










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Salud Animal
versión On-line ISSN 2224-4700
Rev Salud Anim. vol.39 no.3 La Habana set.-dic. 2017
ARTÍCULO ORIGINAL
Identificación de enterobacterias productoras de betalactamasas de espectro extendido (BLEEs) en instalaciones porcinas de la provincia Matanzas
Identification of enterobacteria producing extended-spectrum beta-lactamases (ESBLs) in pig farms in Matanzas province
Carelia Martha Marrero-Moreno1, Martha Mora-Llanes1, Rosa Elena Hernández-Fillor2, Michel Báez-Arias2, Tania García-Morey1, Ivette Espinosa-Castaño2*
1Centro Provincial de Higiene, Epidemiología y Microbiología, Matanzas, Cuba.
2Centro Nacional de Sanidad Agropecuaria, Apartado 10, San José de las Lajas, Mayabeque, Cuba.
RESUMEN
Las betalactamasas de espectro extendido (BLEE) son enzimas codificadas por diferentes genes, que se localizan en plásmidos y facilitan su propagación entre bacterias Gram negativas. Estas enzimas inactivan el anillo betalactámico de cefalosporinas de amplio espectro y monobactámicos, pero no actúan sobre cefamicinas, carbapenémicos y son inhibidas por ácido clavulánico. El objetivo del presente trabajo fue detectar enterobacterias, productoras de BLEEs, en granjas y mataderos de la provincia Matanzas. Las muestras se colectaron de tres granjas y dos mataderos durante 2016 y 2017. Se realizaron hisopados de las manos de operarios, de superficies ambientales, de instrumentos de trabajo e hisopados rectales de animales aparentemente sanos. Las muestras se cultivaron en medio suplementado con cefotaxima. La susceptibilidad microbiana se determinó por el método de difusión en disco. La detección de BLEE se realizó por tres ensayos fenotípicos: una prueba tamiz con discos indicadores y dos ensayos basados en el efecto sinergia de inhibidores de betalactamasas. También se realizó, mediante PCR, la detección genotípica de fragmentos de genes que codifican para estas enzimas. Se identificaron 41 aislados de enterobacterias; de ellos 17 fueron positivos en la prueba tamiz y 15 coincidieron en los ensayos de sinergia y PCR. La mayor frecuencia fue para el gen blaCTX-M seguido del genblaTEM, mientras que el gen blaSHV no se detectó. Se encontraron nueve aislados BLEE positivos en hisopados rectales, dos en superficies, dos en las manos de un operario y cuatro en instrumentos. Cuatro aislados BLEE positivos, procedentes de exudados rectales, fueron también multirresistentes a quinolonas, tetraciclinas y aminoglucósidos. El hallazgo de aislados productores de BLEE indica que estos animales pueden ser un reservorio de genes de resistencia contenidos en elementos móviles que, a su vez, se pueden propagar a través del contacto directo y en la cadena de producción de alimentos.
Palabras clave: enterobacterias, betalactamasas de espectro extendido, multirresistencia, cerdos, Escherichia coli.
ABSTRACT
Extended-spectrum beta-lactamases (ESBLs) are enzymes encoded by different genes (blaSHV, blaTEM, blaCTX-M), located in plasmids, which facilitate their propagation among Gram-negative bacteria. These enzymes inactivate the beta-lactam ring of broad-spectrum, cephalosporins and monobactamic, but they are sensitive to cefamycins, carbapenems and betalactamase inhibitors. The objective of this work was to detect ESBLs-producing enterobacteria in farms and slaughterhouses in Matanzas province. Samples were collected from three pig farms and two slaughterhouses during the years 2016 and 2017. Swabs were made from different sites including animals, workers and environment; and they were grown in medium supplemented with cefotaxime. The antimicrobial susceptibility was determined by the disk diffusion method and ESBL detection by means of a screen test with indicator disks, two synergy tests with betalactamase inhibitors and by polymerase chain reaction (PCR). Forty-one enterobacteria isolates were identified, 17 of them were positive in the sieve test and 15 coincided in the synergy and PCR assays. The highest frequency was for the blaCTX-M gene followed by the blaTEM gene, while the blaSHV gene was not detected. The positive ESBL isolates were found in rectal swabs (9), on surfaces (2), hands of an operator (2) and on instruments (4), only on farm A and slaughterhouse 1. Four of these isolates also presented multiresistance to quinolones, tetracyclines, and aminoglycosides. This work evidences the dissemination of positive E. coli ESBL isolates in non-clinical settings. These pig production facilities are bacteria reservoirs and resistance genes propitiating their propagation through direct contact, in the food production chain or their dissemination to the environment.
Key words: enterobacteria, extended-spectrum β-lactamases, multidrug resistant, pigs, Escherichia coli.
INTRODUCCIÓN
El incremento de la resistencia bacteriana a los antibióticos constituye un problema actual para controlar las infecciones bacterianas, tanto en medicina humana como en veterinaria (1,2).
La resistencia adquirida es la capacidad de los microorganismos que les permite crecer en presencia de antimicrobianos, a los cuales previamente eran sensibles. Este comportamiento ocurre por adquisición de genes, ya sea por mutaciones o por incorporación de elementos genéticos móviles como plásmidos, tramposones o secuencias de inserción (1,2,3). Este material genético puede, además de ser trasmitido a las siguientes generaciones (transmisión vertical), ser trasferido o captado por otras bacterias físicamente próximas a través de los eventos que comprenden la transmisión horizontal (conjugación, transformación y transducción), tanto entre cepas de la misma especie como de especies filogenéticamente alejadas (4,5).
