










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCultivos Tropicales
versión impresa ISSN 0258-5936
cultrop vol.31 no.3 La Habana jul.-set. 2010
ESTUDIO DE LA ASOCIACIÓN Gluconacetobacter diazotrophicus-VIANDAS TROPICALES EN SUELO FERRALÍTICO ROJO. I. SELECCIÓN DE CEPAS EFECTIVAS PARA LA BIOFERTILIZACIÓN DE BONIATO, YUCA Y MALANGA
Association study Gluconacetobacter diazotrophicus-Vianden TROPICAL RED SOIL Ferralitic. I. EFFECTIVE FOR SELECTION OF STRAINS OF SWEET POTATO biofertilization, Cassava and taro
Dr.C. B. DibutI. E-mail: bdibut@inifat.co.cu y R. MartínezI, Yoania RíosII, Liuba PlanaII y Janet RodríguezII, Marisel OrtegaIII y Grisel TejadaIV
I Investigadores Titulares
II Investigadoras Agregadas
III Investigadora
IV Investigadora Auxiliar del Instituto de Investigaciones Fundamentales en Agricultura Tropical (INIFAT), calle 2 esq. a 1, Santiago de las Vegas, Ciudad de La Habana, Cuba, CP 17200.
RESUMEN
El interés de la comunidad científica internacional por la bacteria endófita Gluconacetobacter diazotrophicus, microorganismo fijador de nitrógeno atmosférico y productor de sustancias promotoras del crecimiento vegetal, se ha incrementado en los últimos años. Por lo que teniendo en cuenta su potencial agrobiológico y los escasos estudios sobre el efecto que puede provocar la aplicación de productos elaborados a partir de la fermentación de cepas efectivas, se diseñó esta investigación, con el objetivo de seleccionar una cepa de Gluconacetobacter diazotrophicus efectiva para estimular el crecimiento, desarrollo y rendimiento de viandas tropicales, como son el boniato (Ipomea batatas Lam.), la yuca (Manihot esculenta, Crantz) y la malanga (Xanthosoma spp.), cultivados sobre suelo un Ferralítico Rojo. La cepa INIFAT Abn1, aislada del cultivo del boniato, fue la más promisoria para incrementar los diferentes indicadores en los tres cultivos, con incrementos en valores de estimulación entre 23 y 46 %. Se demostró la importancia de contar con una colección de amplia diversidad para realizar una selección adecuada, así como la no existencia de una relación directa cultivo de procedencia-efectividad de la cepa.
Palabras clave: Gluconacetobacter diazotrophicus, viandas tropicales, cepas, selección.
ABSTRACT
In recent years, the international scientific community has an increasing interest for the endophytic bacterium Gluconacetobacter diazotrophicus, an atmospheric nitrogen-fixing microorganism, which is also a producer of crop growth-promoting substances. Therefore, taking into account its agrobiologic potential and the scarce studies about the effect that may provoke the application of products manufactured through the effective strain fermentation, this investigation was laid out, with the aim of selecting an effective strain of Gluconacetobacter diazotrophicus to promote tropical vegetable growth, development and yield, such as sweet potato (Ipomea batatas Lam.), cassava (Manihot esculenta, Crantz) and taro (Xanthosoma spp.), cultivated on a Red Ferralitic soil. INIFAT Abn1, isolated from sweet potato crop, was the most promising strain to increase all indicators in the three crops, with higher promoting values between 23 and 46 %. The significance of having a wide diversity collection to perform an adequate selection was proved, as well as the non-existence of a direct relationship of crop origin-strain effectiveness.
Key words: Gluconacetobacter diazotrophicus, tropical vegetables, strains, selection.
INTRODUCCIÓN
Los biofertilizantes más antiguos y extendidos a nivel mundial se elaboran a base de las bacterias Rhizobium y Bradyrhizobium. Estos se utilizan para mejorar la producción y el contenido de proteínas en los cultivos de leguminosas para granos y aceites, debido a la alta eficiencia que desarrollan en su asociación con las plantas, fundamentalmente por el suministro de nitrógeno a partir de la fijación simbiótica (1, 2). En este sistema se ha insistido mucho en los estudios de selección de las cepas más efectivas en su interacción con el material genético vegetal, a diferencia de lo que ocurre en otros menos eficientes, como es el caso de las rizobacterias que se asocian con las plantas cultivables (3, 4).
