










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCultivos Tropicales
versión impresa ISSN 0258-5936
cultrop vol.31 no.4 La Habana oct.-dic. 2010
EFECTO DEL ESTRÉS POR NaCl EN EL CRECIMIENTO Y LAS RELACIONES HÍDRICAS EN PLANTAS DE TOMATE (Solanum lycopersicum L.) DURANTE EL PERÍODO VEGETATIVO
STRESS EFFECT OF NaCl on growth and water relations in tomato (Solanum lycopersicum L.) during the growing
Dr.C. D. MoralesI. E-mail: dmorales@inca.edu.cu y Dr.C. J. MI. Dell'AmicoI, Dr.C. P. RodríguezII, Dr. A. TorrecillasIII y Dra. María de J. Sánchez-BlancoIV
I Investigadores Titulares
II Investigador Agregado del departamento de Fisiología y Bioquímica Vegetal, Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, La Habana, Cuba, CP 32700
III Profesor de Investigación
IV Colabora Científica del departamento de Riego y Salinidad, Centro de Edafología y Biología Aplicada del Segura (CEBAS), CSIC, Murcia, España
RESUMEN
Con el objetivo de determinar el comportamiento de plantas de tomate durante el período vegetativo frente a diferentes tratamientos salinos, se evaluaron el crecimiento y las relaciones hídricas en el cv. Amalia, sometido a distintos tratamientos con NaCl en el medio durante 264 h. El trabajo se realizó en una cámara de crecimiento con fotoperíodo de 13 h, una radiación fotosintéticamente activa de 380 µmol.m-2.s-1, temperatura día/noche de 25/180C y humedad relativa día/noche de 60/70 %. Las plantas se desarrollaron en macetas con arena sílice colocadas en bandejas, en las que se adicionó solución nutritiva de Hoagland. Los tratamientos utilizados fueron 0, 50, 100 y 200 mM de NaCl. Una vez concluida la fase de aplicación de los tratamientos, las plantas se colocaron en condiciones de solución nutritiva sin NaCl durante 264 h para evaluar su recuperación. Las evaluaciones de biomasa, conductancia estomática, transpiración, los potenciales hídrico foliar, osmótico, de presión y osmótico a máxima saturación antes del alba y la conductividad hidráulica de las raíces se realizaron a las 24, 48, 120 y 264 h de aplicados los tratamientos, así como a las 264 h de permanecer en condiciones de recuperación. El análisis de los datos reflejó una disminución significativa de las distintas variables evaluadas, al incrementarse el contenido de NaCl en el medio y se precisó que a las 24 h de aplicados los tratamientos, aún no se había producido ajuste osmótico.
Palabras clave: salinidad, relaciones hídricas, conductividad hidráulica, ajuste osmótico, tomate.
ABSTRACT
This study was conducted with the aim of determining tomato plant behavior during the vegetative period under different saline treatments; thus, growth and water relations were evaluated in cv. Amalia, subjected to several NaCl treatments in the medium for 264 h. It was performed in a growth chamber with a 13-h photoperiod, a photosynthetically active radiation of 380 µmol.m-2.s-1, a day/night temperature of 25/180C and a day/night relative humidity of 60/70 %. Plants developed in pots containing silica sand and put in trays, besides adding Hoagland nutrient solution. The treatments used were 0, 50, 100 and 200 mM NaCl. Once treatment application phase was concluded, plants were placed under nutrient solution conditions without NaCl for 264 h to evaluate their recovery. Evaluations of biomass, stomatal conductance, transpiration and leaf water, osmotic, pressure and osmotic at a top saturation potentials before dawn and root water conductivity were recorded after 24, 48, 120 and 264 h of applying treatments, as well as after staying for 264 h under recovery conditions. The analysis of data showed a significant reduction of different variables evaluated, when increasing NaCl content in the medium, besides observing that after 24 h of applying treatments, the osmotic adjustment had not yet occurred.
Key words: salinity, water relations, water conductivity, osmotic adjustment, tomato.
INTRODUCCIÓN
La salinidad es un estrés abiótico complejo, que simultáneamente presenta componentes osmóticos e iónicos (1). Por ello, una concentración elevada de sales en el medio radical afecta negativamente el desarrollo de la planta, debido fundamentalmente a los efectos hiperosmóticos e hiperiónicos del estrés (2).
