










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCultivos Tropicales
versión impresa ISSN 0258-5936
cultrop vol.31 no.3 La Habana jul.-set. 2010
Efecto del análogo de brasinoesteroide (Biobras-16) en la germinación y el crecimiento inicial de las plantas de dos variedades de tomate en condiciones de estrés salino
Effect of brassinosteroid analogue (Biobras-16) on the germination and early growth of plants of two tomato varieties in saline stress conditions
Yanelis ReyesI, Dr.C. L. M. MazorraII, Lisbel MartínezIII y Dra.C. Miriam NúñezIV
I Reserva Científica. Email: yanelisrg@inca.edu.cu
II Investigador Agregado
III Especialista
IV Investigador Titular del departamento de Fisiología y Bioquímica Vegetal, Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, La Habana, Cuba, CP 32 700.
RESUMEN
En este trabajo se estudió el efecto del análogo espirostánico de brasinoesteroide (Biobras-16, BB-16) en la germinación y el crecimiento inicial de plántulas de tomate de las variedades Amalia e INCA 9(1) en condiciones de estrés salino. Para ello, primeramente se realizó un ensayo para seleccionar el tiempo de imbibición de las semillas en agua y las condiciones de salinidad, que afectaba la germinación y el crecimiento de las plántulas de ambas variedades. Se evaluaron tres tiempos de imbibición (0 ,4 y 8 h) y tres concentraciones de NaCl (50, 75 y 100 mM). A partir de los resultados de este ensayo, se seleccionó la concentración de 75 mM y el tiempo de imbibición de 4 h. Posteriormente, se evaluó el efecto del BB-16 en la dinámica de germinación y el crecimiento inicial de las plántulas. Para ello, se colocaron semillas de estas dos variedades durante siete días con diferentes concentraciones de BB-16 (10-10-10-6 mol.L-1) en presencia o no de 75 mM de NaCl. A esta concentración de NaCl se evidenció un efecto protector en el crecimiento inicial de las plántulas de la concentración de 10-7 mol.L-1 de BB-16 en la variedad INCA 9(1), aunque en la dinámica de germinación, a las 48 h, se observó el efecto estimulador a la menor concentración ensayada. La concentración más alta (10-6 mol.L-1) de este análogo inhibió la germinación y el crecimiento de las plántulas en esta variedad. En la variedad Amalia no se observó efecto protector con respecto al control.
Palabras clave: brasinoesteroides, hinchamiento de la semilla, salinidad, tomate, tolerancia a la sal.
ABSTRACT
This work studied the effect of a brassinosteroid spirostane analogue (Biobras-16, BB-16) on tomato seedling germination and early growth of Amalia and INCA 9(1) varieties under saline stress conditions. Firstly, a trial was carried out to select seed imbibition length in water and salinity conditions affecting seedling germination and growth of both varieties. Three imbibition times (0, 4 and 8 h) and three NaCl concentrations (50, 75 and 100 mM) were evaluated. Experimental results enabled to choose the concentration of 75 mM and imbibition time of 4 h. Furthermore, the effect of BB-16 was evaluated on seedling germination dynamics and early growth. Thus, seeds from both varieties were put for seven days with different BB-16 concentrations (10-10-10-6 mol.L-1) in the presence or not of 75 mM of NaCl. A protective effect on the early seedling growth of INCA 9(1) variety became evident at this NaCl concentration from the one of 10-7 mol.L-1 of BB-16; however, the promoting effect to the smallest experimental concentration was observed at the germination dynamics of 48 h. The highest concentration (10-6 mol.L-1) of this analogue inhibited seedling germination and growth in this variety. There was not any protective effect in Amalia variety compared to the control.
Key words: brassinosteroids, seed filling, salinity, tomato, salt tolerance.
INTRODUCCIÓN
La salinidad de los suelos es uno de los estrés más perjudiciales para los cultivos en la actualidad. La inadecuada irrigación de los suelos, así como el cambio climático hacen que este fenómeno alcance un nivel global. Existen informes que demuestran que el área de nuestro planeta afectada por la salinización es de alrededor de 800 millones de hectáreas (1). En Cuba, la superficie agrícola está afectada en un 14 % y otro 15 % más presenta peligros potenciales de salinización (2).