Para las asociaciones endófitas el tema ha sido menos abordado aún (4, 5), ya que no ha despertado a escala mundial el interés agrobiológico aplicado de estas asociaciones. Así, se han realizado investigaciones sobre las bacterias Herbaspirillum sp. y Gluconacetobacter diazotrophicus, en función de su capacidad de asociarse con las plantas y su efecto nitrofijador (3, 6, 7), pero no acerca del efecto de estos microorganismos en el rendimiento de los cultivos a escala de extensión agrícola; de aquí que no abunden las investigaciones sobre los procedimientos de selección de cepas efectivas en su interacción con determinadas especies cultivables.
Gluconacetobacter diazotrophicus, microorganismo aislado por primera vez en 1988 por Cavalcante y Döbereiner (5) a partir de raíces y tallos de caña de azúcar cultivada en distintas regiones del Brasil, ha sido aislado además de diferentes variedades de ese cultivo en otros países, como Australia, México, India y Canadá (8, 9), de hierba elefante, boniato, café, piña, maíz, millo africano, así como de Saccharococcus sacchari, plaga común de la caña de azúcar y del interior de las esporas de hongos micorrizógenos arbusculares (5, 7, 10, 11).
En Cuba, se ha estudiado el microorganismo en el interior de la caña de azúcar, lográndose obtener un número importante de aislados, así como el conocimiento de su actividad nitrofijadora y estimuladora del crecimiento vegetal, y la relación con otros microorganismos asociados a este cultivo (4, 12). A pesar de todos los esfuerzos realizados (3, 6, 13), no abundan referencias acerca del interés agrobiológico que como bacteria endófita presenta este microorganismo sobre el metabolismo vegetal y, por tanto, su actividad sobre el crecimiento, desarrollo y rendimiento de los cultivos.
En este trabajo se presentan los resultados que permiten seleccionar a las cepas eficientes de Gluconacetobacter diazotrophicus sobre el cultivo de yuca (Manihot esculenta Crantz), boniato (Ipomea batatas Lam.) y malanga (Xanthosoma spp.), a partir de un conjunto de aislados de la bacteria en diferentes especies vegetales de interés económico.
MATERIALES Y MÉTODOS
Características del suelo y material biológico empleado (cepas bacterianas). Los experimentos se realizaron sobre un suelo Ferralítico Rojo (14), en áreas agrícolas del Instituto de Investigaciones Fundamentales en Agricultura Tropical (INIFAT), Santiago de Las Vegas, entre 1999 y 2002 (Tabla I).

Se empleó un diseño experimental en bloques al azar con cuatro réplicas, un tamaño de parcela de 30m2 y al menos dos repeticiones de cada ensayo. Se realizaron todas las atenciones culturales de acuerdo con las recomendaciones de las normas técnicas de cada cultivo (15). Se evaluaron los indicadores siguientes: número de raíces y tubérculos por planta, diámetro y peso promedio de estos y rendimiento agrícola para medir la respuesta a la bacterización.
En la Tabla II se presentan los diferentes aislados obtenidos de la bacteria (4) a partir de ocho cultivos con una amplia diversidad genética.

En los experimentos para comprobar el efecto de la inoculación del endófito sobre las plantas desprovistas de la bacteria (vitroplantas), se determinó el largo de la raíz, diámetro del tallo, número de hojas, área foliar, número de tubérculos y rendimiento, tanto para las plantas de boniato inoculadas como las no inoculadas con Gluconacetobacter diazotrophicus.
Inoculación. Los inóculos de cada una de las ocho cepas de Gluconacetobacter diazotrophicus se obtuvieron en medio SG (13), en zaranda rotatoria regulada a 220 rpm de agitación y 32°C de temperatura. La aplicación de los biopreparados, a una concentración entre 3.2-3.5x1011 UFCxmL-1, se realizó con ayuda de una mochila, tratando en todo momento de abarcar toda la planta y la superficie de suelo que la rodea.
Las aplicaciones se realizaron en horas avanzadas de la tarde a razón de 2 L.ha-1 del bioproducto, que es la dosis recomendada en estas condiciones de crecimiento de la bacteria, en relación con la población que alcanza al ser aplicada (13).
Análisis biométrico. Los datos se analizaron mediante un modelo de Análisis de Varianza en Bloques al Azar para cada cultivo, aplicando en los casos de diferencias significativas para cada fuente de variación una prueba de comparación de medias Newman-Keuls al 5 % de significación. El procesamiento de toda la información se realizó mediante el programa Statistic versión 4.0.