La pérdida de la homeostasis, tanto hídrica como iónica, ocurre tanto a escala celular como a nivel de planta y provoca graves daños moleculares, que detienen el crecimiento de la planta, por una parte, la disminución del potencial hídrico del medio restringe la absorción de agua por las raíces y, por otra, la absorción de iones salinos específicos, que se acumulan en los tejidos de la planta en concentraciones que llegan a ser tóxicas, pueden al mismo tiempo inducir desequilibrios nutricionales por modificación de la absorción y distribución de nutrientes esenciales.
El estrés salino altera las relaciones hídricas de las plantas a través del estrés osmótico e hídrico; en respuesta a esto, las plantas desarrollan el ajuste osmótico y el déficit hídrico mantiene una suficiente turgencia para permitir el crecimiento, transporte, la acumulación y compartimentación de los iones inorgánicos y solutos orgánicos en las células de las plantas superiores (3).
El estudio de la tolerancia a la salinidad es en extremo complicado y el intento de solucionar, o al menos mitigar, los efectos perjudiciales de las sales incluye el estudio de cómo estas afectan los procesos fisiológicos, bioquímicos y moleculares del metabolismo de las plantas, así como de los diferentes mecanismos que las plantas ponen en marcha para defenderse del estrés (4).
Lo anteriormente expuesto conllevó a la realización de este trabajo, cuyo objetivo principal estuvo dirigido a determinar el efecto que la salinidad provocada por el NaCl ejerce en el crecimiento, la conductancia estomática, transpiración, conductividad hidráulica de las raíces y los potenciales hídrico, osmótico, de presión y osmótico a máxima saturación en plantas del cultivar cubano de tomate Amalia durante el período vegetativo.
MATERIALES Y MÉTODOS
Material vegetal y tratamientos. Las plantas de tomate (Solanum lycopersicum L. cv. Amalia) crecieron en macetas de 0.5 L con arena de sílice lavada, colocadas en bandejas que se introdujeron en una cámara de crecimiento con un fotoperíodo de 13 h, una radiación fotosintéticamente activa de 380 µmol.m-2.s-1, una temperatura día/noche de 25/180C y una humedad relativa día/noche de 60/70 %.
Todas las bandejas se mantuvieron en una solución nutritiva de Hoagland, a partir de las dos semanas posteriores a la siembra; cuando las plantas emitieron la octava hoja, se sometieron a los siguientes tratamientos: control en el que a la solución nutritiva no se le adicionó NaCl y tres tratamientos salinos, en los que la solución nutritiva contó con 50, 100 y 200 mM de NaCl durante 264 h.
Una vez transcurrido este período, todas las plantas se colocaron por un lapso de tiempo similar en solución nutritiva sin cloruro de sodio, con el objetivo de evaluar su posible recuperación.
El diseño utilizado fue el de bloques completamente aleatorizados con cuatro repeticiones.
Mediciones. Las masas secas de raíces, tallos y hojas se determinaron al comienzo y al final del período experimental.
La conductancia foliar en la superficie abaxial de la hoja (gl), el potencial (?l) antes del amanecer, el osmótico (?s), el de presión (?p), el osmótico a plena turgencia y la conductividad hidráulica de la raíz se midieron a las 24, 48, 120 y 264 h durante el período experimental y cuando finalizó el período de recuperación.
La conductancia foliar (gl) se midió con un porómetro de estado estacionario (LI-COR, Inc., Lincoln NE, USA). El potencial hídrico foliar (?l) se midió con una cámara de presión (Soil Moisture Equipment Co, Santa Barbara, CA).
Las hojas utilizadas para medir el ?l se congelaron en N líquido, después se descongelaron y se les determinó el potencial osmótico (?s) con el empleo de un osmómetro de presión de vapor (Wescor Inc. Logan, UT). El potencial de presión (??p) se estimó por la diferencia entre ?l y ?s.
El potencial osmótico a pleno turgor se midió de la misma forma que el osmótico, pero en este caso las hojas se saturaron en agua destilada durante 24 h antes de ser congeladas.