Durante el desarrollo de las plantas, en oportunidades las semillas son las primeras en enfrentar las condiciones de estrés. La salinidad afecta tanto la reanudación del crecimiento activo del embrión como el crecimiento inicial de la plántula, a través de su influencia en los diferentes procesos fisiológicos y bioquímicos, principalmente por su efecto en las relaciones hídricas así como la toxicidad de los iones (3).
Con esta problemática urge la necesidad de encontrar productos naturales, ecológicamente inocuos, capaces de proteger a las plantas contra este estrés y que sean viables para nuestra economía. Una de estas moléculas prometedoras por sus efectos protectores son los brasinoesteroides, que son potentes reguladores del crecimiento vegetal de naturaleza esteroidal. Estas hormonas tienen efectos pleiotrópicos: estimulan el alargamiento celular y la desdiferenciación de protoplastos, regeneran la pared celular, regulan la diferenciación de elementos traquearios, e incrementan la biomasa y el rendimiento (4). Además, se ha informado el efecto protector de los brasinoesteroides naturales ante diferentes condiciones de estrés abiótico, como las temperaturas altas y bajas (5), sequía y salinidad (6). El inconveniente para su aplicación es su costosa obtención.
En Cuba existen los análogos espirostánicos de brasinoesteroides, obtenidos químicamente por el Centro de Estudios de Productos Naturales de la Facultad de Química de la Universidad de La Habana. Estas moléculas también han demostrado su efecto protector ante el déficit hídrico, la salinidad y las temperaturas altas (7, 8, 9). Sin embargo, muy pocos trabajos los vinculan a la salinidad en los estadios tempranos de las plantas.
El objetivo de este trabajo fue evaluar el efecto de una formulación basada en un análogo espirostánico de brasinoesteroide, conocida como Biobras-16 (BB-16), en la germinación y el crecimiento inicial de dos variedades cubanas de tomate, Amalia e INCA 9(1), que han demostrado tener diferente sensibilidad a la salinidad.
MATERIALES Y MÉTODOS
Primeramente, se realizó un ensayo para seleccionar el tiempo de imbibición de las semillas en agua y las condiciones de salinidad que afectan la germinación y el crecimiento de las variedades de tomate (Solanum lycopersicum) Amalia (10) e INCA 9(1) (11). Para ello, las semillas se esterilizaron con lejía comercial por 30 segundos, luego se lavaron varias veces con agua destilada. Posteriormente, se imbibieron con agua destilada durante 0, 4 y 8 h a 25°C y se colocaron en placas Petri, a razón de 25 semillas/placa y tres placas/tratamiento, con diferentes concentraciones de NaCl (50, 75 y 100 mM) durante siete días en condiciones de oscuridad. Diariamente se determinó la dinámica de germinación y como criterio se tomó la emergencia de la radícula. A los siete días posteriores a la germinación, se evaluaron las longitudes del tallo y la raíz, así como las masas fresca y seca de las plántulas. Con los datos se hizo un análisis de varianza de clasificación simple. En caso de diferencias significativas, la comparación de medias se realizó por los intervalos de confianza mediante la prueba de Bonferroni con una probabilidad del 95 %.
En un segundo experimento se evaluó el efecto de una formulación, cuyo ingrediente activo es un análogo espirostánico de brasinoesteroide de fórmula global C27H42O5, conocida como Biobras-16 (BB-16), a diferentes concentraciones (10-10-10-6mol.L-1).
Esta formulación se obtuvo en el Centro de Estudios de Productos Naturales de la Facultad de Química de la Universidad de La Habana. Para realizar el experimento, se esterilizaron las semillas de manera similar a la descrita anteriormente y luego se mantuvieron en agua destilada durante cuatro horas a 25°C. Las semillas se colocaron, igualmente, en placas Petri con agua destilada o una solución de 75 mM de NaCl, suplementadas o no con las distintas concentraciones de BB-16 y se mantuvieron durante siete días en condiciones de oscuridad a 25ºC. Se realizaron las mismas determinaciones y el tratamiento estadístico fue similar. Cada experimento se realizó por duplicado.
