










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCultivos Tropicales
versión impresa ISSN 0258-5936
cultrop v.30 n.1 La Habana ene.-mar. 2009
Influencia de un oligogalacturónido en la aclimatización de vitroplantas de banano (Musa spp.) del clon FHIA-18 (AAAB)
Ms.C. H. IzquierdoI y Dra.C. Miriam NúñezII, Dra.C. María C. GonzálezIII, Ruth ProenzaIV, y Dr.C. J. C. CabreraVI
I Investigador Auxiliar, email: hioviedo@inca.edu.cu
II Investigadora Titular del departamento de Fisiología y Bioquímica Vegetal
III Investigadora Titular del departamento de Genética y Mejoramiento Vegetal, Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas
IV Especialista de la Biofábrica Habana, San José de las Lajas, La Habana, Cuba
V Investigador de la Facultés Universitaires Notre-Dame de la Paix (FUNDP), Namur, Bélgica
ABSTRACT
The asexual reproduction is one propagating way of Musaceas; therefore, the application of different biotechnological techniques and further vitroplant acclimatization to environmental conditions is very important for its culture. This study was carried out at the National Institute of Agricultural Sciences (INCA) and Vitroplant-factory of San José de las Lajas, with the objective of evaluating oligogalacturonide effect on FHIA-18 banana vitroplant acclimatization. A mixture of organic matter and eutric compacted Red Ferralitic soil at a rate of 3:1 v/v was used as a substrate. After approximately 60 days, survival percentage, leaf number, pseudostem diameter, plant height, root number and length, as well as leaf proline content were evaluated. As a result, vitroplant root imbibition for 15 minutes before being planted and foliar spraying with oligogalacturonide at different concentrations (1, 5 and 10 mg.L-1) 15 days after planting increased plant survival by 8 % approximately compared to the control; every plant receiving oligogalacturonide were higher, had more amount of leaves and less foliar proline content.
Key words: in vitro culture, stress, oligosaccharides, growing media
RESUMEN
La reproducción asexual es una de las vías de propagación de las Musáceas, por lo que la aplicación de diferentes técnicas biotecnológicas y la posterior aclimatización de las plantas a las condiciones ambientales son de gran importancia para su cultivo. Teniendo en cuenta lo antes expuesto, se desarrolló este trabajo en el Instituto Nacional de Ciencias Agrícolas (INCA) y en la Biofábrica de San José de las Lajas, con el objetivo de evaluar el efecto que ejerce un oligogalacturónido en la aclimatización de las vitroplantas de banano del clon ‘FHIA-18’. Se utilizó como sustrato una mezcla de materia orgánica y suelo del tipo Ferralítico Rojo compactado éutrico en una relación 3:1 v/v. A los 60 días aproximadamente, se evaluaron el porcentaje de supervivencia, número de hojas, diámetro del pseudotallo, altura de las plantas, número y largo de las raíces, así como el contenido de prolina foliar. Como resultado se obtuvo que la imbibición de las raíces de las vitroplantas durante 15 minutos antes de su plantación y la aspersión foliar con el oligogalacturónido a diferentes concentraciones (1, 5 y 10 mg.L-1), 15 días después de la plantación incrementó la supervivencia de las plantas en un 8 % aproximadamente con respecto al control; todas las plantas en las que se utilizó el oligogalacturónido presentaron mayor altura, número de hojas y menor contenido de prolina foliar.
Palabras clave: cultivo in vitro, estrés, oligosacáridos, substratos de cultivo
INTRODUCCIÓN
Los primeros informes sobre micropropagación en plátanos y bananos (Musa spp.) se realizaron en la década de 1970 en algunos genotipos triploides (AAA). Posteriormente se incrementaron los estudios a otros genotipos, como son: AA, AAB, ABB y BBB, siendo la principal fuente de explante primario los ápices vegetativos. En Cuba la propagación in vitro de bananos es una realidad en la red nacional de biofábricas existentes en el país, lo que permite la producción de grandes volúmenes de vitroplantas, la conservación de germoplasma nativo o introducido y la realización de ensayos de comportamiento ante factores medioambientales y respuesta a sustancias orgánicas (1, 2).
