pandémicos (H1N1). Los resultados de este trabajo constituyen el primer reporte de aislamiento del virus de influenza porcina en las poblaciones de cerdos de Cuba.
Palabras clave: Influenza porcina, aislamiento, embrión de pollo.
ABSTRACT
In order to accomplish swine influenza virus isolation, 10 tissue lung samples were selected from 49 real time RT-PCR positive samples. Lung homogenate sample were inoculated into 10 day old chicken eggs. Four viral isolates were obtained from the 10 samples selected. Three swine influenza virus isolates obtained were subtyped as pandemic influenza A virus H1N1. This study is the first report of swine influenza isolation in Cuban swine farms.Key words: Swine Influeza, isolation, embrionated chicken eggs.
Las infecciones producidas por virus Influenza A han sido documentadas en aves, cerdos, gatos, perros, hurón, zorros, caballos y mamíferos marinos (1,2). Estos están clasificados dentro de la familia Orthomyxoviridae, son virus envueltos que contienen 8 segmentos de ARN de cadena simple y polaridad negativa que codifican 11 proteínas, algunas de las cuales constituyen elementos estructurales del virión y otras intervienen en la replicación viral (3). Se reconocen 16 subtipos diferentes de hemoaglutininas y 9 de neuroaminidasa las que pueden ser diferenciadas desde el punto de vista antigénico (4).
En el cerdo, circulan, a nivel mundial, tres subtipos principales de influenza A; H1N1, H1N2, y H3N2 (5) y se consideran uno de los patógenos respiratorios más comunes de ahí su importancia en el complejo respiratorio. La infección es caracterizada por fiebre alta, letargia, descarga nasal, tos, disnea y pérdidas de peso aunque también se han reportado brotes de aborto (6,7,8).
La importancia de la infección del cerdo con virus influenza no solo radica en las graves pérdidas económicas que produce en la industria porcina mundial (9) sino también se ha considerado que actúa como un mezclador para la pseudo-recombinación de virus influenza de aves, cerdos y mamíferos, por lo que puede resultar importante en el surgimiento de nuevas cepas de virus con capacidad para producir una pandemia en el humano (10,11).
En nuestro país, a pesar de tener evidencias serológicas de la circulación de virus de la influenza porcina H1 (12) no se habían realizado estudios para obtener el aislamiento de este virus. Por lo que el objetivo de este trabajo fue obtener el aislamiento de virus influenza porcina a partir de muestras que resultaron positivas por RT-PCR en tiempo real en un estudio realizado en paralelo (13), en poblaciones porcinas que presentaron problemas respiratorios.
Durante el año 2010 nuestro laboratorio recibió un total de 238 muestras (157 exudados nasales y 81 provenientes de tejidos pulmonar, tomadas de cerdos con problemas respiratorios las que fueron evaluadas por la técnica de RT-PCR en tiempo real (rRT-PCR) para detectar la presencia de Influenza porcina. Estas muestras se colectaron como parte de un proyecto de la Organización de las Naciones Unidas para la Alimentación y la Agricultura (FAO) coordinado por la Organización Mundial de Sanidad Animal (OIE) para aumentar la vigilancia de la IP a partir de la emergencia de la nueva cepa H1N1/2009 de carácter pandémico.
El procesamiento de las muestras para el diagnóstico por rRT-PCR se realizó según el algoritmo diagnóstico para IP propuesto por OFFLU (14), brevemente: las muestras de tejido se homogenizaron con 1 mL de medio de cultivo DMEM por medio de perlas de cerámica en el equipo MagNa Lyser (Roche Applied Science) a razón de 5000g durante 30s, luego se centrifugaron a 9500g durante 1min y finalmente, se colectó el sobrenadante para la extracción del ARN viral.