RESULTADOS Y DISCUSIÓN
En la Figura 1 se muestran
los resultados del porcentaje de germinación de las semillas a las 48 h (Figura 1A) y 72 h
(Figura 1B) del primer
ensayo realizado, para la selección del tiempo de imbibición y la concentración de NaCl.
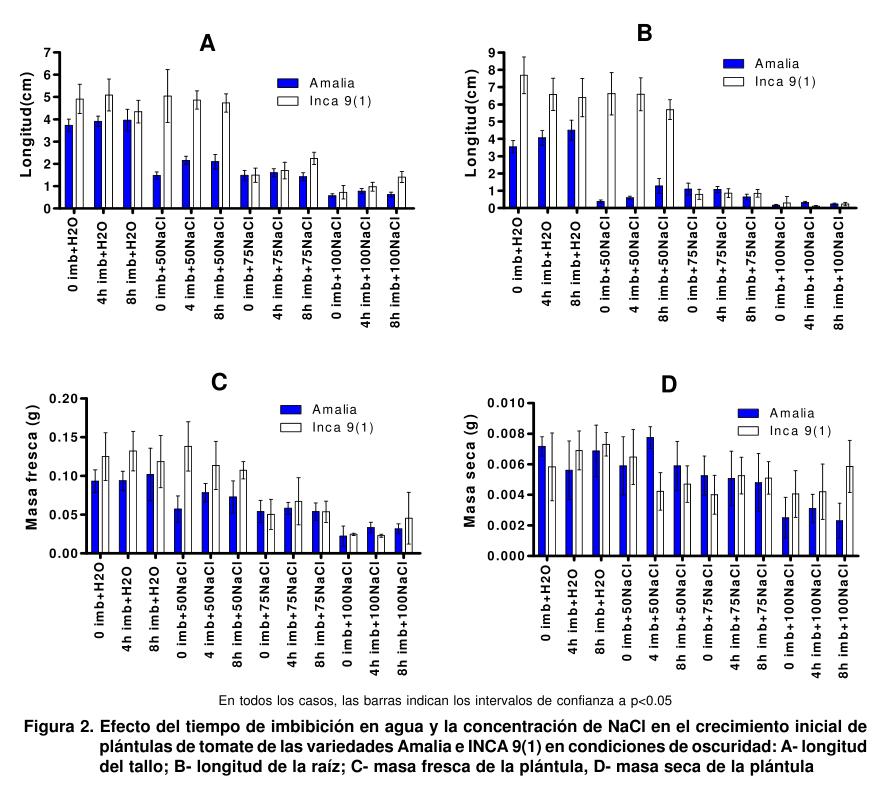
Como se puede apreciar, la imbibición de las semillas en agua destilada aumentó el porcentaje de germinación en la variedad INCA 9(1) en los controles, aunque en las diferentes condiciones de salinidad se evidenció en menor medida. No obstante, se ha planteado que uno de los indicadores más tolerantes a las condiciones de salinidad es la absorción de agua por las semillas, supuestamente debido a que en esta etapa operan procesos físico-químicos, como la imbibición del epiblasto, más resistentes al estrés (12). La imbibición en agua en la variedad Amalia favoreció la germinación cuando la concentración de NaCl fue de 75 y 100 mM.
De forma general, la imbibición en agua destilada por ocho horas incrementó ligeramente el porcentaje de germinación con respecto a la imbibición por cuatro horas; sin embargo, la primera introdujo mayor variabilidad en algunas de las otras variables estudiadas (Figura
2). Por esta razón, se decidió escoger cuatro horas de imbibición para continuar los experimentos.