Dentro del proceso de micropropagación se incluye como última fase la aclimatización denominada como período de endurecimiento (3). En esta fase las vitroplantas se plantan en envases individuales y se dejan durante un tiempo prudencial en condiciones de aviveramiento, mientras alcanzan el tamaño óptimo para su trasplante a campo.
La actividad biológica de los oligogalacturónidos se observa tanto en las plantas monocotiledóneas como en las dicotiledóneas (4, 5). Esta respuesta es intrínseca del tipo, la especie y variedad de planta, así mismo depende de la estructura química del oligogalacturónido que se utilice (6).
El grado de polimerización de los oligogalacturónidos es el factor estructural de mayor significación en la definición de su función biológica (7). La mayoría de los estudios de dependencia estructura–actividad evidencian que un grado de polimerización igual a nueve es el tamaño mínimo de los olígómeros biológicamente activos. Sin embargo, los fragmentos pépticos con grado de polimerización mayor de 16 presentan actividad biológica, pero esos tienen poca movilidad en el apoplasto, por lo que tienen menor importancia como moléculas señalizadoras (8), debido a que es posible que la misma pared celular actúe como una barrera para la interacción de estos macrofragmentos con la membrana citoplasmática (9).
Por otra parte, en Cuba se han obtenido resultados satisfactorios en la síntesis y validación agrobiológica de oligogalacturónidos con grado de polimerización entre 9 y 16 moléculas de ácido galacturónido, que se produce por la degradación parcial de la pared celular de la corteza de los cítricos y tiene una fracción molar que oscila entre 10.4 y 7.2 % (10, 11). Es un estimulante del crecimiento y diferenciación celular de diferentes especies vegetales, presenta un efecto biológico en los medios de cultivo de células vegetales in vitro, similar al de las auxinas o citoquininas, lo que depende del balance hormonal del explante y la composición de reguladores del crecimiento en el medio, pudiendo sustituir parcial o totalmente a los reguladores del crecimiento tradicionalmente empleados en la de micropropagación; además, puede activar los mecanismos de defensa y disminuir o atenuar el estrés ambiental de las plantas.
El oligogalacturónido de producción nacional ha sido empleado con buenos resultados en diferentes procesos biotecnológicos, tales como: embriogénesis somática en caña de azúcar (Saccharum spp.) y mandarina (Citrus reshni Hort. et Tan.) variedad ‘Cleopatra’ (12, 13), en el crecimiento y desarrollo de tomate (Solanum lycopersicum Mill.) a partir de semillas (14) y en Anthurium andreanum L. (15), enraizamiento de pecíolos de violeta africana (Saintpaulia ionantha L.) (16), así como la obtención de semilla artificial de caña de azúcar (Saccharum sp.) (17). Sin embargo, aunque ha sido informada su utilización en la aclimatización de vitroplantas de Anthurium cubense L. con buenos resultados (18), es insuficiente aún el conocimiento que se tiene en los diferentes procesos mencionados con anterioridad, así como en la aclimatización de los distintos genotipos de plantas de plátanos y bananos, en los que por lo general la supervivencia es baja.
El trabajo se realizó con el objetivo de evaluar el efecto que ejerce un oligogalacturónido en la aclimatización de las vitroplantas de banano (Musa spp.) del clon FHIA-18’(AAAB).
MATERIALES Y MÉTODOS
Como material vegetal, se emplearon las vitroplantas de la fase de enraizamiento in vitro del clon ‘FHIA-18’ (AAAB), procedentes del proceso productivo de la Biofábrica de San José de las Lajas, provincia La Habana.
El proceso se realizó según la metodología establecida (19). Las vitroplantas se plantaron en bandejas de polieturano de 70 alvéolos (cepellones), que contenían un sustrato compuesto por materia orgánica (cachaza) y suelo Ferralítico Rojo compactado éutrico (20), en una proporción 3:1 v/v, al que se le garantizó el riego por nebulización en los primeros 10 días, para lograr una alta humedad relativa (95-100 %) y adecuada humedad. Para el sombreo se utilizó una malla zarán negra (70 % de reducción de luz solar). Las características químicas del sustrato se muestran en la Tabla I.

Se empleó un tratamiento control sin aplicación del oligogalacturónido y tres tratamientos con oligogalacturónido (OLG), cuya masa molar es de 2042 g.mol-1; la aspersión se realizó a razón de 2 mL por vitroplanta. Las especificaciones de los tratamientos se muestran en la Tabla II.