La familia de las enzimas betalactamasas hidroliza antibióticos betaláctamicos, cefalosporinas de tercera y cuarta generaciones, así como carabapenemes. Estas enzimas se diferencian según el sitio donde se ubican los genes que dan lugar a su expresión, ya sea en cromosomas o en plásmidos y el tipo de expresión (inducibles o constitutivas). Las betalactamasas de espectro extendido (BLEE) y algunas de tipo AmpC son codificadas en plásmidos y constituyen el mecanismo de resistencia de mayor impacto para las bacterias Gram negativas (enterobacterias y bacterias no fermentadoras). Las BLEEs inactivan el anillo betalactámico de cefalosporinas de amplio espectro y monobactámicos. Sin embargo, a diferencias de las AmpC, no actúan sobre las cefamicinas, carbapenémicos y a los inhibidores de betalactamasas (IB) como ácido clavulánico, sulbactam y tazobactam. Las betalactamasas de tipo AmpC no son resistentes a las cefalosporinas de cuarta generación, cefepime, mientras que las BLEE hidrolizan este antibiótico (6,7).
La identificación de aislados productores de BLEEs supone un reto para los laboratorios de Microbiología Clínica, debido a que su detección fenotípica no es fácil por la coexistencia, en ocasiones, de más de un mecanismo como son la modificación de las porinas en la membrana celular y las bombas de eflujo, que también contribuyen a la resistencia. Se recomienda la utilización de métodos fenotípicos y genotípicos, pero la elección de los ensayos depende de las condiciones de cada laboratorio (8,9).
El empleo de los antibióticos con fines terapéuticos o profilácticos, pero a gran escala, en ocasiones sin control y supervisión, propicia la presencia de los antimicrobianos en diferentes ecosistemas (poblaciones humanas, animales y el medio ambiente). Estos compuestos ejercen una presión de selección para microorganismos resistentes patógenos o no, presentes en estos ecosistemas, los cuales se convierten en un reservorio de bacterias y genes resistentes y representan un peligro potencial para la salud, de ahí la necesidad de conocer sobre su presencia (10,11,12).
Las instalaciones de producción animal constituyen un reservorio para bacterias resistentes, que pueden ser difundidas a la población por contacto directo o a través de la cadena alimentaria. En diferentes especies de animales con destino al consumo humano, principalmente en aves, seguida de cerdos, también en animales de compañía y en especies salvajes se evidenció la prevalencia de enterobacterias productoras de BLEE y se sugiere que estos escenarios sean reconocidos como importantes reservorios deestasbacterias (13,14,15).
En Cuba existen datos sobre la presencia de bacterias productoras de BLEE en instalaciones hospitalarias y, con menor medida, en pacientes de la comunidad. Aislados de Klebsiella spp. productoras de BLEE se detectaron en hospitales de diferentes provincias del país (16). En Cuba, según el conocimiento de los autores, no existen datos publicados sobre la presencia de bacterias productoras de BLEE en cerdos. El objetivo del presente trabajo fue determinar la presencia de enterobacterias productoras de betalactamasas de espectro extendido en granjas y mataderos porcinos de la provincia Matanzas.
MATERIALES Y MÉTODOS
Criterios de inclusión
En este estudio se incluyeron personal con riesgo ocupacional, cerdos aparentemente sanos y superficies ambientales de las instalaciones. Se tuvo en cuenta el previo consentimiento informado de todo personal involucrado en la investigación, además de la autorización de los administrativos de la unidad y del servicio veterinario para la recogida de las muestras. Los cerdos eleccionados se mantuvieron en condiciones de manejo y tenencia propias de cada unidad, bajo las normas descritas por el Instituto de Investigaciones Porcinas (17) y se respetaron estrictamente las normas éticas y de bienestar animal referido por la OIE (18). Los animales que se utilizaron para la investigación se consideraron animales aparentemente sanos, a pesar de que no se realizaron exámenes complementarios para corroborar su estado de salud.
Enfoque y tipo de investigación
Se realizó un estudio descriptivo observacional de tipo transversal dirigido a la identificación de enterobacterias productoras de BLEEs. El estudio se realizó en tres granjas tecnificadas de producción porcina (A, B, C) y en dos mataderos (M-1 y M-2), localizados en la provincia Matanzas, Cuba, durante los meses de julio-septiembre de 2016 y mayo de 2017.
Recogida de muestras
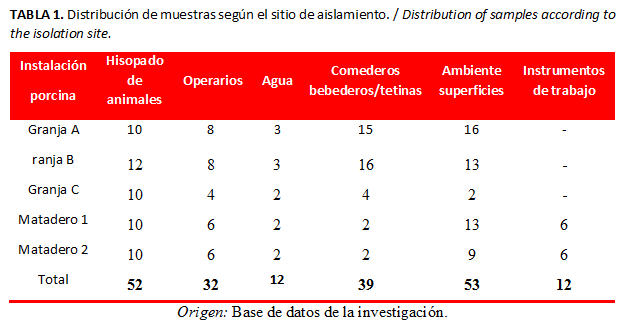
Las muestras se colectaron por personal del laboratorio del Centro Provincial de Higiene, Epidemiología y Microbiología de Matanzas, siguiendo las recomendaciones para recogida, transporte y conservación de muestras (15,22). Se estimó un tamaño de muestra de 200, con una precisión de 5 %, nivel de confianza de 90 % y una prevalencia del 30 % (19). Las 200 muestras se distribuyeron en hisopados rectales de animales, de tetinas/bebederos, comederos, superficies ambientales, operarios e instrumentos de trabajo por cada instalación porcina (granjas y mataderos). Se colectaron muestras de agua (de tanques de alimentación y del pozo que abastece a cada una de las instalaciones) (Tabla 1).
Las muestras se colectaron con hisopos estériles, utilizando un método “no destructivo de arrastre”. Los hisopados rectales se realizaron a partir de animales aparentemente sanos correspondientes a las categorías preceba y ceba final de las granjas; en los mataderos, se les efectuaron a los cerdos horas antes del sacrificio. Las muestras de operarios en cada una de las instalaciones (manos y botas), de superficies (pasamamos, rejas, mesas de trabajo, tetinas/bebederos y comederos) e instrumentos de trabajo se tomaron acorde a Davies et al. (19) y la OIE (20). Todos los hisopados se introdujeron en microtubos de roscas de 2 mL que contenían el medio Caldo Müller–Hilton (Biocen, 4030) suplementado con cefotaxima a 2 µg/mL (15).