RESULTADOS Y DISCUSIÓN
En la Tabla II se relacionan los aislados de la bacteria que se utilizaron en el esquema de selección de cepas sobre los cultivos de yuca, boniato y malanga. Obsérvese que se incluyen cepas provenientes de estos mismos cultivos, unidas a otras cinco estirpes aisladas de especies vegetales muy distantes, desde el punto de vista filogenético, en relación con las viandas tropicales, lo que permite enfrentar una relación cepa-cultivo basada en una amplia diversidad genética por parte del origen del microsimbionte.
Son escasos los estudios de este tipo con Gluconacetobacter diazotrophicus en asociación con especies cultivables de importancia económica. En la India y EE.UU. se ha logrado seleccionar cepas para caña de azúcar y maíz, aunque no se ha planteado aún como estrategia la inoculación de estos cultivos (2).
En Cuba se propone seleccionar la cepa más efectiva por cada cultivo, con el objetivo de dirigir un esquema de aplicación basado en la eficiencia de la interacción planta-microorganismo, capaz de poder ser introducido en áreas de viandas cultivadas, tanto en el sistema nacional de agricultura urbana como en la convencional.
En la Tabla III se evidencia la respuesta del boniato frente a la inoculación de las ocho cepas de la bacteria, resultando la INIFAT Abn1 la más efectiva, según los valores de rendimiento alcanzados.

Las cepas aisladas de yuca y malanga muestran un comportamiento favorable, pero las provenientes de papaya y remolacha no difieren estadísticamente de las estirpes obtenidas de las viandas tropicales. Esta respuesta corrobora el principio de dirigir esquemas de selección sobre la base de una amplia diversidad genética, para asegurar que el sistema cepa–cultivo resulte más efectivo en las condiciones edafoclimáticas así como el material genético vegetal que se presenta en la región de interés a estudiar.
En el caso de la interacción Gluconacetobacter diazotrophicus cepa INIFAT Az3-Ipomea batatas Lam., a pesar de ser la menos efectiva, logró promediar 1.34 t.ha-1 más que el control desprovisto del efecto del microorganismo, lo que pone de manifiesto que se puede mejorar la productividad, aún empleando una cepa poco efectiva del microorganismo que se asocia a este.De haberse contado solamente con este asilado, el incremento del rendimiento para el cultivo sería de 1.3 t.ha-1, en detrimento de las 8-12 t.ha-1 que se alcanzan con el empleo de las cepas más eficientes.
Otros dos aislados que se destacan son INIFAT AcL1 y Azn1, que provocan incrementos del rendimiento cercanos al 22 %, lo que equivale a volúmenes entre 6.08 y 6.19 t adicionales de tubérculos en las plantaciones sin tratar con la bacteria.
Cuando se emplean las cepas más efectivas, se obtienen producciones comerciales más altas, fenómeno que influye sobremanera en la formación del rendimiento, en función de la productividad neta del cultivo. En este sentido, se emplean múltiples reguladores del crecimiento, fundamentalmente de origen químico, que conducen, por lo general, a la alteración del agroecosistema (16), o por otra vía se realizan estudios de mejoramiento genético para cumplimentar este objetivo (17).
En los dos años de investigación siguientes, se obtuvo una respuesta similar, resultando nuevamente la cepa INIFAT Abn1 como la más efectiva (Tabla IV).
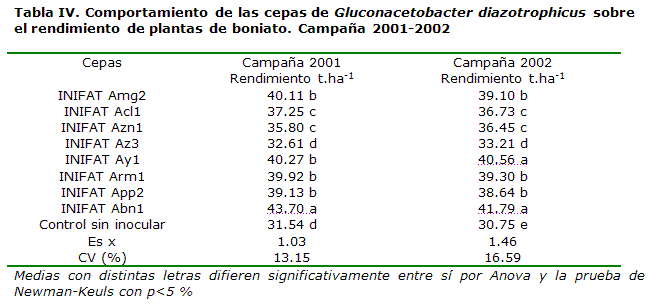
En estos ensayos, la interacción menos efectiva promedió entre 1.07 y 2.46 t.ha-1 tubérculos más respecto al control sin tratar y las más promisorias entre 7.59 y 11 t.ha-1.