Los indicadores que se evaluaron una vez finalizada la aclimatización fueron: porcentaje de supervivencia y altura de las plantas, cuya medición se realizó a 20 plantas desde la base hasta el ápice de la última hoja extendida y seguidamente se calculó la media; también se evaluaron el número de hojas por planta, número y largo de las raíces, así como el contenido de prolina libre en las hojas, para lo cual se emplearon cinco plantas por tratamiento y la determinación cuantitativa se realizó según la metodología descrita por Bates (21).
En el experimento se empleó un diseño completamente aleatorizado, con 125 plantas por tratamiento y se repitió dos veces en la misma época de plantación (febrero). Los datos se procesaron a través de un ANOVA simple y las diferencias entre las medias se compararon según la prueba de rangos múltiples de Duncan al 95 % (22); previo a estos análisis se chequeó la distribución normal (Kolmogorov-Smirnov) y homogeneidad de varianza (Bartlett) de los datos.
RESULTADOS Y DISCUSIÓN
El porcentaje de sobrevivencia es un indicador muy importante en la fase de aclimatización, pues de él depende la cantidad de plantas que pueden llevarse a vivero y posteriormente al campo.
En la Figura 1
se observan los resultados que se obtuvieron con respecto a este indicador. Como puede apreciarse, los tratamientos 2 (1 mg.L-1) y 3 (5 mg.L-1) obtuvieron valores superiores al control (tratamiento 1), difiriendo significativamente del control y el tratamiento 4 (10 mg.L-1), que con un 80 % de sobrevivencia fue el peor. En el caso particular del clon ‘FHIA-18’, en las primeras cuatro semanas es cuando ocurre la mayor cantidad de muertes.

La respuesta favorable a los tratamientos anteriores ante este indicador puede explicarse según los planteamientos que afirman que los oligogalacturónidos pueden funcionar como señales moleculares que regulan la adaptación al ambiente (4, 9). Por su parte, otros señalan que estas sustancias disparan los mecanismos de defensa ante el estrés ambiental (1). Se han informado resultados similares en este genotipo con respecto al porcentaje de sobrevivencia empleando un estimulador del crecimiento cubano (10).
En las biofábricas cubanas lograr que la sobrevivencia de la plantas supere el 85 % es fundamental, pero si se logra que este parámetro sobrepase el 90 % es beneficioso, ya que esto garantiza la eficiencia del proceso de micropropagación, específicamente en el clon ‘FHIA-18’, que es uno de los que más se propagan empleando estas técnicas biotecnológicas, por ser resistente a la Sigatoka negra, pero en condiciones in vitro tiene un bajo coeficiente de multiplicación. En otros genotipos como ‘Prata Anã’, es necesario una fase de pre-acondicionamiento antes de transferir las plantas a la fase de aclimatización, para lograr una elevada sobrevivencia (23), lo cual no fue necesario en las condiciones en que se desarrolló este experimento, debido a que con el empleo del oligogalacturónido se logra que las plantas procedentes del cultivo in vitro presenten una mayor resistencia y tolerancia a las condiciones del estrés durante esta fase de aclimatización.
La altura de las plantas en la fase de aclimatización es otro de los indicadores que reviste gran importancia. Como se observa en la Figura
2, hay un incremento de este parámetro de 0.13 a 0.46 cm; todos los tratamientos en que se aplicó el OLG se diferenciaron estadísticamente del control. El tratamiento en que se emplea la menor concentración (1 mg.L-1) alcanza los mejores resultados con 6.32 cm.
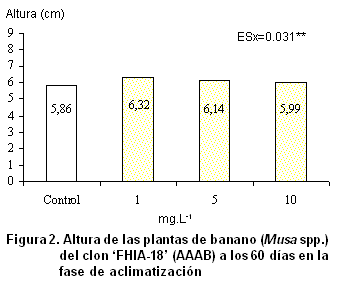
El incremento de la altura de la planta mediante la utilización de nuevas sustancias de producción nacional que estimulan el crecimiento ha sido ampliamente discutido (24, 25, 26), los cuales coinciden en conferirle una gran importancia a este parámetro. Los oligogalacturónidos son capaces de influir en diferentes procesos fisiológicos que ocurren en el vegetal.