Para medir la conductividad hidráulica de las raíces, las plantas se cortaron al nivel del cuello de la raíz, después de extraídas de las macetas y lavado su sistema radical cuidadosamente. Posteriormente, se colocaron en un recipiente con solución nutritiva de Hoagland e introdujeron en la cámara de presión, quedando hacia el exterior una parte del tallo, en el que se ajustó un pequeño tramo de manguera tipo capilar. A partir de ese momento, se comenzó a incrementar la presión dentro de la cámara hasta que se obtuvo un flujo constante; desde entonces esta se fue incrementando a una taza de 0.4 MPa.min-1 hasta alcanzar una presión final de 1 MPa. A cada planta se le extrajeron tres exudados, midiéndose el volumen extraído cada 3 min. La conductividad hidráulica de la raíz se calculó usando la fórmula:
Lp= J/(P x L)
Donde:
Lp es expresada en mg.m-1.s-1.MPa-1, P es la presión hidrostática aplicada (MPa), L es el largo de la raíz (m) y J es la tasa de flujo de agua a través de todo el sistema radical (mg.s-1).
Las tasas relativas de crecimiento de la raíz, el tallo, las hojas y la planta (RGR) se estimaron usando la fórmula:
RGR= (Ln Md2-Ln Md1)???t
Donde:
RGR está expresado en g.d-1, Ln Md1 y Ln Md2 son el logaritmo neperiano de la masa seca (g) al comienzo y al finalizar el período experimental, y ?t? es el tiempo en días transcurridos entre el comienzo y final del estudio.
RESULTADOS Y DISCUSIÓN
La tasa relativa de crecimiento, al finalizar los períodos de estrés y recuperación (Figura 1), presentó un decrecimiento significativo, al aumentar la concentración salina en el medio, aunque las mayores diferencias se encontraron entre el tratamiento salino más elevado y los restantes. Además, se pudo observar que, en el caso de la raíz, aun cuando los valores obtenidos distan unos de otros, sobre todo entre los dos tratamientos extremos, los que contaron con niveles salinos intermedios entre el control y el de salinidad más elevada, las diferencias no fueron estadísticamente significativas respecto al control, pero sí lo fueron con el tratamiento más salinizado, comportamiento que puede deberse al deterioro que se produce en el sistema radical de las plantas expuestas a la sal, originando una gran heterogeneidad en ellas y, por consiguiente, un elevado error estándar, que hace que no se detecten esas diferencias desde el punto de vista matemático.

Por otra parte, al valorar esta variable después de concluir el período de recuperación, se encontró que solo el tallo no logró equiparar los valores de los dos tratamientos intermedios con los del control, mientras que el tratamiento con mayor contenido salino en el medio no logró equiparar sus valores con los demás. De igual forma, el tallo fue el órgano en el que las afectaciones por salinidad no fueron reversibles.
Efectos adversos del estrés salino en el crecimiento de las plantas han sido señalados en Solanum lycopersicum (5), Pennisetum clandestinum Hochst (6), Azolla pinnata–Anabaena azollae (7), al informar acerca del comportamiento de diferentes tipos de plantas en condiciones salinas (8), los que además señalaron que la salinidad ejerce su efecto perjudicial principalmente mediante desajustes del equilibrio iónico y osmótico de la célula, siendo bien conocido que la señal del estrés es primeramente recibida por los receptores a nivel de membrana y después es traducida en la célula. También se encontraron efectos depresivos en la formación de biomasa en diferentes genotipos de Vigna unguiculata sometidos a distintos tratamientos con NaCl (9). Puede señalarse el hecho de que estos resultados concuerdan con la presencia de cierta estabilidad en la tasa relativa de crecimiento a niveles de NaCl que no sobrepasaron los 100mM (10).
Como se aprecia en la Figura 2, el potencial hídrico foliar y el osmótico se diferenciaron entre los distintos tratamientos empleados desde las 24 h de aplicados y, aunque al concluir el período de recuperación mostraron tendencia a incrementarse, no lograron recuperarse.
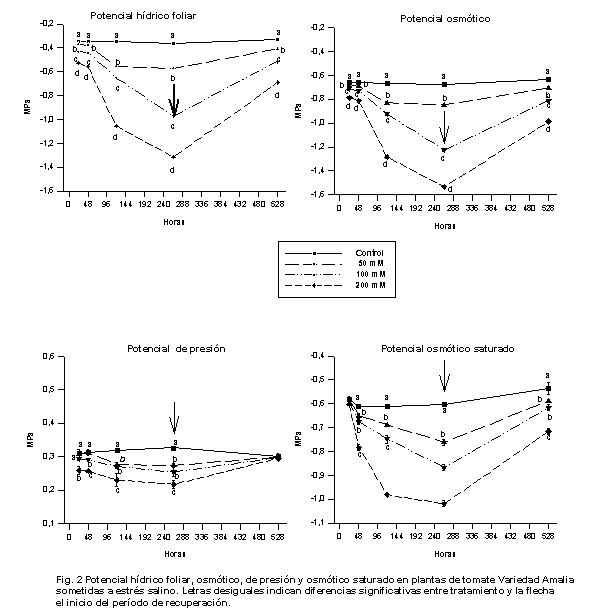
En el caso del potencial de presión, se observó que la turgencia a las 24 h de aplicados los tratamientos, solo descendió significativamente en la variante con mayor contenido de NaCl y estos mismos tratamientos presentaron valores semejantes al concluir la fase de recuperación. Se destaca, así mismo, el hecho de que esta variable haya presentado un comportamiento similar en los niveles de 50 y 100 mM de NaCl en la solución durante casi todo el período experimental.