Los valores de la estimulación del rendimiento por unidad de superficie se corresponden con los referidos por Paula (18), cuando inoculó plantas de boniato dispuestas en pequeñas parcelas con Gluconacetobacter diazotrophicus en asociación con los hongos micorrizógenos arbusculares (HMA). En estos estudios se inoculó igualmente con una cepa aislada y seleccionada a partir del mismo cultivo. Es sobre esta especie vegetal, junto a la caña de azúcar, donde más abundan los informes acerca del interés agronómico de la bacteria en asociación con las plantas superiores y, aún así, no se presentan esquemas de selección basados en la amplia diversidad genética por parte del endófito.
Como se puede observar en los tres años de experimentación, todas las cepas de Gluconacetobacter diazotrophicus probadas mostraron un comportamiento similar en cuanto al efecto estimulador, que permite sobrepasar el rendimiento del cultivo con respecto a la fracción testigo. En esta respuesta hay que considerar que, por lo general, está presente la bacteria de forma autóctona, ya que esta se transmite en la semilla agámica y, además, sus poblaciones en el tejido vegetal pueden estar estimuladas por la acción de los HMA, que actúan en el suelo como vía de penetración del microorganismo.
De esta forma, para demostrar el efecto que pudiera encontrarse en plantas desprovistas de Gluconacetobacter diazotrophicus, se desarrollaron experimentos a partir de material vegetal obtenido por cultivo in vitro de plántulas de boniato, a partir de yemas crecidas en un medio MS suplementado con ácido giberélico y citoquininas, a partir de la metodología desarrollada por Rodríguez*, como se observa en las Tablas V y VI.
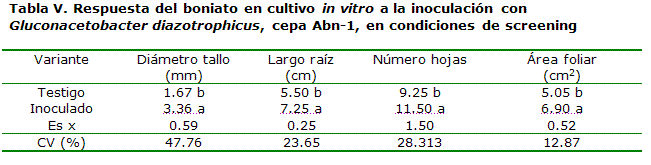
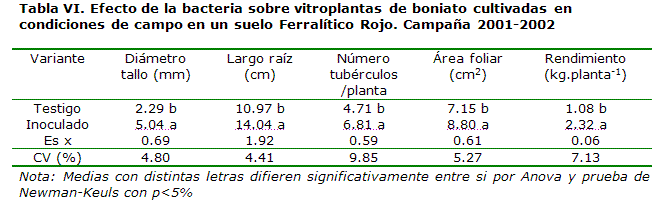
En una evaluación fenológica realizada en etapas intermedias del cultivo (40 días), el diámetro del tallo promedió más del doble en comparación con las plantas no inoculadas (Tabla V) y el resto de los indicadores (largo de la raíz, número de hojas y área foliar) mostró entre 37 y 50 % de aumento. En el momento final del ciclo de cultivo (Tabla VI), el diámetro del tallo de las plantas superó el 100 % de incremento y el número de tubérculos/planta fue de 2.1 frutos más, a favor de la inoculación, con un valor del rendimiento que promedió más del doble (1.24 kg.planta-1 de aumento).
Como se puede comprobar, esta respuesta es muy superior a la encontrada en este mismo cultivo empleando el mismo clon, cuando se condujeron los experimentos de selección de cepas, por lo que se deduce que la respuesta a la bacterización de este o cualquier otro cultivo es muy alta, cuando no ha sido colonizado de forma autóctona en su normal crecimiento dentro del agroecosistema donde habita, lo cual se explica por la competencia entre especies microbianas por el nicho ecológico que las posee, el contenido de nutrientes en esta zona y los mecanismos de antibiosis, biocontrol y predación que normalmente suceden en la rizosfera, entre otros factores; ya que en este caso, las raíces de las plántulas vienen desprovistas de poblaciones microbianas, a diferencia de las plantas que normalmente germinan a partir de la semilla o se plantan mediante semilla agámica, donde en ambos casos ya existe una colonización inicial en los bancos de semilla y cepellones.
Un comportamiento similar ha sido detectado en plantas de maíz y caña de azúcar (8, 19), donde se ha puesto de manifiesto la alta capacidad nitrofijadora y estimuladora de Gluconacetobacter diazotrophicus. De aquí la insistencia en recomendar la inoculación de los bancos de semillas agámicas, obtenidas por la vía biotecnológica del cultivo de tejidos vegetales, como es el caso de la reproducción de caña de azúcar en las condiciones de Cuba, la malanga, la piña y la papa, cultivos donde se ha demostrado la presencia o el efecto beneficioso del endófito, sin excluir el banano, la fresa, entre otros, donde los microbiólogos deberán estudiar la presencia de esta u otra bacteria beneficiosa asociada, que normalmente se pierde en la manipulación del tejido vegetal y que, sin dudas, ha de representar una ventaja evolutiva para la especie botánica que se explota agronómicamente.