Además, en la Figura
1 se muestra una disminución del porcentaje de germinación según aumenta la concentración de NaCl, siendo la variedad INCA 9(1) la que más retarda su germinación con el estrés salino. Se ha demostrado que la INCA 9(1), al inicio del estrés salino, es más sensible que la Amalia, probablemente debido a una mayor afectación por el estrés hídrico, aunque posteriormente aumenta su tolerancia a la salinidad con respecto a Amalia, por su mayor capacidad para excluir los iones cloruro (13). El porcentaje final de germinación se comportó de igual forma en las dos variedades para las concentraciones de NaCl estudiadas.
En la Figura 2 se muestran
las variables de crecimiento de ambas variedades. Se observa que en la INCA 9(1), a la concentración de 50 mM de NaCl, se obtuvieron valores similares a los controles en todas las variables estudiadas. Estos resultados concuerdan con los que demuestran que en condiciones de salinidad moderada (50 mM NaCl), la INCA 9(1) presenta un crecimiento similar a los controles, mientras que en la Amalia se observó una reducción significativa de la longitud del tallo y las raíces, lo que pudiera estar asociado a la mayor capacidad de la INCA 9(1) para excluir iones Cl- como mecanismo de tolerancia (13).
Las concentraciones de 75 y 100 mM afectaron sensiblemente el crecimiento de las dos variedades, siendo más notable a la de 100 mM. Se seleccionó la de 75 mM para realizar los experimentos con el análogo de brasinoesteroide BB-16, pues la reducción en las variables de crecimiento fue similar en ambas variedades y así se podría evaluar si la respuesta de ellas era diferente o no en presencia del análogo de BR.
En la Figura 3 se muestra
la dinámica de germinación de las semillas de las dos variedades con las distintas concentraciones de BB-16. Se observa que a las 48 h, el BB-16 provocó un aumento significativo en la velocidad de germinación de las semillas de la variedad INCA 9(1) en condiciones salinas, especialmente en la concentración de 10-7 mol.L-1, la cual fue capaz de igualarse al control sin NaCl; sin embargo, los porcentajes finales de germinación fueron similares entre todos los tratamientos. Está bien establecido que los brasinoesteroides promueven la germinación de las semillas, particularmente en tomate, estimulando la germinación por la vía de la expansión del embrión (14).
De forma similar, se ha encontrado que los brasinoesteroides naturales, 24-epibrasinólida (24-EBL) y 28-homobrasinólida (28-HBL), mejoraron la germinación a las 48 h y el crecimiento de las plántulas de arroz en condiciones salinas (15).
En experimentos realizados con el análogo de brasinoesteroide Biobras 6 (BB-6) para la germinación de la variedad Campbell-28 en condiciones salinas, no se modificó la dinámica de germinación, aunque se encontraron ligeros aumentos en los porcentajes finales de germinación. Se encontró, también, que la menor concentración del análogo (0.001 mg.L-1) estimuló las masas fresca y seca de las radículas a las concentraciones de 50 y 100 mM de NaCl (16).
Es curioso observar cómo el BB-16 a la concentración de 10-6 mol.L-1 provocó un retraso en la velocidad de germinación en las dos variedades estudiadas, siendo más notable en la INCA 9(1). Este efecto inhibitorio de los brasinoesteroides a concentraciones por encima de la óptima se encuentra ampliamente informado en la literatura. La administración de cantidades excesivas de brasinólida y castasterona provoca el crecimiento anormal de los órganos vegetales, como la inflamación y distorsión de los hipocótilos y raíces, tanto a la luz como en la oscuridad (17).