Es interesante señalar cómo el potencial osmótico saturado, al cabo de las 24 h de aplicado el estrés, no mostró diferencias entre los tratamientos, corroborando de esta forma los resultados anteriores, en los que se encontró que al cabo de este tiempo, aún las plantas no han realizado ajuste osmótico, mecanismo que sí se pudo apreciar desde las 48 h y que se mantuvo durante el período en que las plantas estuvieron sometidas al estrés. Una vez que se suspendieron los tratamientos salinos, la turgencia se incrementó hasta alcanzar niveles muy semejantes en todas las variantes experimentales, como resultado de los aumentos de los potenciales hídrico y osmótico, al disminuir el gradiente de potencial entre el medio y la planta. Pudo apreciarse que el ajuste osmótico fue más notorio cuando los contenidos de NaCl en el medio se incrementaron, respuesta que confirma los resultados informados en plantas de Salicornia europea y Suaeda marítima (11), y Citrus limón (12).
Estudios realizados en el cultivo del tomate indicaron que la salinidad redujo no solo la acumulación de materia seca (13), sino también el potencial hídrico foliar y el osmótico, no resultando así para el de presión, que se incrementó en las condiciones de salinidad utilizadas en su trabajo, que fueron muy inferiores a las empleadas en este estudio, lo que pudiera indicar que niveles relativamente bajos de salinidad en el medio estimulan la absorción de iones y agua por las plantas. El mantenimiento del turgor en plantas de Phaseolus aureus en condiciones de salinidad se atribuyó a la acumulación de prolina libre que se produjo en tales condiciones (14).

Al analizar el comportamiento de la conductancia estomática, transpiración y conductividad hidráulica de las raíces (Figura 3), se encontró que estas variables se vieron afectadas negativamente por la salinidad en el medio desde las 24 h de aplicados los tratamientos y mantuvieron ese comportamiento durante todo el período de estrés. Sin embargo, mostraron recuperación en las dos primeras variables a los niveles de 50 y 100 mM de NaCl, mientras que, en el caso de la conductividad hidráulica, solo se recuperó el tratamiento menos salino, siendo en este caso la respuesta de las restantes variantes poco tendentes a la recuperación, lo que pudiera indicar que ya a 100 mM se producen cambios estructurales en los tejidos conductores que llegan a hacerse irreversibles.
Estudios realizados en Olea europaea L. (15) demostraron que un incremento en el contenido de NaCl en el medio en que se desarrollan las raíces no solo provocaron efectos depresivos en el crecimiento de las plantas, sino también en la fotosíntesis y transpiración, lo que está relacionado con el incremento de la resistencia al flujo de agua hacia el interior de la planta acompañada de la regulación estomática.
Los resultados que concuerdan con los informados en relación con la conductividad hidráulica de las raíces han sido señalados en plantas de otra variedad de Solanum lycopersicum (16), en estudios realizados con dos especies de tomate (17) y también han sido relacionados en Arbutus unedo (18).
Los resultados encontrados en este trabajo evidencian que la variedad cubana de tomate Amalia realiza determinados mecanismos fisiológicos, que le permiten comportarse moderadamente tolerante a los elevados niveles de NaCl durante el período vegetativo; así mismo, estos demuestran su capacidad de recuperación, una vez superadas las condiciones estresantes cuando han estado expuestas a niveles de salinidad de hasta 100 mM.
REFERENCIAS
1. Sam, O.; Ramirez, C.; Coronado, M. J.; Testillano, P. S. y Risueño, M. del C. Changes in tomato leaves induced by NaCl stress: leaf organization and cell ultrastructure. Biologia Plantarum, 2003/4, vol. 47, no. 3, p. 361-366.