En el cultivo de la yuca clon CMC-40, igualmente se detecta una respuesta favorable frente a todas las cepas probadas, destacándose INIFAT Abn1 como la estirpe más promisoria, la cual aumentó en relación con el control en 43 % el rendimiento agrícola y en 32 y 39 % el número y el diámetro de las raíces, respectivamente (Tabla VII).
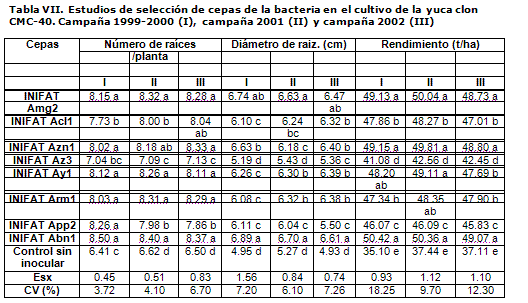
INIFAT Ay1, procedente originalmente del cultivo de la yuca, demostró un buen comportamiento, pero fue superada por las cepas INIFAT Amg2 y Azn1, obtenidas de malanga y zanahoria respectivamente, por lo que queda demostrado que no necesariamente debe existir relación entre el origen o procedencia de la bacteria y el cultivo al que beneficia. También se destacan por su efecto las cepas INIFAT Acl1 (calabaza) y Arm1 (remolacha). Coincide el comportamiento manifestado por la cepa INIFAT Az3 (maíz) con los resultados en boniato.
Los valores alcanzados durante el segundo y tercer años de experimentación fueron ligeramente superiores, lo que coincide con el comportamiento que presentó el boniato y ratifica la explicación dada para este cultivo.
Los resultados no pueden compararse con otros que refleja la literatura, ya que este cultivo no ha sido estudiado en su interacción con Gluconacetobacter diazotrophicus y los avances que aquí se presentan constituyen un primer informe. No obstante, el comportamiento detectado sí se corresponde con los efectos referidos en boniato (13, 19), especie que se ubica dentro del grupo de las viandas tropicales.
El cultivo de la yuca presenta un gran interés comercial y de base alimentaria para Centroamérica y África, por lo que sería muy conveniente introducir los resultados en estos mercados, con el objetivo de poder seleccionar cepas altamente efectivas, que permitan obtener rendimientos aceptables en condiciones de niveles nutricionales (reserva del suelo) e hídricos no elevados, características que imperan sobre todo en el continente africano.
Según la respuesta que es capaz de provocar la cepa aislada de maíz en estos cultivos, todo parece indicar que en la gramínea, al contener menores tenores de azúcar en el sistema planta, la bacteria no alcanza la misma eficiencia fisiológico-bioquímica que cuando proviene de la asociación de forma autóctona a partir otros cultivos, que se caracterizan por presentar altos tenores de estos carbohidratos como el boniato y la yuca.
En el tercer cultivo, la malanga, la cepa INIFAT Abn1 resultó la más efectiva (10 t.ha-1 de aumento en relación con el control) en su asociación con esta especie, desde el primer año de experimentación (Tabla VIII), solo seguida en su efecto mejorador sobre la productividad del cultivo por las cepas INIFAT Ay1 y Acl1 (9.61 y 9.02 t.ha-1). La cepa proveniente del cultivo de maíz mostró el menor índice de estimulación del rendimiento agrícola, aunque logró incrementar este indicador en 3.73 t.ha-1 en relación con el control desprovisto de la inoculación del microorganismo.
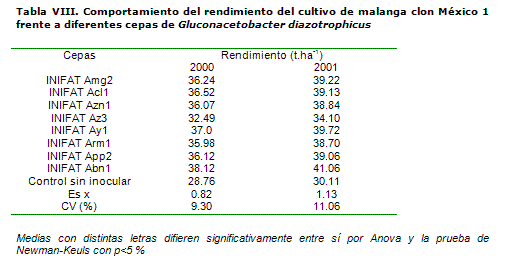
En estudios de biofertilización de este mismo cultivo, pero en este caso inoculando Azotobacter chroococcum, no se lograron respuestas tan elevadas en relación con las áreas sin tratar, en cuanto a la estimulación del rendimiento agrícola (20). En estas pruebas el aumento de la productividad de la malanga con respecto al control solo alcanzó un umbral entre 6.5 y 7.3 t.ha-1 con la obtención de tubérculos de mayor calidad.