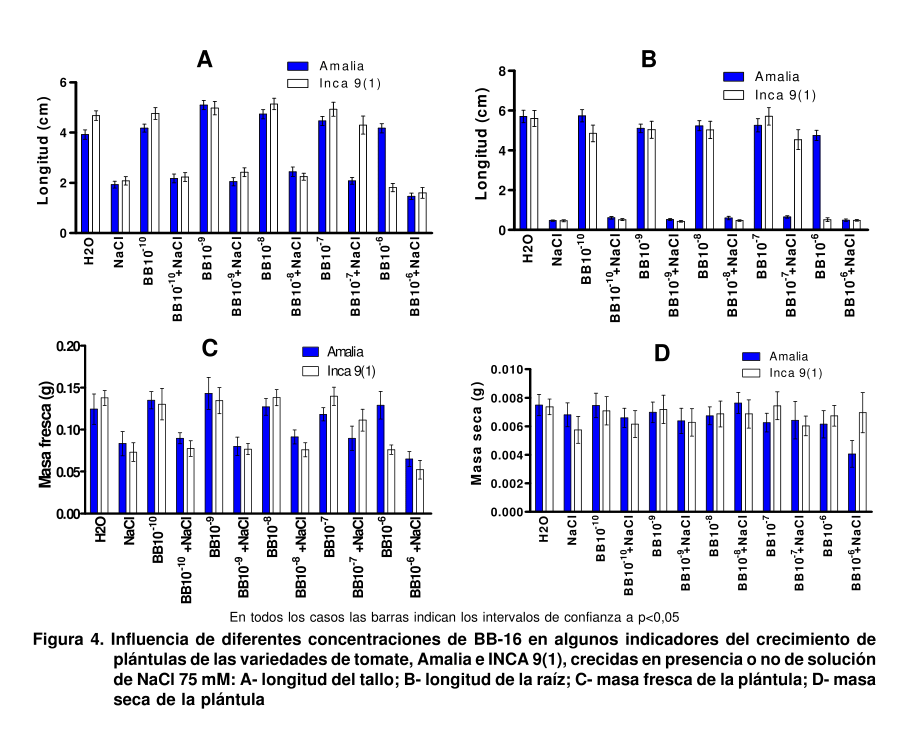
En la Figura 4 se muestran
las variables de crecimiento de las dos variedades estudiadas. Se observa que la concentración de 10-7 mol.L-1 de BB-16 fue capaz de revertir totalmente los efectos inhibitorios de la salinidad en la longitud del tallo (4A) y las raíces (4B), así como en la masa fresca (4C) de las plántulas de la INCA 9(1). Sin embargo, esto no se puso de manifiesto en la masa seca (4D), probablemente por un posible papel regulatorio de los BR en el contenido de agua de la célula. En la variedad Amalia no se observó efecto del análogo en las variables estudiadas.
Está bien demostrado que las respuestas inducidas por brasinoesteroides requieren síntesis de novo de proteínas y, por tanto, inducen síntesis de ARNm. Uno de los genes inducidos por BR es BRU1, aislado de frijol de soya, que codifica para una xiloglucano endotransglicosilasa (XET) (18). Posteriormente, se han encontrado estas XET inducidas por brasinoesteroides en otras especies vegetales como Arabidopsis, arroz y tomate, demostrando que la inducción de XET correlaciona con el ablandamiento de la pared celular durante la elongación celular inducida por BR (19). Otro de los genes inducidos por brasinoesteroides es CycD3, quesugiere que los BR pudieran tener un posible papel en la regulación del ciclo celular (20). También se ha observado la inducción de proteínas del citoesqueleto como la actina, α- y β-tubulina por los BR, lo cual pudiera tener un papel importante en la reorganización de los microtúbulos durante la división celular.
En general, los niveles de brasinoesteroides, como la castasterona (CS), son más bajos en las raíces que en los meristemos y, al parecer, los requerimientos en este órgano también son menores (21). Los primeros informes mostraron que la 24-EBL tenía efectos inhibitorios en la formación de las raíces en maíz, tomate, Arabidopsis y plántulas de soya (22). Sin embargo, otros han informado incrementos en la longitud de la raíz de plantas de berro (Lepidium sativum) crecidas en la oscuridad y las raíces primarias de maíz (23). Algunos han demostrado que esta respuesta es dependiente de la concentración, pues las bajas concentraciones de BR externos (0.1 nM de EBL y 10 nM de EpiCS) estimularon el crecimiento de la raíz de plantas de genotipos silvestres y restablecieron los valores normales en los mutantes deficientes de brasinoesteroides. Las altas concentraciones de estos compuestos resultaron inhibitorias (10 nM de EBL y 500 nM de EpiCS), probablemente por la estimulación de la biosíntesis de etileno, el cual se conoce que inhibe la elongación de las raíces (24).