Las muestras de agua se tomaron en frascos estériles de 120 mL y se procesaron según la norma cubana 827 del 2012 (21); se utilizó el medio caldo Lactosado (Biocen, 4028) y caldo Bilis Verde Brillante (Biocen, 4029).
Todas las muestras obtenidas de las granjas y mataderos se transportaron en neveras con pastillas refrigeradas hasta los laboratorios y se sembraron en el medio agar Mac Conkey (Biocen, 4014) suplementado con cefotaxima a 2 μg/mL (15). Los cultivos se incubaron en ambiente aerobio a 37ºC de 18 a 24 horas y se realizaron lecturas a las 24 horas de la siembra.
Aislamiento e identificación
Se seleccionaron los cultivos en los que se observaron colonias con características presuntivas de especies de la familia Enterobacteriaceae (colonias rosadas o blancas). Para garantizar la pureza de los aislados se realizó un segundo subcultivo en el mismo medio y se procedió a su identificación mediante el empleo de pruebas bioquímicas convencionales, según Kan et al. (22). La prueba de oxidación/fermentación se utilizó para distinguir bacterias Gram negativas fermentadoras o no; se realizaron cultivos en el medio Hugh and Leifson (Oxoid), se incubaron a 37ºC en condiciones aerobias y no anerobias mediante la adición de parafina líquida durante 18 horas. La confirmación de las especies de enterobacterias se realizó mediante el empleo de los sistemas comerciales API 20 E (BioMérieux, Francia), según las condiciones prescritas por el fabricante.
Conservación de los aislados mediante congelación
A partir de una colonia de cada aislamiento se realizaron subcultivos en 500 μL de medio Caldo cerebro corazón (CCC) (BIOCEN, 4035) y se incubaron a 37ºC durante 18 horas; posteriormente se tomaron 200 μL y se mezclaron con 200 μL de medio CCC suplementado con glicerol al 20 %; se homogenizaron mediante agitación suave y se conservaron a -20ºC.
Procedimiento estándar para la realización de la susceptibilidad in vitro a los antimicrobianos por difusión en Agar con Discos
A partir de los aislados conservados se realizaron cultivos en el medio Agar Base suplementado con sangre ovina al 5 %; se seleccionaron colonias aisladas y se prepararon suspensiones bacterianas en solución salina fisiológica, las cuales se ajustaron con el patrón 0.5 de la escala Mac Farland, equivalente a 1,5 x108ufc/mL. Se realizó la inoculación en placas de agar Müller Hinton (BIOCEN, 4013) con hisopo por toda la superficie y se colocaron manualmente los discos con pinzas estériles a una distancia de 2 cm uno de otro y a 1,5 cm del borde de la placa. Se utilizó la cepa Escherichia coli ATCC 25922 como control de la calidad. Se procedió a la lectura (diámetro del halo incluyendo al disco). La clasificación de la sensibilidad, la sensibilidad intermedia y la resistencia de los aislados se basó en los criterios establecidos por el Instituto de Estándares Clínicos y de Laboratorios (23).
Detección fenotípica de BLEE
La producción de BLEEs se determinó por la combinación de tres metodologías:
- Prueba tamiz: el método consistió en una prueba de difusión en agar con discos, tal y como se describió anteriormente, para la cual se utilizaron los siguientes discos de antibióticos, procedentes de la casa comercial Liofilchem: tres discos de cefalosporinas de tercera generación ceftazidima (CAZ, 30 µg), ceftriazona (CRO, 30 µg), cefotaxima (CXT, 30 µg), un disco de cefalosporina de cuarta generación, cefepime (FEP, 30 µg), un disco de monobactamen aztreonam (ATM, 30 µg) y un disco de la cefamicina, cefoxitin (FOX, 30 µg), una vez colocados los discos las placas se incubaron a 37 ºC por 16 a 20 horas. Se procedió a la lectura (diámetro del halo incluyendo al disco). Se consideró la presencia de aislados potenciales productores de BLEEs cuando se obtuvieron halos de inhibición para los discos CAZ, CRO, CXT, FEP y ATM iguales o inferiores a 17, 19, 22, 18 y 17 mm, respetivamente, y un halo igual o superior a 14 mm para el disco FOX (23).
- Prueba de sinergia de doble disco: la producción de BLEE se determinó por la técnica de sinergismo de doble disco propuesta por Jarlier et al. (15) y siguiendo los procedimientos establecidos por CLSI (23). El método consistió en situar un disco que contiene amoxicilina y el inhibidor de betalactamasa, ácido clavulánico (AUM), en la relación 20/10 µg, próximo a discos de betalactámicos indicadores CXT (30 µg), CAZ (30 µg), FEP (30 µg) y AZM (30 µg) a una distancia lineal de 20 mm del disco central. El criterio de positividad para aislados productores de BLEE fue la observación de una ampliación del halo de inhibición de cefotaxima, ceftazidima, cefepime o aztreonam en la zona próxima al disco que contenía amoxicilina-ácido clavulánico (sinergia) o presencia de una "zona fantasma" (inhibición del crecimiento) entre las cefalosporinas o aztreonam y el inhibidor.
- Prueba de epsilometría (E- test): se realizó para los aislados que resultaron positivos en la prueba tamiz descrita anteriormente; se siguió el procedimiento estándar para la realización de un antibiograma por difusión con discos y se aplicaron tiras comerciales de Etest®, según las directrices del fabricante (Liofilchem). Se emplearon tiras de plástico no poroso de 5 cm de largo y 5 mm de ancho, que contenían ceftazidima (CAZ) en una mitad con una concentración decreciente de 32 µg/mL hasta 0.5 µg/mL; la otra mitad contiene desde 4 µg/mL hasta 0.064 µg/mL con 4 µg/mL de ácido clavulánico en concentración fija. La concentración mínima inhibitoria (CMI) se determinó observando el punto en que la elipse de inhibición interceptó la escala impresa en la tira. Se leyeron el extremo del antibiótico sin el ácido clavulánico y el extremo que contenía ácido clavulánico; se dividieron las dos concentraciones en ese orden. El resultado es positivo para BLEE cuando la proporción es ≥8 y negativo <8.