Otra posible vía por la cual se incrementa el tamaño de la planta es porque las oligosacarinas pueden estimular la actividad fotosintética; por tanto, hay una mayor ganancia de esqueletos carbonados que pueden ser utilizados para la síntesis de nuevos compuestos, como son las proteínas (27, 28).
En relación con el número de hojas (Figura
3), se observa que las plantas que se trataron con el OLG se favorecieron, ya que se diferenciaron estadísticamente del control.
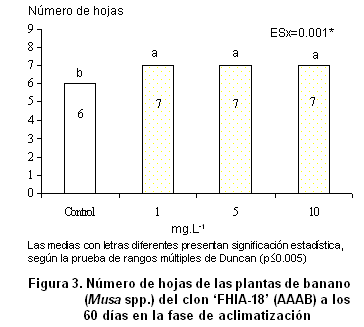
El efecto positivo del oligogalacturónido sobre el número de hojas evidencia la acción que ejerce sobre los procesos fisiológicos como la elongación, el alargamiento y la diferenciación celular, lo cual ha sido estudiado en este y otros cultivos (2, 28).
Las hojas de las plantas que se producen in vitro se emplean como almacén de sustancias carbonadas y se utilizan en el crecimiento y desarrollo de las plántulas; luego de su transferencia de las condiciones in vitro a ex vitro, mantienen esta función hasta tanto no exista una nueva emisión foliar (29).
El diámetro del pseudotallo indica el grosor de las vitroplantas (Tabla
III). Como se puede observar, hubo diferencias significativas: los mejores resultados fueron para los tratamientos 2 y 3, seguido del 4, pero este último no se diferenció estadísticamente del control. Estos resultados coinciden parcialmente con los informados anteriormente (10, 18), donde la concentración más elevada (10 mg.L-1) permitió alcanzar los mejores resultados y en este trabajo la concentración más baja (1 mg.L-1) fue la más efectiva. Algunos plantean que el Pectimorf actúa de forma sinérgica con las auxinas, específicamente con el AIA, por lo que los resultados de este trabajo son similares a otros (17).

En la misma Tabla III se observa, además, el número y largo de las raíces de las plantas, que en ambos casos hubo diferencias significativas. Respecto al número de raíces, los tratamientos 2 y 3 fueron los que alcanzaron los mejores resultados, superando al 1 y 4. Cuando se empleó el OLG a las diferentes concentraciones, aumentó el largo de las raíces.
Los oligogalacturónidos y las auxinas actúan de forma antagónica, modulando el crecimiento y la morfogénesis de las raíces, no solo en explantes de hojas sino también en fragmentos de plantas transgénicas de tabaco que contienen el oncogen rolB proveniente de Agrobacterium rhizogenes (30). Otros también plantean que los oligogalacturónidos son inhibidores del enraizamiento inducido por auxinas (27, 31). Así mismo, se informa que estos pueden estimular o inhibir el enraizamiento en dependencia de la fracción del oligosacárido de origen péctico que haya empleado y del material vegetal (14), lo que indica que las respuestas vinculadas al crecimiento y desarrollo de las plantas dependen del balance hormonal endógeno del explante, medio de cultivo y su procedencia.
Los resultados antes expuestos son contrarios a algunos anteriores, ya que este OLG estimula la rizogénesis, porque al parecer actúa de forma sinérgica y no antagónica con las auxinas endógenas de las plantas de banano del clon FHIA-18. En tal sentido, utilizando este oligogalacturónido, se ha informado un aumento en el número de raíces en margullos de Ficus benjamina de la variedad ‘Golden King’ y ‘Nítida’ en dos y 18 raíces como promedio, respecto al enraizador comercial Rhizopon-B, que es un producto químico (25).
El contenido de prolina libre registrado en las plantas se muestra en la Figura
4. Las plantas del tratamiento 1 (control) fueron las que mayor valor alcanzaron, ya que estuvieron sometidas a un mayor estrés ambiental, debido a que no se sometieron a ningún tratamiento con el OLG. La reducción significativa del contenido de prolina libre en las hojas de las plantas, a medida que se redujo la concentración del oligogalacturónido de 10 a 1 mg.L-1, demuestra su efecto antiestrés.