El ARN viral se extrajo por medio del kit comercial QIAamp Viral RNA Mini Kit (Qiagen GmbH) según las instrucciones del fabricante. La reacción de RT-PCR en tiempo real se realizó en un solo paso en un volumen total de 20 µL que contenían 10µL de la mezcla de reacción QuantiTectTM Probe RT-PCR master mix (Qiagen), 1 µL de cada un los cebadores (10 µM) y sonda (4 µM), 5 µL del RNA extraído y 2 µL de agua libre de nucleadas (Promega, Madison, USA). Para la tipificación de influenza tipo A la pareja de cebadores y sondas fueron M+25 5´-AGATGAGTCTTCTAACCGAGGTCG-3´,M-124-SIV- 5´-CAGAGACTGGAAAGTGTCTTTGCA-3´, ProbeM+64- 5´- FAM-TCAGGCCCCCTC AAAGCCGA-TAMRA-3´, respectivamente. Para la discriminación del virus pandémico se emplearon los cebadores y sondas N1-F120-5´-CAACACCAACTTTGCTGC-3´; N1-R330-5´-GGAACCGATTCTTACACTGTTGTC-3´, ProbeN1-F2325´-FAM-CAGTCAGTGGTTTCCGTGAAATTAGC BHQ-1-3´, respectivamente.
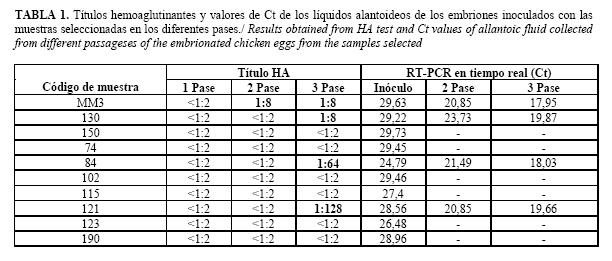
Para el aislamiento viral se seleccionaron un total de 10 muestras de tejido pulmonar de aquellas que resultaron positivas por rRT-PCR y se encontraban almacenadas a -80°C por un período de 6 meses. Los criterios de selección de las muestras para el aislamiento viral fueron: i) el área geográfica y ii) aquellas muestras positivas con un valor del ciclo umbral (CT-de inglés Cycle Threshold) por debajo de 30 (13). A partir de cada muestra se preparó una suspensión al 10% (p/v) en medio de cultivo DMEM y se inocularon en embriones de pollo comerciales a razón de 0,2 mL por embrión y 5 embriones por muestra. La incubación se realizó a 37°C con observación diaria durante un período de 3 días. Los embriones que murieron a las 24 horas post inoculación fueron eliminados y los líquidos alantoideos de los que murieron a partir de las 48 horas o los que permanecieron vivos fueron evaluados por hemoaglutinación rápida (HAR) en placa utilizando una suspensión de eritrocitos de pollo al 10% y por rRT-PCR. En los casos que se obtuvo HAR positiva las colectas fueron tituladas por la técnica de hemoaglutinación según se establece en el manual de la OIE (15).
La identificación de los aislamientos se realizó por rRT-PCR y por el protocolo de secuenciación propuesto por la Organización mundial de la Salud (World Health Organization, WHO) (16).
Brevemente, los productos de PCR purificados se secuenciaron bidireccionalmente con cada uno de los cebadores específicos recomendados por la Organización Mundial de la Salud (16) y el estuche comercial Big Dye Terminator v3.1 cycle sequencing Kit (Applied Biosystems), según las indicaciones del fabricante. Se mezclaron 4 µL de Big Dye, 1 µL de cebador 10 µM y 5 µL de los amplicones purificados para un volumen final de 10 µL.
Del total de muestras evaluadas 4 resultaron positivas a partir del segundo pase. Una de las muestras (MM3) produjo mortalidad de un embrión a las 72 horas pi, el cual al ser evaluado por HAR resultó positivo. El resto de las muestras evaluadas (3) fueron positivas solo por rRT-PCR, con una disminución de los valores de Ct entre 20,85 y 23,73 (Tabla 1) que sugiere un aumento en la carga viral (Figura 1). Las mismas no produjeron mortalidad embrionaria y los líquidos alantoideos fueron negativos por HAR. Todos los aislados obtenidos fueron identificados como subtipos H1N1 con número de acceso en la base de datos GenBank HE584748, HE584750, HE578808, HE578810, HE584756 y HE584758.