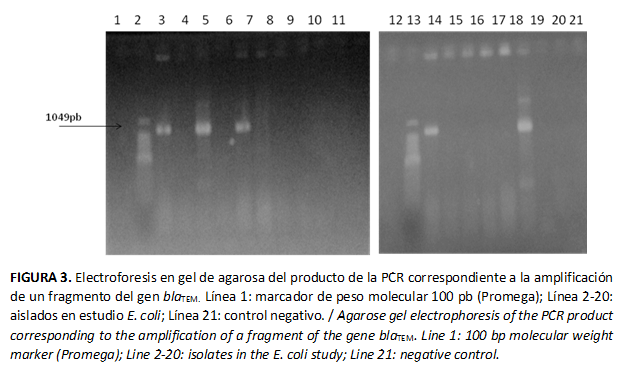
- Ensayos de ácidos nucleicos: los aislados bacterianos positivos en la prueba tamiz se analizaron por la técnica molecular de reacción en cadena de la polimerasa (PCR, de sus siglas en inglés, Polymerase chain reaction) para los genes blaTEM, blaSHV y blaCTX-M relacionados con la producción de BLEE (24, 25). A partir de una colonia se suspendió una asada en 100 μl de agua destilada estéril en tubos de microcentrífuga de 1.5 ml; se sometió a una temperatura de 100 ºC durante 10 minutos. Pasado este tiempo se colocaron en hielo por 20 minutos. Se centrifugó a 5000 g durante 15 minutos y se tomaron 5 μL del sobrenadante (ADN) para agregarlos a la mezcla de PCR. La mezcla de PCR se realizó en un volumen final de 25 μL, conteniendo 1X GoTaq® Green Master Mix (Promega, Madison, EE.UU.) y 20 pM de cada cebador. Las cantidades utilizadas fueron: 12.5 μL de Master Mix (Promega), 1.5 µL de cada cebador y 7.5 µL Agua MilliQ estéril. Las condiciones de la reacción y las secuencias de los cebadores aparecen en la Tabla 2. Los cebadores se sintetizaron en el Centro de Ingeniería Genética y Biotecnología (CIGB), Cuba. Los ensayos de PCR se realizaron en un termociclador Mastercycler.
Visualización de los productos de PCR
Los productos de PCR se visualizaron por electroforesis en gel de agarosa al 2 % en TBE 0.5X con Bromuro de Etidium (1 mgmL-1) en un transiluminador de luz ultravioleta (Pharmacia Lkb). Se utilizó un marcador de peso molecular de 100 pb (Promega). Cada amplificación incluyó un control negativo, en el cual se adicionó agua.
Se evaluó la susceptibilidad antimicrobiana utilizando los discos de otras familias de fármacos ciprofloxacina (5 µg), tetraciclina (30 µg), gentamicina (10 µg), azitromicina (15 µg) para todos los aislados que resultaron positivos por las pruebas anteriores.
Análisis estadístico
Los resultados se organizaron en una base de datos que comprendió el lugar de procedencia, el sitio de aislamiento, la identificación del microorganismo (género y especie), el perfil de susceptibilidad antimicrobiana, la detección fenotípica de BLEE por los métodos de cribado, así como su confirmación por los métodos moleculares realizados.
El análisis de los resultados se realizó mediante estadística descriptiva y cálculo de las frecuencias relativas en el programa estadístico Epidat Versión 3.1 (26). Se compararon los porcentajes y se ordenaron los datos obtenidos en tabla de contingencia con el fin de facilitar la interpretación de la información.
RESULTADOS Y DISCUSIÓN
De 200 muestras colectadas en las instalaciones porcinas, se obtuvieron 41 aislados de bacterias Gram negativas, correspondientes a la familia Enterobacteriaceae resistentes a la concentración 2 µg/mL de cefotaxima. Según las características culturales, morfológicas y pruebas bioquímicas se identificaron los siguientes géneros y especies: 36 aislados correspondieron a Escherichia coli, dos a Serratia marcescens, dos a Enterobacter cloacae y un aislado a Enterobacter sakazakii.
E. coli fue la especie más frecuente encontrada entre los aislados resistentes a cefotaxima. Este microorganismo es la enterobacteria resistente a betalactámicos que se aísla con más frecuencia en los últimos años en los seres humanos y en los animales de granja, especialmente en pollos y cerdos, así como en diversos alimentos. Es una bacteria comensal en los seres humanos y los animales, se encuentra presente en el medio ambiente y se considera un indicador de la contaminación fecal en los alimentos y el agua. E. coli es la principal causa de infección del tracto urinario (ITU), tanto en pacientes hospitalizados como en las infecciones que se manifiestan en la comunidad, donde se aísla en un porcentaje aún mayor. El flujo de plásmidos con genes de resistencia de cepas comensales de E. coli a patógenas constituye un problema de salud, pues reduce las opciones terapéuticas (6,27).
Para el aislamiento de bacterias Gram negativas productoras de BLEE, un aspecto de interés resulta la clase de cefalosporina y la concentración que se adiciona al medio de cultivo. García et al. (28) plantean que para identificar bacterias productoras de BLEE a partir de hospederos portadores se pueden utilizar los medios suplementados con cefotaxima o ceftazidima a una concentración de 1µg/L; sin embargo, otros estudios publicados (29) refieren concentraciones de cefotaxima que oscilan entre 0,5 y 4 µg/mL. Otros autores refieren que si se utiliza una concentración superior a 4 µg/mL, es decir, demasiado alta, entonces disminuye la sensibilidad en la detección (30). Es conveniente que para la elección de la concentración del agente antimicrobiano, se considere la situación epidemiológica de la unidad o centro donde se lleve a cabo el estudio (31). Teniendo en cuenta que en Cuba no existen antecedentes sobre la presencia de aislados productores de BLEE en instalaciones de producción animal, se utilizó una concentración equivalente a 2 µg/mL de cefotaxima, superior a la CMI para favorecer la sensibilidad.
