










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCultivos Tropicales
versión impresa ISSN 0258-5936
cultrop vol.31 no.4 La Habana oct.-dic. 2010
RELACIONES HÍDRICAS Y CONTENIDO IÓNICO EN PLANTAS DE MANDARINA CV FORTUNE DURANTE UN CICLO DE SEQUÍA Y RECUPERACIÓN EN LA ETAPA DE RÁPIDO CRECIMIENTO DEL FRUTO
Water relations and ionic content MANDARIN IN PLANTS FOR A FORTUNE CV cycle of drought and recovery in the stage of rapid fruit growth
Dr.C. J. M. Dell’AmicoI. E-mail: amico@inca.edu.cu, Dr.C. R. DomingoII, Dr.C. A. PérezIII, Director, M. GarcíaIII, María PeñalverIII, F. VillanuevaIII y P. PuertoIII
I Investigador Titular del departamento de Fisiología y Bioquímica Vegetal, Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, La Habana, Cuba, CP 32700
II Catedrático,
III Ingenieros Agrónomos de la Escuela Técnica Superior de Ingenieros Agrónomos de la Universidad Politécnica de Cartagena (UPCT), Murcia, España.
RESUMEN
Plantas de mandarina cv Fortune de 18 años fueron sometidas a un ciclo de sequía durante la fase de rápido crecimiento del fruto hasta alcanzar un potencial hídrico de tallo (?tallo) al mediodía de -2,5 MPa. El agua de riego se caracterizó por una CE de 25ºC y un contenido de Cl- que oscilaron entre 3,97-4,39 dS.m-1 y 14-25 meq.L-1, respectivamente, y un pH en torno a 8,4. La sensibilidad a la pérdida o ganancia de agua en hojas, frutos y tallos se manifestó a través de tasas específicas diferentes y el ?tallo fue el indicador donde primero se detectaron diferencias a favor de las plantas bien regadas. Se encontró una buena relación entre el potencial hídrico del fruto (?fruto), el potencial osmótico real de la corteza (?ORcorteza) y el potencial de presión de la corteza (?Pcorteza). Condiciones favorables de abastecimiento hídrico y transpiración propiciaron mayores contenidos de Cl- y Na+ en hojas, corteza y jugo, por considerarse las vesículas un sumidero pobre de solutos inorgánicos y escaso flujo de masa. Los mayores contenidos de Cl- en las hojas, corteza y jugo con respecto al Na+ se deben a la retención de este último en la parte maderable de raíces y tallo. El rendimiento y la calidad de la cosecha no se afectaron negativamente por la supresión del riego en esta etapa.
Palabras clave: cítricos, mandarina, estrés hídrico, relaciones hídricas, contenido iónico.
ABSTRACT
18-year-old mandarin plants cv Fortune were subjected to a drought cycle during the fast fruit growth phase until reaching a stem water potential (?stem) of -2,5 MPa at noon. Irrigation water was characterized by a CE of 25ºC and Cl- content ranging between 3,97-4,39 dS.m-1 and 14-25 meq.L-1, respectively, and a pH of about 8,4. The sensibility to water loss or gain in leaves, fruits and stems was showed through different specific rates and ?stem was the indicator where differences were first detected in favour of well-watered plants. A good correlation was found between fruit water potential (?fruit), bark actual osmotic potential (?ORbark) and bark pressure potential (?Pbark). Favourable conditions of water supply and transpiration propitiated greater Cl– and Na+ contents in leaves, bark and juice, since vesicles were considered a poor drain of inorganic solutes and scarce mass flow. The major Cl- contained in leaves, bark and juice with regard to Na+ are due to the retention of the latter one in the woody part of roots and stem. Yield and crop quality were not negatively affected by irrigation suppression in this stage.
Key words: citrus, mandarin, water stress, water relations, ion content.
INTRODUCCIÓN
La escasez de agua es una de las principales causas de la disminución del rendimiento de los cultivos, en la mayoría de las regiones agrícolas del mundo (1). Los cítricos son árboles perennes con un largo período productivo y están expuestos a frecuentes limitaciones hídricas tanto del suelo como atmosféricas (2).
Los requerimientos de riego de los cítricos son normalmente determinados en base a la evapotranspiración de referencia (ETo) y un coeficiente de cultivo estacional (Kc). Sin embargo, desafortunadamente debido a la intercepción de la luz (3) o la carga de frutos (4), estos valores pueden cambiar la estimación del uso del agua. Por ello, es necesario desarrollar métodos de programación del riego novedosos y más precisos, que permitan determinar con exactitud los requerimientos hídricos de los cítricos en cualquier momento.
El empleo de indicadores de las plantas puede ser un método ideal para la programación del riego, debido a la naturaleza dinámica del estado hídrico de las plantas, lo cual está directamente relacionado con las condiciones climáticas y del suelo (5, 6, 7).