El aumento de la masa fresca, no de la masa seca de las plántulas, en la INCA 9(1) podría sugerir un posible papel de los brasinoesteroides en la regulación del contenido de agua en la célula, aún más cuando se ha observado un incremento de la actividad de las aquaporinas por los brasinoesteroides (25), lo cual podría conllevar a una recuperación de la turgencia y expansión celular.
El hecho de que en la variedad Amalia no se observó ningún efecto del análogo en las variables estudiadas, pero sí en la INCA 9(1), indica la importancia de la sensibilidad de la variedad al estrés para la respuesta a la aplicación de brasinoesteroides, aún más cuando la Amalia tampoco respondió al BB-16 cuando la concentración de NaCl fue de 50 mM (datos no mostrados).
Este mismo efecto se observó en trigo, donde el tratamiento con 24-EBL fue capaz de mejorar la pérdida de biomasa solo en la variedad tolerante a la salinidad (26).
Existen pocos informes sobre los mecanismos de tolerancia a la salinidad en tomate y menos aún están dilucidados cómo los BR incrementan la tolerancia a la salinidad, particularmente en las variedades menos susceptibles.
La salinidad afecta el metabolismo de la pared celular vegetal por cambios en su elasticidad y composición (27). Se ha demostrado que algunos genes involucrados en el metabolismo de la pared celular son altamente modulados por el estrés salino, particularmente en tomate; entre ellos se encuentran: DY523676, que codifica para una xiloglucano endotransglicosilasa hidrolasa (XTH), DY523636, una glucano endo-1,3-β-glucosidasa y cinco genes que codifican para proteínas arabinogalactano (AGP).
Estos genes fueron severamente reprimidos por el estrés salino, tanto en un genotipo tolerante como susceptible, excepto XTH y una de las AGP que solo fueron reprimidas en el genotipo susceptible (28). XTH corta y libera los polímeros de xiloglucano, un constituyente esencial de la pared celular primaria y, por tanto, participa en la elongación y estructuración de las paredes celulares (29). AGP son componentes de las paredes celulares y membranas plasmáticas, y se considera que juegan un papel esencial en la diferenciación celular, el desarrollo y la interacción célula-célula, germinación y crecimiento de plántulas de tomate (30). Se conoce el papel de los BR para inducir genes que codifican para enzimas relacionadas con el ablandamiento de la pared celular y regulación del ciclo celular, por lo que este podría ser un mecanismo mediante el cual estas hormonas esteroidales disminuyen el efecto del estrés.
Otros genes afectados por la salinidad en tomate son los involucrados en la reducción y fijación del nitrógeno, así como en la biosíntesis de metionina. Por ejemplo, existe información donde se plantea que la nitrato y nitrito reductasa y la glutamina sintetasa han sido reguladas negativamente ante el estrés salino (31). Además, tres genes involucrados en la biosíntesis de metionina (S-adenosilmetionina sintasa, S-adenosil -L- metionina sintetasa y metionina sintasa) fueron regulados negativamente en una variedad tolerante y una susceptible, aunque en la tolerante esta represión fue más lenta. La asociación entre la metionina y la tolerancia a la salinidad se descubrió en la levadura, donde se evidenció que el suplemento con este aminoácido mejoró la tolerancia al NaCl (28). Recientemente, otros encontraron que algunas de las enzimas involucradas en el ciclo de la S-adenosil-L-metionina, como: S-adenosil-L-metionina: carboxil metiltransferasa, S-adenosilmetionina sintetasa, y S-adenosil-L-homocisteína hidrolasa se incrementaban con los BR (19). Este pudiera sugerir otro posible mecanismo de los BR para aumentar la tolerancia a este estrés.
Sin duda, mucho queda por investigar todavía sobre los efectos de los BR en la disminución del estrés abiótico, en general, por lo que los esfuerzos futuros deben concentrarse en dilucidar los mecanismos fisiológicos y moleculares, por los cuales estos compuestos inducen tolerancia a la salinidad, particularmente en las variedades menos susceptibles.