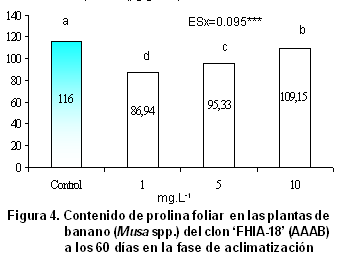
Las pectinas se encuentran entre los polisacáridos más abundantes de la pared celular de los vegetales; estos polisacáridos son altamente hidrofílicos y el agua que ellos atraen a la pared celular les proporciona flexibilidad, que es un requisito indispensable para su alargamiento (32, 33), por lo que la imbibición previa de las raíces y posterior aspersión foliar de las plantas con el oligogalacturónido fue útil para los tejidos en crecimiento, donde aún no se ha formado una pared celular secundaria, que es característica de los tejidos adultos.
La aplicación de esta sustancia péctica en las fases iniciales del crecimiento de las vitroplantas durante la fase de aclimatización proporcionó una mejor adaptación a las condiciones ex vitro, por lo que el éxito de la aclimatización depende de que las plantas puedan pasar de condiciones heterotróficas o mixotróficas (mezcla de autotróficas con heterotróficas) al autotrofismo, proceso que obedece en su generalidad a las reservas que adquieren las plántulas durante la fase in vitro (34, 35, 36, 37, 38). Por consiguiente, al estar las plántulas menos estresadas, se recuperan más rápido y presentan mejores condiciones para su trasplante definitivo a campo.
Los resultados de este trabajo constituyen la base para futuros estudios relacionados con la morfogénesis, fisiología y bioquímica de este oligogalacturónido, el cual pudiera aplicarse eficientemente en los sistemas agrícolas sin que se afecte el medio ambiente, por ser un producto natural. Sin embargo, se plantea la necesidad de profundizar en los mecanismos de acción de este oligogalacturónido a nivel molecular (12), para lograr determinar cuáles son realmente las vías con las que ejerce su acción, ya sea con las auxinas y/o citoquininas que están involucradas en los diferentes procesos biotecnológicos.
AGRADECIMIENTOS
A la Dra.C. Amelia Capote, del Instituto de Investigaciones Fundamentales en Agricultura Tropical “Alejandro de Humboldt” (INIFAT), y a la Dra.C. María E. González, del departamento de Genética y Mejoramiento Vegetal del Instituto Nacional de Ciencias Agrícolas (INCA), por la revisión exhaustiva y las sugerencias que le realizaron al trabajo para su publicación.
REFERENCIAS
1. Pérez, J. N.; Suárez, M. y Orellana, P. Posibilidades y potencial de la propagación masiva de plantas en Cuba. Biotecnología Vegetal, 2000, vol. 1, p. 3-12.
2. Gübbük, H. y Pekmezc, M. In vitro propagation of some new banana types (Musa spp.). Turk. J. Agric. For., 2004, vol. 28, p. 355-361.
3. Murashige, T. Clonal crops through tissue culture. En: Barz, W.; E. Reinhard y M. H. Zenk (Eds.). Tissue culture and its biotechnological application. New York, 1977, p. 392-403.
4. Moerschbacher, B. M.; Mierau, M.; Graebner, B.; Noll, U. y Mort, A. J. Small oligomers of galacturonic acid and endogenous suppressors of disease resistance reaction in wheat leaves. J. Exp. Bot., 1999, vol. 50. p. 605-612.
5. Inngjerdingen, K.; Debes, S.; Inngjerdingen, M.; Hokputsa, S.; Harding, S.; Rolstad, B.; Michaelsen, T.; Diallo, D.; Paulsen, B. Bioactive pectic polysaccharides from Glinus oppositifolius (L.) Aug. DC., a Malian medicinal plant, isolation and partial characterization. Journal of Ethnopharmacology, 2005, vol. 101, no. 3, p. 204-214.
6. Shibuya, N. y Minami, E. Oligosaccharide signalling for defence responses in plant. Physiological and Molecular Plant Pathology, 2001, vol. 59, p. 223- 233.
7. Monsoor, M. Effect of drying methods on the functional properties of soy hull pectin. Carbohydrate Polymers, 2005, vol. 63, no. 3, p. 362-367.
8. Messiaen, J. y Van Cutsem, P. Defense gene transcription in carrot cells treated with oligogalacturónidos. Plant Cell Physiology, 1993, vol. 34, no. 7, p. 111-112.
