










 Servicios personalizados
Servicios personalizados
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
INTRODUCCIÓN
Los crisomélidos Cerotoma trifurcata (Coleoptera: Chrysomelidae) y Cerotoma arcuata (Coleoptera: Chrysomelidae), son algunas de las mayores plagas que afectan el cultivo de soya a nivel mundial (Bortolotto et al., 2015). En Colombia, Cerotoma tingomariana (Bechyné) (Coleoptera: Chrysomelidae) es considerado una de las principales plagas limitantes para este cultivo. Este insecto puede causar reducciones significativas ocasionando daños hasta del 40 % en el área foliar y hasta un 45 % en la fijación de nitrógeno (Hadi et al., 2012). El manejo de la plaga se establece durante los periodos de floración y desarrollo de la planta con tres o cuatro aplicaciones de plaguicidas de síntesis química, los cuales tienen un efecto negativo sobre la salud humana y el medio ambiente; además, la aplicación excesiva conlleva al desarrollo de resistencia a estos químicos por parte del insecto plaga (Szendrei et al., 2012).
El uso de métodos alternativos para el manejo de la plaga como el control biológico es recomendado. En Brasil, los aislamientos fúngicos de Beauveria bassiana (Ascomycota: Hypocreales) y Metarhizium anisopliae (Ascomycota: Hypocreales) han sido evaluados contra larvas de segundo instar de Cerotoma arcuata (Coleoptera: Chrysomelidae) con mortalidades entre 80 y 100 % bajo condiciones de laboratorio y campo (Teixeira y Franco, 2007). De igual manera, la aplicación de estos hongos a dosis de 500 y 1000 g conidios/ha en suelo y de 250 y 500 g conidios/60 kg de semilla, mejoró el control de larvas de primer instar en condiciones de casa de malla (Quintela et al., 1990). Sin embargo, el uso de hongos entomopatógenos ha sido limitado debido a su baja eficacia y persistencia en condiciones de campo, como resultado de la baja tolerancia a condiciones abióticas de estrés (Rangel et al., 2015, Butt et al. 2016). Con el fin de seleccionar hongos entomopatógenos eficientes, la virulencia ha sido considerada como el factor más importante. Sin embargo, varios autores han establecido la importancia de la tolerancia a condiciones ambientales (temperatura, pH y radiación UV) en el proceso de selección los cuales impactan el desarrollo tecnológico de un bioplaguicida y su aplicación en campo (Shin et al., 2017). En ese sentido y con el propósito de contribuir al control biológico de C. tingomariana en cultivos de soya Glycine max (Fabales: Fabaceae) en Colombia, el objetivo de este estudio fue seleccionar y caracterizar ecofisiológicamente aislamientos de hongos entomopatógenos para el control de C. tingomariana, teniendo en cuenta su eficacia y tolerancia a condiciones como temperatura, radiación ultravioleta (UV-B) y pH.
MATERIALES Y MÉTODOS
Este trabajo se realizó en los laboratorios de entomología de los centros de investigación de La Libertad (Meta, Colombia) y Tibaitatá (Cundinamarca, Colombia) de la Corporación colombiana de investigación agropecuaria - AGROSAVIA. Se evaluaron seis aislamientos de Beauveria bassiana codificados como Bv003, Bv015, Bv025, Bv054, Bv060, Bv061; tres aislamientos de Metarhizium anisopliae codificados como Mt004, Mt031, Mt043, y un aislamiento de Metarhizium robertsii codificado como Mt015. Todos los aislamientos fueron suministrados por el Banco de Germoplasma con Interés en Control Biológico de AGROSAVIA y están cobijados dentro del contrato de acceso a recursos genéticos No. 168 de 2017. Los aislamientos se sembraron en Agar Papa Dextrosa (Sigma®) y se incubaron a 25 ± 2°C durante 7 días y fueron conservados a 4 ± 2 °C. Los adultos de C. tingomariana se colectaron de cultivos de soya en la Orinoquia, Colombia (Villavicencio - Meta, N 04.05922; W 073.45953), con una altitud de 336 m, precipitación anual promedio de 2933 mm y una temperatura promedio de 26°C, y posteriormente mantenidos en condiciones de casa de malla (T: 27 ± 5°C; RH: 75 ± 5 %).