La explicación de este comportamiento parece estar relacionada a la diferente capacidad de asociación con la planta que presentan estos dos microorganismos, donde sí se conoce que las bacterias endófitas poseen las mayores ventajas en el intercambio de nutrientes y la asimilación por parte de la planta del producto de su actividad metabólica (11).
Si se considera la problemática en cuanto al normal comportamiento que presenta el cultivo de la malanga en el país, cuyos rendimientos están alarmantemente degradados por diferentes factores (15), se puede aceptar hasta la idea de inocular aún con la cepa menos efectiva (INIFAT Az3) del endófito.
No se dispone de información acerca del efecto de la bacteria Gluconacetobacter diazotrophicus sobre la productividad de los tubérculos de malanga, ni aún en relación con la actividad del endófito sobre los parámetros de crecimiento de esta especie vegetal, por lo que estos resultados constituyen un primer informe sobre el tema; debido a ello, no se pueden comparar estos resultados con los aportados por la literatura especializada para esta especie cultivable; no obstante, el efecto agronómico sobre el rendimiento aquí encontrado con la cepa INIFAT Abn1 amerita tomar en consideración la estrategia de inocular, de inmediato, plantaciones de Xanthosoma spp., tanto en el marco de producción urbana y periurbana como rural.
Como se ha podido constatar, la cepa INIFAT Abn1 se ha comportado como la más efectiva en su asociación con los cultivos de boniato, yuca y malanga, por lo que esta estirpe ha de ser la recomendada para la elaboración del nuevo biofertilizante propuesto a base de Gluconacetobacter diazotrophicus, para el beneficio de estas tres importantes especies que inciden marcadamente en la dieta de nuestra población.
Por otra parte, se debe aclarar que los rendimientos aquí obtenidos en las tres especies de plantas estudiadas se corresponden con los que se refiere en la literatura (15), sin dejar de considerar que en los últimos años muchos de estos clones han sufrido una fuerte erosión, causada fundamentalmente por el deficiente manejo agronómico; de aquí que si se comparan los valores de productividad que actualmente se alcanzan en las viandas con los que inicialmente representaron las características o atributos de estos clones, por lo general resultan inferiores aunque con un impacto superior, ya que la microbiología aplicada busca mejorar la productividad vegetal a partir del potencial genético y el manejo agronómico adecuado de las variedades y clones de las más diversas especies de plantas.
Finalmente, como salida fundamental de este resultado se ofrece la obtención de la cepa INIFAT Abn1 del endófito, ubicada en el cepario de fijadores biológicos del nitrógeno (FBN) del Laboratorio de Microbiología del INIFAT, que logra estimular el crecimiento, desarrollo y rendimiento de los cultivos de boniato, yuca y malanga entre 27 y 46 % en relación a cuando no se procede a la inoculación del microsimbionte, con la obtención de notables beneficios socioeconómicos y ambientales que serán expuestos en trabajos posteriores de la serie.
REFERENCIAS
- Ríos, Y. Efecto de Gluconacetobacter diazotrophicus sobre el crecimiento, desarrollo y rendimiento del cultivo de la yuca (Manihot esculenta Crantz) var. CMC-40 (Tesis de Maestría). Universidad de La Habana. 2007, 57 p.
- Dibut, B. Biofertilizantes como insumos en Agricultura Sostenible. HUMIWORM SPR de RL México. 2005, 107 p.
- Oliveira, A. L. M., Canuto, E. D. L., Urquiaga, S., Reis, V. M. y Baldani, J. I. Yield of micropropagated sugarcane varieties in different soil types following inoculation with diazotrophic bacteria. Plant and Soil, 2006, vol. 284, p. 23-32.
- Ríos, Y. y Dibut, B. Gluconacetobacter diazotrophicus: un microorganismo promisorio en la elaboración de biopreparados. Cultivos Tropicales, 2007, vol. 28, no. 4, p. 19-24.
- Cavalcante, V. A. y Döbereiner, J. A new acid-tolerant nitrogen-fixing bacterium associated with sugarcane. Plant and Soil, 1988, vol. 108, no. 1, p. 23-31.