En la prueba tamiz, 17/41 aislados mostraron halos iguales o inferiores a 17, 22 y 29 mm para ceftazidima, ceftriaxona y cefotaxima, respectivamente, tres cefalosporinas de tercera generación (C3G). Estos aislados fueron resistentes al aztreonam y cefepime con halos menores de 17 y 18 mm, mientras que mostraron sensibilidad a cefoxitin con halos superiores a 14 mm (Figura 1). Según estos criterios, los aislados manifestaron propiedades descritas para cepas productoras de BLEE (10,11).
El empleo de discos "indicadores" en la prueba tamiz reveló características específicas de las BLEE y permitió discriminar sobre otros mecanismos de resistencia. El uso del disco de cefamicina (cefoxitina, FOX) descartó a los aislados, en los que la resistencia se debió a deficiencias en las porinas (proteínas de la membrana celular). Los aislados BLEE positivos fueron sensibles a este compuesto, sin embargo, cuando la resistencia se debió a deficiencias en las porinas, la cefoxitina no penetró a las células; en tal caso los aislados se manifestaron resistentes (30,31) y no se consideraron como posibles productores de BLEE.
El uso de la cefalosporina de cuarta generación (cefepime) también contribuyó a descartar otro mecanismo de resistencia; en este caso la presencia de betalactamasas no inducibles, denominadas AmpC, ya que este antibiótico no es hidrolizado por esta betalactamasa, mientras lo es por la BLEE. La identificación de AmpC es de importancia, pues los genes que codifican para estas enzimas también pueden estar localizados en plásmidos, elementos transmisibles que contribuyen a su propagación, pero requieren de metodologías específicas para su identificación y no se consideraron en este trabajo; además, las AmpC aún presentan baja frecuencia en aislados de origen animal (32,33).
Con la finalidad de detectar BLEE se emplean inhibidores de estas enzimas, usualmente el ácido clavulánico en combinación con cefalosporinas de tercera generación. Este compuesto inhibe a las BLEE y reduce el nivel de resistencia de estas a las cefalosporinas. Un total de 17 aislados mostraron la sensibilidad frente a inhibidores, específicamente a amoxicilina/ácido clavulánico, el cual indujo la aparición de halos fantasmas o a la ampliación del halo de inhibición para las cefalosporinas en la zona cercana al disco que contenía esta mezcla (Figura 2).
A través de la prueba de sinergia Epsilon, que contenía el antibiótico ceftazidima, se confirmó la producción de BLEE en 15/17 aislados que fueron previamente positivos por las pruebas de tamizaje y sinergia descritas anteriormente; la proporción a partir de los valores de CMI con y sin inhibidor resultó ≥8 para estos aislados. Sin embargo, 2/17 aislados no presentaron el patrón fenotípico esperado para la producción de BLEE; es posible que las enzimas producidas no presenten actividad sobre la ceftazidima. Las BLEE del grupo CTX-M hidrolizan mejor la cefotaxima (CTX) y la ceftriaxona (CRO) que la ceftazidima (CAZ), incluso el grado en que son inhibidas por inhibidores también es variable, esta puede ser superior por tazobactam que por ácido clavulánico (34,35).
La recomendación es utilizar varias cefalosporinas simultáneamente en el antibiograma, aunque algunos cambios en la secuencia favorecieron la hidrólisis sobre ceftazidima, específicamente en los aminoácidos que se encuentran en las posiciones 240 y 267 y desempeñan un papel importante en la evolución de estas betalactamasas (32,33,34). Los sistemas automáticos para la determinación de la sensibilidad a los antimicrobianos consideraron esta experiencia e incluyeron simultáneamente cefotaxima y ceftazidima con ácido clavulánico (34,35,36).
La producción de BLEE es uno de los mecanismos de resistencia más difíciles de detectar en los laboratorios de Microbiología clínica por métodos fenotípicos. La selección depende de las condiciones del laboratorio. La prueba tamiz y de sinergia son de fácil realización e interpretación y no requiere disponer de equipamiento. En general, la presencia de mecanismos mixtos de resistencia a antibióticos betalactámicos (BLEE y/o AmpC) dificultan en gran medida las técnicas de confirmación fenotípica a nivel de clase. Por lo que para obtener resultados confirmatorios con mayor rapidez, se aconseja la utilización de técnicas moleculares, específicamente la PCR (34,35).
El gen blaTEM se detectó en 5/17 aislados (Figura 3) y el gen blaCTX-M en 15/17, mientras el gen blaSHV no se detectó. Los aislados positivos para el gen blaTEM también lo fueron para blaCTX-M. La ausencia del gen blaSHV coincide con los informes de Lili et al. (15) en el estudio de E. coli procedente de cerdos; mientras que la detección simultánea en un mismo aislado de los genes blaTEM y blaCTX-M concuerda con otros trabajos (37,38,39). El fragmento del gen blaCTX-M se encontró con mayor frecuencia en hisopados rectales de cerdos y en superficies ambientales.
Fueron negativos por PCR dos aislados negativos en la prueba de E-test–cefazidima. La familia de las enzimas betalactamasas de tipo CTX-M es muy amplia y existen numerosas secuencias de cebadores descritas para la amplificación de fragmentos de los genes para su expresión (24,25); sin embargo, es posible la existencia de cambios en la secuencia nucleotídica en las regiones de hibridación con los cebadores empleados en este trabajo, y esta podría ser la razón para que estos dos aislados, que mostraron un comportamiento fenotípico compatible con BLEE, no amplifiquen fragmentos por PCR.