Selección de hongos entomopatógenos para el control de C. tingomariana
La selección de los hongos entomopatógenos se realizó mediante la evaluación de la actividad biológica en condiciones de laboratorio, en donde se determinó la eficacia de los 10 aislamientos sobre C. tingomariana. Para esto, adultos de C. tingomariana se asperjaron con 20 mL de una suspensión de conidios de cada aislamiento, ajustada a una concentración de 1x107 conidios/mL en tween®80 al 0,1 %. Los adultos inoculados se ubicaron en cajas plásticas de 20 x 20 x 9 cm con hojas de fríjol provenientes de una siembra de plantas en casa de malla. Se tomaron las hojas de plantas en estado vegetativo. Las cajas plásticas se incubaron durante 20 días a 27 ± 4 °C y HR 70 ± 5 %. El experimento fue completamente al azar y cada tratamiento contó con tres réplicas de 10 individuos. El porcentaje de mortalidad fue determinado a los 20 días después de la inoculación y los resultados fueron corregidos con la mortalidad del tratamiento control para expresar la eficacia utilizando la fórmula de Schneider-Orelli (Zar, 1999):
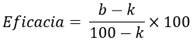
Donde, b = mortalidad en el tratamiento y k = mortalidad en el tratamiento control
Concentraciones letales de hongos entomopatógenos
Los aislamientos seleccionados se sembraron en Agar Papa Dextrosa (Sigma®) y fueron incubados a 25 ± 2 °C durante 7 días. Los conidios de cada aislamiento se recuperaron en una solución estéril de tween®80 al 0,1 %. Cada suspensión de conidios se ajustó a la concentración evaluada mediante el recuento de conidios en cámara de Neubauer. Para la evaluación de cada aislamiento, los adultos de C. tingomariana se inocularon con las concentraciones de 1x104, 1x105, 1x106, 1x107 y 1x108 conidios/mL y como tratamiento control se utilizó una solución tween®80 a 0,1 %. El diseño experimental fue completamente al azar y cada tratamiento contó con tres réplicas de 10 adultos. La mortalidad fue registrada diariamente y se determinaron las concentraciones letales mediante el análisis Probit, utilizando el programa POLO plus (versión 2.0) (LeOra Software 2002-2015).
Eficacia de hongos entomopatógenos en condiciones de casa de malla
Los hongos entomopatógenos seleccionados se evaluaron contra C. tingomariana en condiciones de casa de malla (T: 27 ± 5°C; RH: 75 ± 5 %). La unidad experimental consistió en una jaula de (90 x 90 x 120 cm) con dos plantas de soya de 30 días sembradas en materas de 5 kg con suelo. Utilizando una bomba de aspersión manual de 5 L, se aplicó la concentración letal noventa (CL90) de cada hongo, asegurando la cobertura del follaje (40 mL por planta). Como tratamiento control se aplicó una solución de tween®80 al 0,1 %. Posterior a la aplicación, se liberaron 10 adultos de C. tingomariana en cada jaula. El diseño experimental fue completamente al azar con 5 réplicas por tratamiento. La mortalidad se registró cada 5 días durante 20 días considerando la longevidad de los adultos que se encuentra entre 40 a 65 días. El porcentaje de mortalidad fue corregido con la mortalidad del control usando la fórmula de Schneider-Orelli (Zar, 1999).