Por otra parte, los cítricos comúnmente se cultivan en zonas donde el contenido de sales del agua de riego es bajo, por ejemplo en Australia, normalmente la conductividad eléctrica (CE) del agua es <0,5 dS.m-1, pero estos también se cultivan en regiones donde la salinidad del agua alcanza valores de CE superiores a 1,4 dS.m-1.
La capacidad de los cítricos de poder ser cultivados en suelos salinos está asociada generalmente al mecanismo de ‘exclusión’ más que a la ‘tolerancia’ de iones en las hojas (8).
Distintos ensayos de riego deficitario controlado (RDC), realizados en cítricos, demuestran que dotaciones hídricas inferiores a las necesidades totales no implican necesariamente pérdidas de producción ni de calidad del fruto y que pueden contribuir al control del crecimiento vegetativo. Sin embargo, existen pocos trabajos que estudien los efectos combinados de la reducción del riego y el empleo de aguas salinas, lo que resulta de gran interés para especies sensibles a las sales y cultivadas en climas semiáridos.
Por ello, el objetivo principal de este trabajo fue evaluar los efectos de la aplicación de un ciclo de sequía y recuperación en etapas tempranas del crecimiento del fruto, sobre las relaciones hídricas de mandarinos cv Fortune regados por goteo con agua salina.
MATERIALES Y MÉTODOS
El trabajo se realizó durante la campaña 2008-2009 en una finca comercial de mandarina cv Fortune, injertada sobre mandarino Cleopatra (Citrus reshni Hort. ex. Tanaka), situada en Miranda, Cartagena, Murcia. Los árboles de 18 años de edad, dispuestos a un marco de plantación de 5x3 m, fueron regados por goteo con dos laterales de riego por hilera de árboles y seis emisores por árbol de 4 L.h-1 cada uno.
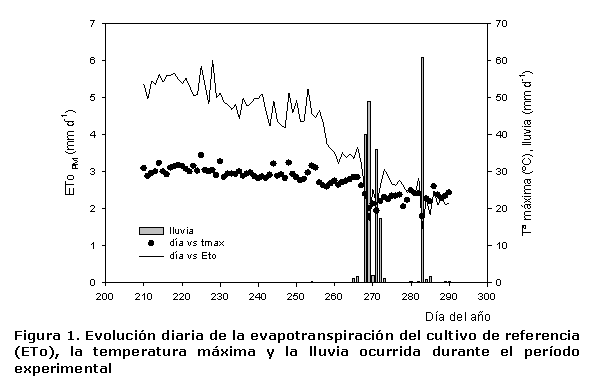
La climatología fue típicamente mediterránea (Figura 1). El suelo muy pedregoso, de textura franco-arcillosa y nivel medio de materia orgánica; está clasificado como Typic Petrocalcid (9). El agua de riego es una mezcla de aguas procedentes del trasvase Tajo-Segura y de pozo, con una CE de 25º C que osciló, durante el experimento, entre 3,97 y 4,39 dS.m-1, el pH de 7,7-9,1 y el contenido de cloruros de 500-900 mg.L-1 en función de la disponibilidad de agua del trasvase. Todos los árboles recibieron idénticos cuidados agrícolas.
Se ensayaron dos tratamientos de riego, distribuidos según un diseño experimental de bloques al azar con tres repeticiones de 15 árboles cada una. Cada réplica consistió en tres filas de cinco árboles y se consideraron los cinco de las filas centrales como área de cálculo. Los tratamientos de riego ensayados fueron:
CTL “Control” regado al 125 % de la ETc (Evapotranspiración estándar del cultivo).
EST “Estrés” supresión del riego desde el 12 de agosto hasta alcanzar el valor umbral de -2,5 MPa de potencial hídrico del tallo (?t) al mediodía (estrés severo), posteriormente con riego idéntico al CTL hasta la cosecha.
La evapotranspiración del cultivo de referencia (ETo) se calculó utilizando datos meteorológicos de una estación meteorológica cercana (aproximadamente a 2 km de la plantación) y se empleó el método de FAO Penman-Monteith. La evapotranspiración del cultivo bajo condiciones estándar (ETc) se calculó mediante la siguiente ecuación:
ETc= ETo*Kc
Donde:
ETc. Evapotranspiración del cultivo [mm.d-1]
Kc. Coeficiente del cultivo [adimensional]
ETo. Evapotranspiración del cultivo de referencia [mm.d-1]
El coeficiente de cultivo Kc empleado fue el informado por Castel para cítricos.
En el ensayo se consideró aplicar el 125 % de la ETc como tratamiento control (CTL), con el objetivo de garantizar que los árboles en este tratamiento estuviesen bien regados y sin ninguna limitación hídrica.
Durante el período comprendido entre el 14 de agosto y 3 de octubre, se realizaron 12 evaluaciones: nueve correspondieron al período de estrés y tres al de recuperación. En estas evaluaciones se midió el potencial hídrico y sus componentes en hojas (?foliar) y frutos (?fruto), el potencial osmótico a máxima saturación (?OSat) en hojas y potencial hídrico de tallo (?tallo). Todas estas evaluaciones se realizaron en tres plantas por réplica (18 medidas).