Los resultados coinciden con la situación epidemiológica que se describe en la actualidad para las enzimas BLEE. La epidemiología y la evolución de las bacterias productoras de BLEE constituyen un proceso dinámico. En un principio, el género Klebsiella en pacientes hospitalizados se aisló con mayor frecuencia asociado a la presencia de BLEE derivadas de TEM y SHV. En los últimos 10 años se incrementa la prevalencia, ahora de E. coli BLEE+, en portadores fecales en pacientes hospitalizados y se extiende a pacientes de la comunidad, además, ahora las BLEE más frecuentes son del tipo CTX-M (40).
La prueba de susceptibilidad para otras familias de antibióticos (ciprofloxacina, tetraciclinas, gentamicina y azitromicina) reveló multirresistencia en 3/17 aislados de hisopados rectales y un aislamiento de la superficie de un pasamanos en la granja A, que fueron BLEE positivos. Estos antibióticos se utilizan con mayor frecuencia que las cefalosporinas en medicina veterinaria (39,40), que tiene una indicación muy restringida; sin embargo, los genes que contribuyen a la resistencia frente a estos compuestos se localizan, en ocasiones, en los mismos elementos móviles que transportan genes para la expresión de BLEE, por lo que el uso en la crianza animal de estos antimicrobianos favorece la coselección de estos marcadores de resistencia en aislados productores de BLEE (41,42).
La presencia de aislados productores de BLEEs en mayor frecuencia, a partir de muestras de hisopados rectales y el hallazgo de aislados también productores de BLEE en manos de operarios, superficies ambientales e instrumentos de trabajo, sugiere la posible diseminación de estos desde el excremento de los cerdos al ambiente circundante (Tabla 3). Un aspecto relevante en las granjas de cerdos es el hacinamiento en el que, con frecuencia, se crían estos animales, lo cual propicia enormemente la dispersión y diseminación de estas bacterias, así como el estrés al que se exponen los animales, la temperatura ambiental y la presencia de vectores (33). La posible transferencia de cepas BLEE+ de animales a humanos por contacto cercano parece posible, pero las evidencias aún son escasas; los estudios basados en herramientas de epidemiología molecular han podido mostrar la relación clonal entre cepas resistentes de origen humano y animal (43-47).
Las bacterias contenidas en reservorios fómites, dispositivos de uso común o superficies ambientales, son subestimadas en cuanto a su capacidad de portar genes de resistencia y su capacidad de trasmisión (48). Las enterobacterias requieren de humedad para su sobrevivencia en superficies inanimadas y muchas especies lo logran por meses. Las manos colonizadas del personal también pueden contribuir a su diseminación, luego del contacto con superficies contaminadas (36, 49). Las medidas de higiene y manejo son necesarias para establecer un equilibrio en las áreas donde se encuentran animales en cría intensiva, una apropiada limpieza y desinfección de las superficies y, sobre todo, el lavado de manos después del contacto con reservorios potencialmente contaminados (50,51).
La cadena alimentaria es un posible vehículo de transferencia de genes de resistencia al hombre, en tal sentido se sugirió un origen alimentario a la presencia de cepas de E. coli portadoras de BLEE en la microbiota intestinal de humanos, y un posible origen animal a las cepas multirresistentes de E. coli implicadas en infecciones urinarias en clínica humana (36,37,43). La presencia de BLEE en los aislados analizados enfatiza la necesidad de profundizar en estudios adicionales sobre las características moleculares relacionadas con este mecanismo de resistencia y confirmar su posible propagación mediante los ensayos de conjugación.
La prevalencia de enterobacterias productoras de BLEEs en la granja A fue de 24,3 % (10/41), en el Matadero M-1 fue de 12,1 % (5/41) y hubo instalaciones donde no se detectaron bacterias productoras de BLEE (granjas B, C y M-2). Una posible explicación a las diferencias encontradas ente las instalaciones investigadas podría ser el consumo de medicamentos en ellas y aspectos de bioseguridad, sin embargo, estos factores no se consideraron en el estudio. Investigaciones realizadas en países europeos demostraron que el uso limitado de antibióticos, la aplicación de estos solamente a animales enfermos en granjas y un alto nivel de bioseguridad se asociaron con una baja prevalencia de enterobacterias productoras de BLEE (52,53). Estos datos enfatizan la necesidad de acompañar los estudios microbiológicos de datos epidemiológicos que revelen las características de cada instalación y la asociación entre estos factores.
Este trabajo evidenció la presencia de aislados de E. coli BLEE positiva en ámbitos no clínicos. Se encontró que la microbiota intestinal actúa como reservorio de estos microorganismos en los animales que pueden ser un vehículo importante para su transferencia al hombre y a otros animales. Es necesario profundizar en el estudio de los factores que contribuyeron a la diseminación de E. coli BLEEs+ para poder controlar este problema de creciente repercusión en la salud humana y animal.
- López-Pueyo MJ, Gaite B, Amaya-Villar R, Garnacho-Montero J. Puesta al día en medicina intensiva: el enfermo crítico con infección grave Multirresistencia antibiótica en unidades de cuidados críticos. Med Intensiva. 2011;35(1):41-53.
- Nguyen TN, Nguyen VC, Thwaites G, Carrique-Mas J. Antimicrobial Usage and Antimicrobial Resistance in Animal Production in Southeast Asia: A Review Antibiotics. 2016;5(37).
- Gang Zhou, Qing-Shan S, Xiao-Mo H, Xiao-Bao X. The Three Bacterial Lines of Defense against Antimicrobial Agents. Int J Mo. Sci. 2015;16:21711-21733.