9. Frirdich, E.; Bouwman, C.; Vinogradov, E.: Whitfield, C. The role of galacturonic acid in outer membrane stability in Kiebsiella pneumoniae. Journal of Biological Chemistry, 2005, vol. 280, no. 30, p. 27604-27612.
10. Cabrera, J. C. Obtención de una mezcla de oligogalacturónidos a partir de corteza de cítrico. Tesis en opción al grado de Doctor en Ciencias Químicas. INCA, 2000, p. 25-67.
11. Cabrera, J. C.; Gómez R.; Diosdado, E.; Hormaza, J. V.; Iglesias, R.; Gutiérrez, A. y González, S. Procedimiento de obtención de una mezcla de oligosacáridos y pécticos estimuladora del enraizamiento vegetal. Cuba, patente de invención. Certificado 22859. 2003, RES 155/2003.
12. Nieves, N.; Poblete, A.; Cid, M.; Lezcano, Y.; González-Olmedo, J. L.; Cabrera, J. C. Evaluación del Pectimorf como complemento del 2,4-D en el proceso de embriogénesis somática de caña de azúcar (Saccharum spp.). Cultivos Tropicales, 2006, vol. 27, no. 1, p. 25-30.
13. Hernández, R. M.; Lara, R. M.; Diosdado, E.; Cabrera, J. C.; González, C.; Valdés, M.; Xiqués, X. Evaluación de la actividad del Pectimorf en la embriogénesis somática de la mandarina ‘Cleopatra’ (Citrus reshni Hort. et Tan.) mediante marcadores isoenzimáticos. Cultivos Tropicales, 2007a, vol. 28, no. 4, p. 25-31.
14. Costales, D; Martínez, L y Núñez, M. Efecto del tratamiento de semillas con una mezcla de oligogalacturónidos sobre el crecimiento de plántulas de tomate (Lycopersicon esculentum Mill.). Cultivos Tropicales, 2007, vol. 28, no. , p. 85-91.
15. Hernández, L.; Benítez, B.; Soto, F.; Dominí, M. E. Efecto de una mezcla de oligogalacturónidos en el crecimiento y desarrollo de Anthurium andreanum. Cultivos Tropicales, 2007b, vol. 28, no. 4, p. 83-86.
16. Falcón, A. B. y Cabrera, J. C. Actividad enraizadora de una mezcla de oligogalacturónidos en pecíolos de violeta africana (Saintpaulia ionantha L.). Cultivos Tropicales, 2007, vol. 28, no. 2, p. 87-90.
17. Cid, M.; González-Olmedo, J. L.; Lezcano, Y.; Nieves, N. Influencia del Pectimorf sobre la calidad de la semilla artificial de caña de azúcar (Saccharum sp.). Cultivos Tropicales, 2006, vol. 27, no. 1, p. 31-34.
18. Montes, S.; Aldaz, J. P.; Cevallos, M.; Cabrera, J. C. y López, M. Uso del biorregulador Pectimorf en la propagación acelerada del Anthurium cubense. Cultivos Tropicales, 2000, vol. 21, no. 3, p. 29-31.
19. Anon, J. Fase IV. Adaptación o aclimatización a las condiciones ambientales. pp. 18-20. En: Instructivo técnico para la micropropagación de plátanos y bananos. Capítulo 8. Empresa de semillas, La Habana, Cuba., 2003, p. 3-9.
20. Cuba. MINAG. Nueva versión de clasificación genética de los suelos de Cuba. Instituto de Suelos. Ministerio de la Agricultura. La Habana. Agrinfor, 1999, 64 p.
21. Bates, L. S.; Waldron, R. D. y Team, L. D. Rapid determination of free-proline for water stress studies. Plant and Soil, 1973, vol. 39, no. 1, p. 205-207.
22. Cochran, W. y Cox, G. Diseños experimentales. C. México. Ed. Trellas, 1990, p. 132-135.
23. Toledo, M.; Nietsche, S.; Cabral, A.; Ferreira, C.; de Lima, C.; Dias, V.: Panicalle, B.; Batista, D. L. y Koji, M. Aclimatização de mudas micropropagadas de bananeira sob diferentes condições de luminosidade 1. Rev. Bras. Frutic., 2005, vol. 27, no. 2, p. 238–240.