REFERENCIAS
- FAO. Land and Plant Nutrition Management Service. http://www.fao.org/ag/agl/agll/spush. 2008.
- Lau, A., Garea, E., Ruiz, M. E. Estimación de la salinidad de los suelos utilizando una imagen espectrozonal y el sistema de información geográfica TELEMAP. Revista Ciencias Técnicas Agropecuarias, 2005, vol. 14, no. 1, p. 47-54.
- Camejo, D. y Torres, W. La salinidad y su efecto en los estadios iniciales del desarrollo de dos cultivares de tomate (Lycopersicon esculentum, Mill). Cultivos Tropicales, 2000, vol. 21, no. 2, p. 23-26.
- Bajguz, A. y Hayat, S. Effects of brassinosteroids on the plant responses to environmental stresses. Plant Physiology and Biochemistry, 2009, vol. 47, p. 1-8.
- Singh, I., Shono, M.Physiological and molecular effects of 24-epibrassinolide, a brassinosteroid on thermotolerance of tomato. Plant Growth Regulation, 2005, vol. 47, p. 111-119.
- Kagale, S., Divi, U. K., Krochko, J. E., Keller, W. A., Krishna, P. Brassinosteroid confers tolerance in Arabidopsis thaliana and Brassica napus to a range of abiotic stresses. Planta, 2007, vol. 225, p. 353-364.
- García, A.; Rodríguez, T.; Héctor, E. y Núñez, M. Efecto del análogo de brasinoesteroide MH-5 en el crecimiento del arroz (Oryza sativa L.) en condiciones de déficit hídrico. Cultivos Tropicales, 2005, vol. 26, p. 87-91.
- Núñez, M.; Mazorra, L. M.; Martínez, L.; González, M. C. y Robaina, C. Análogos de brasinoesteroides revierten parcialmente el impacto del estrés salino en el crecimiento inicial de las plántulas de dos genotipos de arroz (Oryza sativa L.). Cultivos Tropicales, 2007, vol. 28, no. 2, p. 95-99.
- Mazorra, L. M. Inducción de respuestas antioxidantes y tolerancia al choque térmico por los brasinoesteroides y sus análogos en plántulas de tomate. [Tesis de Maestría en Biología Vegetal, Mención Fisiología Vegetal] La Habana : Universidad de La Habana. 2004.
- Alvárez, M.; Armas, G. de y Martínez, B. Amalia y Mariela, dos nuevas variedades de tomate para consumo fresco. Cultivos Tropicales, 1997, vol. 18, no. 1, p. 83.
- González, M. C. INCA 9-1, nueva variedad de tomate para diferentes épocas de siembra. Cultivos Tropicales, 1997, vol. 18, no. 1, p. 82.
- González, L. M. Apuntes sobre la fisiología de las plantas cultivadas bajo estrés de salinidad. Cultivos Tropicales, 2001, vol. 23, no. 4, p. 47-57.
- Dell’Amico, J. M. y Parra, M. Efecto del estrés por NaCl en el contenido de cloruros, el potencial osmótico real y el crecimiento de dos cultivares de tomate cubanos. Cultivos Tropicales, 2005, vol. 26, no. 2, p. 39-44.
- Vert, G.; Nemhauser, J. L.; Geldner, N.; Hong, F. X. y Chory, J. Molecular mechanisms of steroid hormone signalling in plants. Annu. Rev. Cell Dev. Biol., 2005, vol. 21, p. 177-201.
- Anuradha, S. y Rao, S. Effect of brassinosteroids on salinity stress induced inhibition of seed germination and seedling growth of rice (Oryza sativa L.). Plant Growth Regul., 2001, vol. 33, p. 151-153.
- Nuñez, M., Análogos de brasinoesteroides cubanos como biorreguladores en la agricultura. [Informe final de Proyecto de Investigacion. PNCT. Biotecnología Agrícola. Código 00300047]. La Habana : INCA, junio 2000.