- Frances M, Ellis RJ, Raymond B. Ecological and genetic determinants of plasmid distribution in Escherichia coli. Environment Microbiol. 2016;18(11):4230-4239
- Madigan MT, Martinko JM, Dunlap PV, Clark DP. Brock: Biología de los microorganismos (12th ed.). Madrid (España): Pearson.http://cienciaybiologia.com/microbiologia/familiaenterobacteriaceae. 2009
- Li XZ, Mehrotra M, Ghimire S, Adewoye L. β-Lactam resistance and β-lactamases in bacteria of animal origin. Vet Microbiol. 2007;121:197-214.
- García ML, Hendriksen RS, Fraile L, Aarestrup FM. Antimicrobial resistance of zoonotic and commensal bacteria in Europe: the missing link between consumption and resistance in veterinary medicine. Vet Microbiol. 2014;170:1-2.
- Bou AG, Chaves SF, Palomo OA, Iglesias OJ. Métodos microbiológicos para la vigilancia del estado de portador de bacterias multirresistentes Nro. 55 de Enfermedades Infecciosas y Microbiología Clínica Recomendaciones de la Sociedad Española de Procedimientos en Microbiología Clínica. 2015; SEIMC.
- Villegas M, Guzman B, Sifuentes O, Rossi F. Increasing prevalence of extended-spectrum-betalactamase among Gram-negative bacilli in Latin America – 2008 update from the Study for Monitoring Antimicrobial Resistance Trends (SMART). Brazil. Braz J Infect Dis.2011;15(1):34-39.
- Nguyen TN, Nguyen V C, Thwaites G, Carrique-Mas J. Antimicrobial Usage and Antimicrobial Resistance in Animal Production in Southeast Asia: A Review Antibiotics. 2016;5(37).
- Levy S, Fritz Gerald G, Macone A. Changes in intestinal flora of farm personnel after introduction of a tetracycline-supplemented feed on a farm. N Engl J Med. 1976; 295:583-588.
- Chen G, Hao H, Xie S, Wang X, Dai M, Huang L, Yua Z. Antibiotic alternatives: the substitution of antibiotics in animal husbandry? Frontiers in Mycrobiology. 2014;5(217).
- Pitout JD, Laupland KB. Extended-spectrum beta-lactamase-producing Enterobacteriaceae: an emerging public-health concern. Lancet Infect Dis. 2008;8:159-166.
- Verraes C , Van Boxstael S, Meervenne EV, Van Coillie E, Butaye P, Catry B, et al. Antimicrobial Resistance in the Food Chain: A Review Int. J EnvironRes Public Health.2013;10:2643-2669
- Lili G, Yeke T, Xiaodan Z, Jiaqing H, Zengmin M, Liangmeng W, Tongjie C. Emissions of Escherichia coli Carrying Extended-Spectrumβ-Lactamase Resistance from Pig Farms to the Surrounding Environment Int. J Environ Res Public Health.2015;12:4203-4213.
- Quiñones D, Valverde A, Rodríguez-Baños M, Kobayashi N, Zayaz A, Abreu M, Cantón R, Del Campo R. High Clonal Diversity in a Non-Outbreak Situation of Clinical ESBL-Producing Klebsiella pneumoniae Isolates in the First National Surveillance Program in Cuba. Microbial Drug Resistance. 2013.
- Instituto de Investigaciones Porcinas (IIP). Manual de Procedimientos Técnicos para la Crianza Porcina, MINAG, La Habana, Cuba. 2015.
- OIE. Código Sanitario para los Animales Terrestres. Capítulo 7.8. Utilización de animales en la investigación y educación. Decimonovena edición. Paris, France. 2014.
- Davis MF, Price L, Liu C, Silbergeld EK. An ecological perspective on U.S. industrial poultry production: the role of artificial ecosystems on the emergence of drug-resistant bacteria from agricultural environments. Current Opinion in Microbiology. 2011;14(3):244-250.
- OIE. Código Sanitario para los Animales Terrestres .Capítulo 6.7.- Armonización de los programas nacionales de vigilancia y seguimiento de la resistencia a los agentes antimicrobianos. 2016.
- Norma cubana NC 827: 2012.
- Khan F, Rizvi M, Shukla I, Malik A. A novel approach for identification of members of Enterobacteriaceae isolated from clinical samples. Biol Med. 2011;3(2):313-319.
- Clinical and Laboratory Standards Institute. (CLSI). Performance Standandars for Antimicrobial Susceptibility Testing 26eth ed. CLSI Supplment Informational Supplement M100S. Wayne: Clinical and Laboratory Standards Institute, USA.2016.
- Costa D, Poeta P, Sáenz Y, Vinué L, Rojo-Bezares B, Jouini A, et al. Detection of Escherichia coli harbouring extended-spectrum β-lactamases of the CTX-M, TEM and SHV classes in faecal samples of wild animals in Portugal. J Antimicrob Chemother. 2006;59:1311-1312.
- Briñas L., M. Lantero, I. de Diego, M. Alvarez, M. Zarazaga, Torres C. Mechanisms of resistance to expanded-spectrum cephalosporins in Escherichia coli isolates recovered in a Spanish hospital.. Antimicrob Chemother. 2005;56:1107-1110.
- InfoStat versión 1.1. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. 2002
- García CT, Castillo MA, Salazar RD. Mecanismos de resistencia a betalactámicos en bacterias gramnegativas Revista Cubana de Salud Pública Instituto de Medicina Tropical "Pedro Kourí". La Habana, Cuba. 2014. 40(1):129-135.
- García ML, Hendriksen RS, Fraile L and Aarestrup FM. Antimicrobial resistance of zoonotic and commensal bacteria in Europe: the missing link between consumption and resistance in veterinary medicine. Vet Microbiol. 2014;170:1-9.
- Navarro F, Miró E, Mirelis B. Lectura interpretada del antibiograma de enterobacterias. Enferm Infecc Microbiol Clin. 2010; 28(9): 638-645.
- Thomson KS. Controversies about extended-espectrum and AmpC beta-lactamases. Emerg. Infect Dis. 2017;333-336.
- Bradford PA. Extended-spectrum β-lactamases in the 21st century: characterization, epidemiology, and detection of this important resistance threat. Clin Microbiol Rev. 2001;14:933-951.