Caracterización ecofisiológica de hongos entomopatógenos
Efecto de la radiación UV-B
Se preparó una suspensión de conidios a la concentración de 1x106 conidios/mL mediante recuento en cámara de Neubauer. Como variables respuesta se evaluaron la germinación de conidios y la viabilidad expresada en unidades formadoras de colonia (UFC). Para la germinación de conidios, se sembraron 100 µL en una caja de Petri de 9 cm de diámetro con agar agua suplementado con benomil al 0,00005 %. Las cajas de Petri inoculadas se expusieron a una dosis de 0,198 a 0,990 J/cm2 de radiación UV-B generada por una lámpara monocromática (Sylvania T8G30, 302 nm) durante 9 a 47 minutos. El tiempo de exposición fue determinado usando el Atlas de Radiación Solar en Colombia (IDEAM, 2018), en el que se encuentra registrada la intensidad de radiación de la zona de los Llanos Orientales de Colombia en donde se cultiva soya. Las cajas de Petri inoculadas e irradiadas se incubaron a 25 ± 2 °C durante 24 horas, tiempo después del cual se cuantificó el número de conidios germinados y no germinados mediante observación al microscopio (Axio Lab.A1 ZEISS, Alemania) de 10 campos ópticos, por unidad experimental. Se consideró como conidio germinado cuando este presentaba un tubo germinal con longitud igual o superior al diámetro del conidio. Para determinar el número de unidades formadoras de colonia UFC/mL, se inoculó una suspensión del hongo a una concentración de 102 a 103 conidios/mL en cajas de Petri con agar papa dextrosa. Las cajas de Petri inoculadas se expusieron a las dosis de irradiación como se mencionó anteriormente y se incubaron durante a 25 ± 2°C durante 7 días. El tratamiento control correspondió al hongo no expuesto a radiación e incubado bajo las mismas condiciones. El diseño experimental fue completamente al azar y para cada variable respuesta se emplearon 3 unidades experimentales por tratamiento.
Efecto del pH
Se evaluó el efecto de los valores de pH de 3, 5, 7 y 9 sobre las variables germinación y crecimiento diametral. Para la variable germinación, se utilizó el medio de cultivo agar agua suplementado con benomil 0,00005 % con pH ajustado a 3, 5, 7 y 9, en donde se sembraron 100 µL de una suspensión a 1x106 conidios/mL. Las cajas de Petri inoculadas se incubaron durante 24 horas a 25 ± 2 °C en oscuridad y posteriormente se estableció el porcentaje de germinación. Para el crecimiento diametral, un disco de 5 mm del hongo crecido se colocó en una caja de Petri con agar papa dextrosa ajustada a los diferentes valores de pH evaluados. A los 3, 6, 9, 12 y 15 días de incubación se midió crecimiento diametral de forma perpendicular en cada una de las réplicas. El diseño experimental fue completamente al azar y se emplearon tres unidades experimentales por tratamiento.
Análisis estadístico
La normalidad y homocedasticidad de los resultados se determinaron mediante las pruebas de Shapiro Wilks (95 %) y Bartlett (95 %), respectivamente. Una vez demostrados estos principios, se procedió a un análisis de varianza ANOVA y a una prueba de comparación de medias de Tukey (95 %) utilizando el programa Statistix® versión 8.0 (Analitycal Software, Florida, USA).
RESULTADOS Y DISCUSIÓN
Selección de hongos entomopatógenos para el control de C. tingomariana
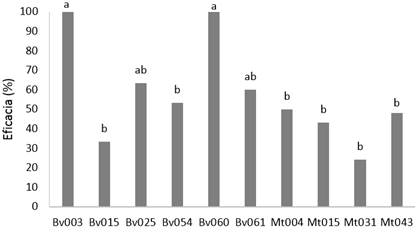
Los aislamientos de M. anisopliae y M. robertsii presentaron valores de eficacia inferiores al 50 % (F=0,68; gl= 8; p<0,05) mientras que los aislamientos de B. bassiana Bv003 y Bv060 tuvieron una eficacia del 100 %, seguidos por Bv025 y Bv061 con eficacias del 63,3 % y 60,0 %, respectivamente (Figura 1). Teniendo en cuenta estos resultados, se seleccionaron los aislamientos Bv003 y Bv060 por presentar una eficacia del 100 % y un rendimiento alto como característica deseable de producción (datos no mostrados). La mortalidad del tratamiento control fue de 6,6 %. La elevada mortalidad causada por estos aislamientos puede estar relacionada con una mayor expresión de factores de virulencia como lipasas, proteasas y quitinasas (Mondal et al., 2016).