2. Yeo, A. Molecular biology of salt tolerance in the context of whole plant physiology. J. Exp. Bot., 1998, vol. 49, p. 915-929.
3. Zhu, J. K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol., 2002, vol. 53, p. 247-73.
4. Parra, M. Respuestas adaptativas inducidas por halo-acondicionamiento en especies hortícolas cultivadas en medio salino. [Tesis de grado]; Universidad de Murcia, 2002, 236 p.
5. Goykovic-Cortés, V. y Saavedra del Real, G. Algunos efectos de la salinidad en el cultivo del tomate y prácticas agronómicas de su manejo. IDESIA, 2007, vol. 25, no. 3, p. 47-58.
6. Muscolo, A., Panuccio, M. R. y Sidari, M. Effects of salinity on growth, carbohydrate metabolism and nutritive properties of kikuyu grass (Pennisetum clandestinum Hochst). Plant Science, 2003, vol. 164, no. 6, p. 1103-1110.
7. Ashwani, K. R. y Vandna, R. Effect of NaCl on growth, nitrate uptake and reduction and nitrogenase activity of Azolla pinnata–Anabaena azollae. Plant Science, 2003, vol. 164, no. 1, p. 61-69.
8. Mahajan, S. y Tuteja, N. Cold, salinity and drought stresses: An overview. Archives of Biochemistry and Biophysics, 2005, vol. 444, no. 2, p. 139-158.
9. Murillo, B., Troyo, E.; García, J. L., López, R., Ysac Ávila, N., Zamora, S., Rueda, E. O. y Kaya, C. Effect of NaCl salinity in the genotypic variation of cowpea (Vigna unguiculata) during early vegetative growth. Scientia Horticulturae, 2006, vol. 108, no. 4, p. 423-431.
10. Dell’Amico, J. M. y Parra, M. Efecto del estrés por NaCl en el contenido de cloruros, el potencial osmótico real y el crecimiento de dos cultivares de tomates cubanos. Cultivos Tropicales, 2005, vol. 26, no. 2, p. 39-44.
11. Reda E., Moghaieb, A., Saneoka, H. y Fujita, K. Effect of salinity on osmotic adjustment, glycinebetaine accumulation and the beraine aldehyde dehydrogenase gene expression in two halophytic plants, Salicornia europaea and Suaeda maritime. Plant Science, 2004, vol. 166, no. 5, p. 1345-1349.
12. Pérez-Pérez, J. G., Robles, J. M., Tovar, J. C. y Botía, P. Response to drought and salt stress of lemon ‘Fino 49’ under field conditions: Water relations, osmotic adjustment and gas exchange. Scientia Horticulturae, 2009, vol. 122, no. 1, p. 83-90.
13. Romero-Aranda, R.; Soria, T. y Cuartero, J. Tomato plant-water uptake and plant-water relationships under saline growth conditions. Plant Science, 2001, vol. 160, no. 2, p. 265-272.
14. Neelam, M. y Ajay, K. G. Effect of salt stress on proline metabolism in two high yielding genotypes of green gram. Plant Science, 2005, vol. 169, no. 2, p. 331-339.
15. Tabatabaei, S. J. Effects of salinity and N on the growth, photosynthesis and N status of olive (Olea europaea L.) trees. Scientia Horticulturae, 2006, vol. 108, no. 4, p. 432-438.
16. Rodríguez, P., Dell'Amico, J., Morales, D., Sánchez-Blanco, M. de J. y Alarcón, J. J. Effects of salinity on growth, shoot water relations and root hydraulic conductivity in tomato plants. Journal of Agricultural Sciences, 1997, vol. 128, p. 439-444.
17. Morales, D., Rodríguez, P., Dell’Amico, J. M., Sánchez-Blanco, M. de J. y Torrecillas, A. Efecto de la salinidad en la conductividad hidráulica de las raíces y las relaciones hídricas en hojas de dos especies de tomate (L. esculentum y L. chessmanii). Cultivos Tropicales, 2003, vol. 24, no. 1, p. 41-45.
18. Navarro, A; Bañon, S., Olmos, E. y Sánchez-Blanco, M. de J. Effects of sodium chloride on water potential components, hydraulic conductivity, gas exchange and leaf ultrastructure of Arbutus unedo plants. Plant Science, 2007, vol. 172, no. 3, p. 473-480.
Recibido: 19 de octubre de 2009
Aceptado: 28 de Julio de 2010