- Tanaka, K.; Nakamura, Y.; Asami, T.; Yoshida, S.; Matsuo, T. y Okamoto, S., Physiological roles of brassinosteroids in early growth of Arabidopsis: Brassinosteroids have a synergistic relationship with gibberellin as well as auxin in light-grown hypocotyl elongation. J. Plant Growth Regul., 2003, vol. 22, p. 259-271.
- Zurek, D. M. y Clouse, S. D. Molecular cloning and characterization of a brassinosteroid regulated gene from elongating soybean (Glycine max L.) epicotyls. Plant Physiol., 1994, vol. 104, p. 161-170.
- Deng, Z.; Zhang, X.; Tang, W.; Oses-Prieto, J. A.; Suzuki, N.; Gendron, J. M.; Chen, H.; Guan, S.; Chalkley, R. J.; Peterman, T. K.; Burlingame, A. L. y Wang, Z. Y. A proteomics study of Brassinosteroid response in Arabidopsis. Molecular & Cellular Proteomics, 2007, vol. 6, p. 2058-2071.
- Hu, Y. X.; Bao, F. y Li, J. Y. Promotive effect of brassinosteroids on cell division involves a distinct CycD3-induction pathway in Arabidopsis. Plant J., 2000, vol. 24, p. 693-701.
- Shimada, Y.; Goda, H.; Nakamura, A.; Takatsuto, S.; Fujioka, S. y Yoshida, S., Organ-specific expression of brassinosteroid-biosynthetic genes and distribution of endogenous brassinosteroids in Arabidopsis. Plant Physiol., 2003, vol. 131, p. 287-297.
- Sasse, J. M. Physiological actions of brassinosteroids: an update. J. Plant Growth Regul., 2003, vol. 22, p. 276-288.
- Kim, S. K.; Chang, S. C.; Lee, E. J.; Chung, W. S.; Kim, Y. S.; Hwang, S. y Lee, J. S. Involvement of brassinosteroids in the gravitropic response of primary root of maize. Plant Physiol., 2000, vol. 123, p. 997-1004.
- Mussig, C.; Shin, G. H. y Altmann, T. Brassinosteroids promote root growth in Arabidopsis. Plant Physiol., 2003, vol. 133, p. 1261-1271.
- Morillon, R.; Catterou, M.; Sangwan, R. S.; Sangwan, B. S. y Lassalles, J. P. Brassinolide may control aquaporin activities in Arabidopsis thaliana. Planta, 2001, vol. 212, p. 199-204.
- Shahbaz, M.; Ashraf, M. y Athar, H. R. Does exogenous application of 24-epibrassinolide ameliorate salt induced growth inhibition in wheat (Triticum aestivum L.)? Plant Growth Regul., 2008, vol. 55, p. 51-64.
- Munns, R. y Tester, M., Mechanisms of salinity tolerance. Annu. Rev. Plant Biol., 2008, vol. 59, p. 651-681.
- Ouyang, B.; Yang, T.; Li, H.; Zhang, L.; Zhang, Y.; Zhang, J.; Fei, Z. y Ye, Z., Identification of early salt stress response genes in tomato root by suppression subtractive hybridization and microarray analysis. Journal of Experimental Botany, 2007, vol. 58, no. 3, p. 507-520.
- Fonseca, S.; Monteiro, L.; Barreiro, M. G. y Pais, M. S.Expression of genes encoding cell wall modifying enzymes is induced by cold storage and reflects changes in pear fruit texture. Journal of Experimental Botany, 2005, vol. 56, p. 2029-2036.
- Lu, H.; Chen, M. y Showalter, A. M. Developmental expression and perturbation of arabinogalactan-proteins during seed germination and seedling growth in tomato. Physiologia Plantarum, 2001, vol. 112, p. 442-450.
- Debouba, M.; Maaroufi-Dghimi, H.; Suzuki, A.; Ghorbel, M. H. y Gouia, H. Changes in growth and activity of enzymes involved in nitrate reduction and ammonium assimilation in tomato seedlings in response to NaCl stress. Annals of Botany, 2007, vol. 99, p. 1143-1151.


{kind=link}
{kind=link}
{kind=link}
{kind=link}