Concentraciones letales de hongos entomopatógenos
La mortalidad de los insectos se relacionó con el aumento de la concentración de conidios de los aislamientos evaluados. La concentración letal media (CL50) fue de 5,8x105 conidios/mL y 5,6x104 conidios/mL para los aislamientos Bv060 y Bv003, respectivamente. Respecto a la concentración letal CL90, esta fue de 5,8x107 conidios/mL y de 1,3x106 conidios/mL, respectivamente (Tabla 1). Al calcular la potencia se demostró que el aislamiento Bv003 fue 10,4 veces más virulento que Bv060. Los resultados obtenidos de las concentraciones letales de Bv003 y Bv060 son similares a los reportados para el control de otros insectos. Espinel et al. (2014) evaluaron un aislado de Metarhizium rileyi (anteriormente Nomuraea rileyi) en larvas de Spodoptera frugiperda, obteniendo un CL50 de 1,17x104 conidios/mL, resultado muy similar al obtenido con este trabajo. También, Baja et al. (2020) reportó concentraciones letales CL50 de 3,5 x 105 con/mL y 6,8 x 106 con/mL al evaluar B. bassiana para el control de larvas de primer y segundo ínstar de Duponchelia fovealis y García et al. 2018, obtuvo una concentración de 9,0 x 106 con/mL para el control de larvas de segundo ínstar de Diatraea saccharalis.
Tabla 1 Concentraciones letales de aislamientos de B. bassiana sobre C. tingomariana en laboratorio
| Aislamiento | CL50 (conidios/ml) | CL90 (conidios/ml) | Límites de confianza LC50 (conidios/ml) | Límites de confianza LC90 (conidios/mL) | P | Heterogeneidad | |||
|---|---|---|---|---|---|---|---|---|---|
| inferior | superior | inferior | superior | χ2 | df | ||||
| Bv060 | 5,8 x 105 | 5,8 x 107 | 2,4 x 105 | 1,3 x 106 | 1,6 x 107 | 3,6 x108 | 0,759 | 1,54 | 3 |
| Bv003 | 5,6 x 104 | 1,3 x 106 | 2,3 x 104 | 1,1 x 105 | 5,9 x 105 | 5,9 x106 | 0,678 | 1,42 | 3 |
Eficacia de hongos entomopatógenos en casa de malla
El aislamiento Bv060 aplicado a la CL90 (5,8 x 107 conidios/ml) obtuvo una eficacia del 79,5 %, significativamente más alta que el aislamiento Bv003 (54,5 %) a una concentración de 1,3 x 106 conidios/mL (F=5,76; gl=1; P<0,05). La mortalidad en el tratamiento control fue del 12 %. El aislamiento Bv060 presentó una menor reducción en la eficacia en comparación con la eficacia obtenida en condiciones de laboratorio, siendo más tolerante a factores ambientales como la radiación UV, variaciones de temperatura y humedad. Esta tolerancia a condiciones de estrés está relacionada con diferentes mecanismos como la producción de trehalosa y polioles como el manitol que favorecen la estabilidad de las células, así como el origen de cada aislamiento, por lo cual puede presentar una mayor adaptación a dichas condiciones de estrés (Rangel et al., 2015; Shin et al., 2017).
Caracterización ecofisiológica de hongos entomopatógenos
Efecto de la radiación UV-B
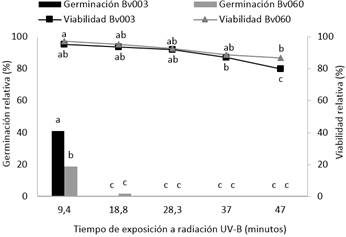
La radiación UV-B tuvo un efecto deletéreo sobre la germinación y viabilidad de los conidios de B. bassiana. Después de 9,42 minutos de exposición a la radiación, la germinación relativa de Bv003 fue significativamente más alta (40,0 %) comparada con el aislamiento Bv060 (18,9 %) (F=48,9, gl= 29; p < 0,001). Además, se obtuvo una germinación cercana al 2 % después de 18,8 minutos de exposición. Aunque la germinación fue completamente inhibida después de 18,8 minutos de exposición, la viabilidad relativa (UFC/mL) fue superior al 90 %, alcanzando viabilidades del 79,6 % para Bv003 y 86,8 % para Bv060 después de 47 minutos de radiación (F=9.83, gl = 29; p < 0,001) (Figura 2). Este efecto deletéreo puede estar relacionado con interrupción del ciclo celular, daño en la estructura del ADN, ARN, proteínas y membranas, así como la inducción de especies reactivas de oxígeno que afectan la viabilidad celular (Rangel et al., 2015, Santos et al., 2017, Wang et al. 2019).