Los resultados obtenidos en el éxito del aislamiento viral y la evaluación por HAR corresponden con lo reportado por Metreveli et al. (17) quienes al estudiar dos aislados de influenza porcina en Suiza encontraron que ambos fueron negativos por hemaglutinación durante el segundo pase en células MDCK pero sus títulos infectivos oscilaron entre 6,2 y 7.0 TCID50 /mL. Lo que evidencia que los resultados por hemoaglutinación dependen de las concentraciones de virus en las muestras. En este sentido Gopinath et al. (18) reportaron, en un estudio comparativo de la sensibilidad analítica de un rRT-PCR cuantitativo, RT-PCR convencional y hemoaglutinación (HA) en función de dosis infectivas (DI), que solo fue posible obtener títulos hemoaglutinantes o resultados positivos por RT-PCR convencional cuando las concentraciones de virus en la muestras analizadas fueron altas (aproximadamente 107 DI) mientras que el rRT-PCR resultó positivo en un mayor rango (DI entre 107 y 10-1).
Los resultados obtenidos en el presente estudio, así como los resultados de otros grupos de investigación (17,18), evidencian la necesidad del uso de los ensayos de rRT-PCR en el seguimiento de los virus de IP durante el proceso de aislamiento, como técnica cuantitativa y sensible que oriente la investigación en este campo para una disminución de posibles falsos negativos.
Las diferencias encontradas en el número de muestras positivas por aislamiento viral y por rRT-PCR (4 de10) pudiera estar influenciado por el período de congelación de las muestras lo que pudo afectar la viabilidad del virus o motivadas por la sensibilidad que tiene el rRT-PCR al poder detectar tanto fragmentos de genoma como partículas virales no viables.
Pamela et al. (19) encontraron, al estudiar 541 muestras positivas por rRT-PCR, que solo un 25.1% de estas (136) fueron positivas por aislamiento de Influenza aviar, lo que atribuyen a la no adición de crioprotectores antes de la congelación de las muestras. Munster et al. (20) encontraron coincidencias de un 33.5% (332 de 992) entre el aislamiento viral y RT-PCR y señalaron que aún en condiciones ideales de transporte, almacenamiento y procesamiento no de todas las muestras positivas por RT-PCR se puede lograr aislamiento.
Con la emergencia del H1N1/2009 pandémico el mundo confirmó la potencialidad de los virus influenza de intercambiar genomas de diferentes especies y diferentes continentes (21), de ahí que la constante vigilancia, el análisis de secuencias en busca de marcadores de adaptación a hospederos, diseminación y virulencia sean tareas urgentes de la comunidad científica internacional. Estos análisis dependen en gran medida del éxito de obtener aisladas aquellas cepas de virus que circulen en un área determinada en un momento dado, de ahí la relevancia de aumentar los estudios que faciliten este objetivo.
Finalmente en el presente trabajo se obtuvo por primera vez en el país el aislamiento de cuatro virus de influenza porcina a partir de cerdos con signos respiratorios procedentes de poblaciones porcinas del occidente, centro y oriente del país y constituye las bases para la futura caracterización molecular y filogenético de estos virus.
REFERENCIAS
1. Rimmelzwaan GF, Baars M, van Riel D, Bestebroer TM, Fouchier, RA, Osterhaus AD, et al. Influenza A virus (H5N1) infection in cats causes systemic disease with potential novel routes of virus spread within and between hosts. Am J Pathol. 2006;168:176-183.
2. Reperant LA, van Amerongen G, van de Bildt MWG, Rimmelzwaan GF, Dobson AP, Osterhaus AD, et al. Highly pathogenic avian influenza virus (H5N1) infection in red foxes fed infected bird carcasses. Emerg Infect Dis. 2008;14:1835-1841.
3. Palese P, Shaw ML. Orthomyxoviridae: the viruses and their replication. In: Knipe DM, Howley PM (eds.) Fields virology. Williams & Wilkins, Philadelphia; 2007; pp 1647-1689.
4. Fouchier RA, Munster A, Wallensten TM, Bestebroer S, Herfst D, Smith GF, et al. Characterization of a novel influenza virus hemagglutinin subtype (H16) obtained fromblack-headed gulls. J Virol. 2005;79:2814-2822.
5. Jung K, Ha Y, Chae C. Pathogenesis of swine influenza virus subtype H1N2 infection in pigs. J Comp Pathol. 2005;132:179-184.