24. González-Olmedo, J. L.; Córdova, A.; Aragón, C. E.; Pina, D.; Rivas, M. y Rodríguez, R. Efecto de un análogo de brasinoesteroides sobre plántulas de ‘FHIA-18’ expuestas a un estrés térmico. INFOMUSA, 2005, vol. 14, no. 1, p. 18-20.
25. Dominí, M. E. y Benítez, B. Uso de biopreparados como promotores del enraizamientos en margullos de Ficus (Ficus benjamina). Cultivos Tropicales, 2004, vol. 25, no. 3, p. 45-48.
26. Nuri, M. y Read, P. E. Improved rooting and acclimatization of micropropagated Hazelnut shoots. HortScience, 2004, vol. 39, no. 7, p. 132-137.
27. Albersheim, P. y Darvill, A. G. Oligosaccharins. Sci. Ann., 1985, vol. 253, no. 3, p. 58–64.
28. El-Sharkawy, M. A. Utility of basic research in plant/crop physiology in relation to crop improvement: a review and a personal account. Bras. J. Plant Physiol, 2006, vol. 18, no. 4, p. 419-446.
29. Van Huylenbroeck, J. M.; Piqueras, A. y Debergh, P. C. Photosynthesis and carbon metabolism in leaves formed prior and during ex vitro acclimatization of micropropagated plants. Plant Science, 1998, vol. 134, p. 21-30.
30. Bellincampi, D.; Cardarelli, M.; Zaghi, D.; Serino, G. y Salvi. G. Oligogalacturonides prevent rhizogenesis in rolB-transformed tobacco explants by inhibiting auxin-induced expression of the rolB gene. The Plant Cell, 1996, vol. 8, p. 477-487.
31. Hernández, G.; Sepúlveda, B.; Richards, A.; Soriano, E. The architecture of Phaseolus vulgaris roots is altered when a defense response is elicited by an oligogalacturonide. Braz. J. Plant Physiol., 2006, vol. 18, no. 2, p. 351-355.
32. Frirdich, E.; Bouwman, C.; Vinogradov, E. y Whitfield, C. The role of galacturonic acid in outer membrane stability in Kiebsiella pneumoniae. Journal of Biological Chemistry, 2005, vol. 280, no. 30, p. 27604-27612.
33. Moscatiello, R.; Mariani, P.; Sanders, D. y Maathuis, F. J. Transcriptional analysis of calcium-dependent and calcium-independent signalling pathway induced by oligogalacturonides. Journal of Experimental Botany, 2006, vol. 57, no. 11, p. 2847-2865.
34. Aragón, C.; Escalona, M.; Capote, I.; Pina, D.; Cejas, I.; Rodríguez, R.; Noceda, C.; Sandoval, J.; Roels, S.; Debergh, P. y González-Olmedo, J. L. Importancia metabólica del almidón en la aclimatización de plantas de plátano CEMSA 3/4 (AAB). INFOMUSA, 2006, vol. 15, no. 1-2, p. 32-35.
35. Martínez, R.; Azpíroz, R. H.; Rodríguez de la O, J. L.; Cetina, V. M. y Gutiérrez, M. A. Aclimatización de plantas obtenidas in vitro de Eucalyptus urophylla S. T. Blake y Eucalyptus grandis Hill ex Maiden. Ra Ximhai. Universidad Autónoma Indígena de México, 2005, vol. 1, no. 3, p. 591-597.
36. Rodríguez, R. Aclimatización de plántulas de caña de azúcar (Saccharum sp. híbrido) propagandas en biorreactores de inmersión temporal. Tesis presentada en opción al Grado Científico de Doctor en Ciencias Agrícolas. Centro de Bioplantas. Universidad de Ciego de Avila, 2005, 95 p.
37. Viegas, P. H.; Liner, A. M.; Ambrosano, G. M. y Batista, M. de F. Acclimatization of micropropagated Heliconia bihai (Heliconiaceae) plants. Sci Agric., 2005, vol. 62, no. 3, p. 13-18.
38. Abreu, E.; González, G.; Ortíz, R.; Rodríguez, P.; Domech, R. y Garriga, M. Evaluación de vitroplantas de henequén (Agave fourcroydes Lem.) durante la fase de aclimatización. Cultivos Tropicales, 2007, vol. 28, no. 1, p. 5-11.
Recibido: 30 de mayo de 2008
Aceptado: 19 de enero de 2009