- Adriano-Anaya, M. L., Salador-Figueroa, M., Ocampo, J. A. y García-Romera, I. Hydrolytic enzyme activities in maize (Zea mays) and sorghum (Sorghum bicolor) roots inoculated with Gluconacetobacter diazotrophicus and Glomus intraradices. Soil Biol. Biochem., 2006, vol. 38, p. 879-886.
- Madhaiyan, M., Saravanan, V. S., Bhakiya-Silba-Sandal-Jovi, D., Lee, H., Thenmozhi, R., Haris, K y Sa, T. M. Ocurrence of Gluconacetobacter diazotrophicus in tropical and subtropical plants of Western Gahts, India. Microbiol Res., 2004, vol. 159, p. 233-243.
- Suman, A., Gaur, A., Shrivastava, A. K. y Yadav, R. L. Improving sugarcane growth and nutrient uptake by inoculating Gluconacetobacter diazotrophicus. Plant Growth Regul., 2005, vol. 47, p. 155-162.
- Medeiros, A. F. A., Polidoro, J. C. y Reis, V. M. Nitrogen source effect on Gluconacetobacter diazotrophicus colonization sugarcane (Saccharum spp). Plant and Soil, 2006, vol. 279, p. 141-152.
- Cocking, E. C., Stone, P. J, y Davey, M. R. Intracellular colonization of roots of Arabidopsis and crop plant by Gluconacetobacter diazotrophicus. In vitro Cell Dev Biol-Plant., 2006, vol. 42, p. 74-82.
- Carrizo-de-Bellone, S. y Bellone, C. H. Presence of endophytic diazotrophs in sugarcane juice. World J. Microbiol. Biotechnol., 2006, vol. 22, p. 1065-1068.
- Ortega, P.; Ortega, E.; Fernández, L. y Rodés, R. Composición de cinco aislados fijadores de nitrógeno obtenidos del interior de la caña de azúcar y de Saccharococcus sacchari. XIIICongreso Científico del INCA (2002, nov. 12–15, La Habana). Memorias CD-ROM Instituto Nacional de Ciencias Agrícolas, ISBN 959–7023229.
- Döbereiner, J.; Reis, V. M.; Paula, M. A. y Olivares, F. Endophytic diazotroph in sugarcane, cereals and tuber plants. En: New Horizons in Nitrogen Fixation. R. Palacio, J. Mora y W. E. Newton. Eds. Klumer Academic Publisher, Netherlands. 1993, p. 671–676.
- Cuba. Instituto de Suelos. Nueva versión de Clasificación Genética de los Suelos de Cuba, Ed. AGRINFOR, La Habana. 2000, 64 p.
- Cuba. MINAGRI. Evaluación de los resultados de 1999-Grupo de Cultivos Varios. Ciudad de La Habana. 2000, 17 p.
- Muthukumarasamy, R.; Govindarajan, M., Vadivelu, M. y Rebathi, G. N fertilizer saving by the inoculation of Gluconacetobacter diazotrophicus and Herbaspirillum sp. in micropropagated sugarcane. Plant Microbiol. Res., 2006, vol. 161, p. 238-245.
- Baldani, J. I. y Baldani, V. L. D. History on the biological fixation research in graminaceous plants: special emphasis on the Brazilian experience. An Acad. Bras. Cienc., 2005, vol. 77, p. 549-579.
- Paula, M. A., Urquiaga, S., Siqueira, J. O. y Döbereiner, J. Synergistic effects of vesicular-arbuscular mycorrhizal fungi and diazotrophic bacteria on nutrition and growth of sweet potato (Ipomea batatas). Biol. Fert. Soils, 1992, vol. 14, p. 61-66.
- Sevilla, M.; Burris, R. H.; Nirmala-Gunalapa, G. L.; Kennedy, C. y Gunalapa, N. Comparison of Benedit to sugarcane plant growth and N15 incorporation following inoculation of sterile plants with Acetobacter diazotrophicus wild-type and Nif-mutant strains. Molec. Plant. Micro. Interactions, 2001, vol. 14, no. 3, p. 358–366.
- Martínez, R., López, M., Dibut, B., Parra, C. y Rodríguez, J. La fijación biológica de nitrógeno atmosférico en condiciones tropicales. Venezuela. MPPAT. 2007, 172 p.
Recibido: 1 de octubre de 2009
Apreobado: 7 de abril de 2010