- Jacoby G, Muñoz S. The new β-lactamases. N Engl J Med; 2005; 352:380-391.
- Poirel L, Naas T, Thomas I, Karim A, Bingen E, Nordmann P. CTX-M-Type Extended-Spectrum ß-Lactamase That Hydrolyzes Ceftazidime through a Single Amino Acid Substitution in the Omega Loop. Antimicrobial Agents and Chemotherapy. 2008;45(12):3355-3361.
- Cano EM, Domínguez AM, Ezpeleta BC, Martínez ML, Padilla OB. Ramírez de Arellano E. Cultivos de vigilancia epidemiológica de bacterias resistentes a los antimicrobianos de interés nosocomial. Recomendaciones de la Sociedad Española de Procedimientos en Microbiología Clínica. 2007;(SEIMC). ISBN-978-84-611-9636-4.
- Treviño M, Martínez-Lamas L, Romero-Jung P, Varón C, Moldes L, et al. Comparación entre las pruebas para la detección de betalactamasas de espectro extendido de los sistemas Vitek2 y Phoenix. Enferm Infecc Microbiol Clin. 2009.
- Truppia LA, Mollerach A, di Conza JA. Comparación de tres métodos microbiológicos para la detección de betalactamasas de espectro extendido en enterobacterias aisladas en Santa Fe (Argentina). Enfermedades infecciosas y microbiología clínica. 2005;23(9):525-528.
- Von Salviati C, Laube H ,Guerra B, Roesler U , Anika Friese AEmission of ESBL/AmpC-producing Escherichia coli from pig fattening farms to surrounding areas. Vet Microbiol. 2015;175:77-84.
- Liao XP, Xia J, Yang L, Li L, Sun J, Liu Y-H, Jiang H-X. Characterization of CTX-M-14-producing Escherichia coli from food-producing animals. Front Microbiol. 2015;6:1136.
- Bonnet R. Growing Group of Extended-Spectrum β-Lactamases: the CTX-M Enzymes Antimicrob Agents Chemother. 2004;48(1):1-14.
- Hopkins KL, Davies RH, Threlfall EJ. Mechanisms of Quinolone Resistance in Escherichia coli and Salmonella: Recent Developments. Inter. J Antimicrob Agents. 2005;25(5):358-373.
- Schmithausen RM, Schulze-Geisthoevel SV, Stemmer F, El-Jade M, Reif M, Hack S, et al. Analysis of Transmission of MRSA and ESBLE among Pigs and Farm Personnel. 2015. PLoS ONE 10 (9): e0138173. doi:10.1371/journal.pone.0138173.
- Yang H, Chen S, While D, Zhao S, Mc Dermantt P, Walker R, Merig J. Characterization of multiple- antimicrobial- resistant Escherichia coli isolates from diseased chicken and swine in China. J Clin Microbiol. 2014;42(8):3483-3489.
- Livermore DM, and Woodford N. The beta-lactamase threat in enterobacteriaceae, pseudomonas and acinetobacter. Trends Microbiol. 2006;14(9):413-420.
- Torres C y Zarazaga M. BLEE en animales y su importancia en la transmisión a humanos Enferm Infecc Microbiol Clin. 2007;25 Supl. 2:29-37.
- Josef DJ, Stedt J and Gustafsson L. Zero prevalence of extended spectrum beta-lactamase-producing bacteria in 300 breeding Collared Flycatchers in Sweden. Infection Ecology and Epidemiology 2013;3:2
- Moodley A, Guardabassi L. Transmission of IncN plasmids carrying blaCTX-M-1 between commensal Escherichia coli in pigs and farm workers. Antimicrob Agents Chemother. 2009;53(4):1709-1711.
- Hammerum AM, Larsen J, Andersen VD, Lester CH, Skovgaard Skytte TS, Hansen F, et al. Characterization of extended-spectrum beta-lactamase (ESBL)-producing Escherichia coli obtained from Danish pigs, pig farmers and their families from farms with high or no consumption of third- or fourth-generation cephalosporins. J Antimicrob Chemother. 2014.
- Huijbers PM, Graat EA, Haenen AP, van Santen MG, van Essen-Zandbergen A, Mevius DJ, et al.Extended-spectrum and AmpC beta-lactamase-producing Escherichia coli in broilers and people living and/or working on broiler farms: prevalence, risk factors and molecular characteristics. J Antimicrob Chemother. 2014;69(10):2669-2675.
- Hota B. Contamination, disinfection, and cross-colonization: are hospital surfaces reservoirs for nosocomial infection? Clin Infect Dis. 2004; 39:1182-1189.
- Kramer A, Schwebke I, Kampf G. How long do nosocomial pathogens persist on inanimate surfaces? A systematic review. BMC Infectious Diseases. 2006;6:130.
- Bhalla A, Pultz NJ, Gries DM. Acquisition of nosocomial pathogens on hands after contact with environmental surfaces near hospitalized patients. Infect Control Hosp Epidemiol. 2004;25:164-167.
- Collineau L., Backhans A., Dewulf, J., Emanuelson U.F., Grosse B.E., Lehébel A., Loesken, S.E., Nielsen O., Postma M., Sjölund M., Stärk K.D.C, Belloc C.. Profile of pig farms combining high performance and low antimicrobial usage within four European countries Veterinary Record 201710.1136/vr.103988. Disponible http://veterinaryrecord.bmj.com
- Paivarinta M, Pohjola L, Fredriksson-Ahomaa M, Heikinheimo A. Low Occurrence of Extended-Spectrum b-lactamase-Producing Escherichia coli in Finnish Food-Producing Animals Zoonoses and Public Health Blackwell Verlag GmbH. 2016.
Recibido: 15/7/2017
Aceptado: 20/10/2017
* Autor para correspondencia: Ivette Espinosa-Castaño. E-mail: espinosa@censa.edu.cu


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}