6. Zhou NN, Senne DA, Landgraf JS, Swenson SL, Erickson G, Rossow K, et al. Genetic reassortment of avian, swine, and human influenza A viruses in American pigs. J Virol. 1999;73:8851-8856.
7. Karasin AI, Olsen CW, Anderson GA. Genetic characterization of an H1N2 influenza virus isolated from a pig in Indiana. J Clin Microbiol. 2000;38:2453-2456.
8. Pascua PN, Song MS, Lee JH, Cho HW, Han JH, Kim JH, et al. Seroprevalence and genetic evolutions of swine influenza viruses under vaccination pressure in Korean swine herds. Virus Res. 2008;138:43-49.
9. Lee CS, Kang BK, Kim HK, Park SJ, Park BK, Jung K, et al. Phylogenetic analysis of swine influenza viruses recently isolated in Korea. Virus Genes. 2008;37:168-176.
10.Marosin S, Gregory V, Cameron K, Bennett M, Valette M, Aymard M, et al. Antigenic and genetic diversity among swine influenza A H1N1 and H1N2 viruses in Europe. J Gen Virol. 2000; 83:735-745.
11.Vincent Amy L, Ma W, Lager Kelly M, Gramer Marie R, Richt Juergen A, Janke Bruce H. Characterization of a newly emerged genetic cluster of H1N1 and H1N2 swine influenza virus in the United States. Virus Genes. 2009;39:176-185.
12.Barreras Maritza, González Rosa, Gutiérrez E. Influenza porcina. Pesquisa serológica en 2 Provincias en Cuba. Rev Salud Anim. 1983;5:433-436.
13.Pérez LJ, Perera CL, Vega A, Frías MT, Roseaux D, Ganges L, Núñez JI, Díaz de Arce H. Isolation and complete genomic characterization of pandemic H1N1/2009 influenza viruses from Cuban swine herds. 2011; (submitted to VETMIC for publication).
14.OFFLU interim guidance on detecting pandemic H1N12009 in pigs: laboratory testing algorithm. Paris, World Organisation for Animal Health-United Nations Food and Agriculture Organization Network of Expertise on Animal Influenza, 2009 (accessed 20 October 2010).Disponible en: http://www.offlu.net/OFFLU%20Site/OFFLU_SIV_Surveillance_Testing_Algorithm.pdf.
15.Office International des epizooties. Terrestrial Manual. Swine Influenza. 2008; Pp 1128-1138.
16.World Health Organization. Global Alert and Response. Sequencing primers and protocol. 2009 May 12, cited 2009 Nov 18. Disponible en: http://www.who.int/csr/resources/publications/swineflu/GenomePrimers_20090512.pdf.
17.Metreveli G, Emmoth Eva, Zohari S, Balint A, Widen F, Muradrasoli S, et al. Comparison of two H1N2 swine influenza A viruses from disease outbreaks in pigs in Sweden during 2009 and 2010. Virus Genes. 2011;42:236-244.
18.Gopinath VP, Gopal Dhinakar Raj, Raja A, Kumanan K, Subbiah Elankumaran. Rapid detection of Newcastle disease virus replication in embryonated chicken eggs using quantitative real time polymerase chain reaction. J Virological Methods. 2011;171:98-101.
19.Pamela JF, Peterson JM, Merendino T, Nelson M, Lupiani Blanca. Comparison of Real-Time Reverse Transcription-PCR and Virus Isolation for Estimating Prevalence of Avian Influenza Virus in Hunter-Harvested Wild Birds at Waterfowl Wintering Grounds Along the Texas MidGulf Coast (2005-2006 Through 2008-2009). Avian Dis. 2010;54:655-659.
20.Munster VJC, Baas P, Lexmond J, Waldenstrom A, Wallensten T, Fransson GF, et al. FouchierSpatial, temporal, and species variation in prevalence of influenza A viruses in wild migratory birds. PLoS Pathog. 2007;3:0630-0638.
21.Capua I, Cattoli G. One Flu for one Health (Letter). Emerg Infec Dis. 2010;16(4):719.
Recibido: 20-1-2012.
Aceptado: 21-6-2012.

{kind=link}