Efecto del pH
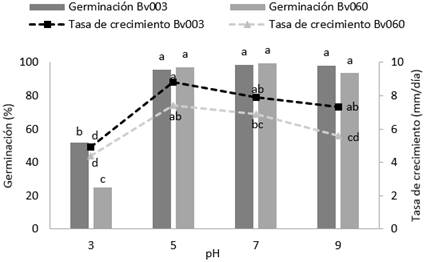
Los aislamientos Bv003 y Bv060 presentaron una alta tasa de crecimiento entre el rango de pH de 5 a 9, siendo mayor para el aislamiento Bv003 (F=20,5, gl= 23; p < 0,001) (Figura 3). Aunque el crecimiento diametral fue significativamente inhibido a pH de 9, la germinación fue superior al 90 % (F=241; gl=23; p < 0,001). Por otro lado, a pH 3 se inhibió significativamente la germinación y el crecimiento diametral de los hongos evaluados. Resultado similar al descrito por Zhu et al., (2016), quienes encontraron una disminución significativa en la germinación y en el proceso de conidiación de B. bassiana a los pH 3 y 9.
Efecto de la temperatura
A las cuatro temperaturas evaluadas, el crecimiento diametral de Bv003 fue significativamente más alto en comparación con el aislamiento Bv060; siendo la temperatura óptima para ambos aislamientos de 25 °C (F=149; gl=23; p < 0,001) (Figura 4). El aislamiento Bv060 fue más tolerante al incremento de temperatura al presentar una germinación del 88,4 % en comparación del 46,2 % obtenido por Bv003 a 30 °C (F=298; gl=23; p < 0,001), a diferencia de la temperatura de 15°C en donde el aislamiento Bv003 fue más tolerante. Lo anterior, puede estar relacionado con el origen del aislamiento Bv060 el cual fue recuperado de una región con mayor altitud. El crecimiento de los dos aislamientos fue inhibido significativamente a la temperatura de 35 °C.

Los dos aislamientos de B. bassiana tuvieron respuestas similares a las condiciones ambientales evaluadas, sin embargo, difieren en la mortalidad causada sobre C. tingomariana en condiciones de laboratorio y de casa de malla, lo que indica que podrían existir otros factores relacionados con la virulencia, como la actividad enzimática que interviene en el proceso de degradación de la cutícula de los insectos (Mondal et al., 2016) y la tolerancia a otras condiciones de estrés (Rangel et al., 2015). Además, la resistencia a condiciones extremas de temperatura y pH es una ventaja de los hongos entomopatógenos, ya que favorecen procesos como producción masiva y estabilidad de almacenamiento (Devi et al., 2005). El aislado Bv003 se seleccionó de acuerdo con la concentración letal más baja (10,4 veces más virulento que el aislado Bv060) y una mayor tasa de crecimiento bajo las variables de temperatura y pH. Estos resultados muestran el potencial de B. bassiana para controlar C. tingomariana y otras especies de Cerotoma como se demostró previamente con resultados satisfactorios (Teixeira y Franco 2007; Payah y Boethel 1986).
CONCLUSIONES
El aislamiento Bv003 perteneciente a la especie de B. bassiana fue seleccionado debido a su alta virulencia frente a C. tingomariana, mayor tolerancia a radiación UV-B y mayor tasa de crecimiento bajo diferentes valores de pH y temperatura. Este microorganismo Bv003 tiene un alto potencial para ser utilizado como principio activo de un bioplaguicida e implementado en un programa de manejo integrado de plagas y enfermedades