Se midieron los ?foliar y ?fruto con cámara de presión entre las 10: 00 y las 11:00 a.m., mientras que el ?tallo se midió a mediodía solar (? 13 h). Para la medida de ?tallo se cubrieron las hojas con una lámina de polietileno negro y posteriormente con papel aluminio, con una antelación mínima de dos horas. Estas evaluaciones se realizaron atendiendo a la metodología descrita (10) para evitar pérdidas de agua durante las medidas. Las hojas seleccionadas al azar se tomaron del tercio superior de las plantas, bien expuestas al sol y completamente desarrolladas. Inmediatamente después de evaluar ?foliar y ?fruto, ambas muestras se cubrieron con papel de aluminio y se congelaron en nitrógeno líquido. Para la determinación de ?OSat, se tomaron hojas aledañas a las seleccionadas para la medida de ?foliar y se colocaron en cámaras de hidratación con agua destilada a la oscuridad y entre 6 y 8ºC durante 24 h. Inmediatamente después, se envolvieron con papel de aluminio para su congelación en nitrógeno líquido y se almacenaron en congelador a –80ºC.
Posteriormente, se descongelaron a temperatura ambiente y por centrifugación se obtuvo el jugo celular de hojas y corteza. A partir de alícuotas de 100 ?L, se determinó el potencial osmótico real (?OR) y ?Osat de hojas, el ?ORCorteza y ?ORJugo de los frutos con un osmómetro automático tipo 13/13DR, calibrado con una solución estándar de 300 mOsm de NaCl. El potencial de presión (?P) de las hojas, corteza y jugo del fruto se calculó a partir de la diferencia entre el potencial hídrico y osmótico de las diferentes partes. A partir de los extractos obtenidos anteriormente, se valoraron los contenidos de Cl-, Na+ y K+ por cromatografía líquida de intercambio iónico. Para la determinación de Cl- se utilizó una columna Metrosep A Supp 5-250 con las siguientes características:
- material de relleno: resina de alcohol polivinílico con grupos de aminocuaternario
- dimensiones: 250 x 4 mm
- cuerpo de la columna: PEEK (polieteretercetona)
- flujo de trabajo: 0.7 mL.min-1
- presión de trabajo: 10-12 MPa
- tamaño de la partícula de relleno: 5 micras
- rango de pH: 3-12
- Eluyente utilizado: 1 mM en hidrogenocarbonato sódico y 3.2 mM en carbonato sódico.
El sistema cromatográfico para esta determinación consta de los siguientes dispositivos:
1. detector de conductividad
2. supresor de CO2
3. supresor iónico.
Para la determinación de Na+ y K+ se utilizó una columna Metrosep C 2-250 con las siguientes características:
- material de relleno: gel de sílice con grupos carboxilo
- dimensiones: 250 x 4 mm
- cuerpo de la columna: PEEK
- flujo de trabajo: 1mL.min-1
- presión de trabajo: 10-12 MPa
- tamaño de partícula: 7 micras
- rango de pH: 2-7
- Eluyente utilizado: 4mM en ácido tartárico y 0.75mM en ácido dipicolínico.
El sistema cromatográfico para la determinación de cationes también tiene un detector de conductividad. Los módulos utilizados para la determinación de Cl-, Na+ y K+ fueron: 838 Advanced Sample Processor y 861 Advanced Compact IC, Metrohm, Suiza.
A los 40 días de iniciado el período de recuperación, se realizó un muestreo foliar en ambos tratamientos, consistente en la recogida de 50 hojas por repetición (tres muestras por tratamiento). Las hojas de cada muestra se lavaron primero en agua corriente, después en una solución de detergente comercial al 0,1% v/v pasando los dedos índice y pulgar por toda la superficie de las hojas; posteriormente se enjuagaron en agua corriente y, por último, se sumergieron en agua desionizada durante tres minutos. Su secado en estufa de tiro forzado se realizó a 65-70ºC durante 72 h y las hojas secas se molinaron hasta obtener un polvo muy fino (11). Las determinaciones de los contenidos foliares de Na+, Cl- y K+ se realizaron por la técnica de espectrometría de fluorescencia de rayos X por dispersión de longitudes de onda, el modelo del equipo empleado fue S4 Pioneer, Broker AXS GMBH, Karlsruhe, Alemania. Se tomaron 5 g del material molinado por muestra para la elaboración de las pastillas mediante su prensado.
El rendimiento y sus componentes se evaluaron en cinco árboles por cada repetición (tres repeticiones por tratamiento). La determinación de diferentes indicadores físico-químicos de la calidad de los frutos se realizó en tres momentos de la etapa de maduración: inicio (17-11-2008), mediados (04-02-2009) y final (06–03-2009) y para ello se tomaron 10 frutos por repetición (30 frutos/tratamiento). Los indicadores de calidad evaluados fueron: masa de fruto (g), diámetro ecuatorial (cm), masas fresca y seca de corteza, masa locular y porcentaje de jugo (g), pH (pHmetro), ºBrix (refractometría), CE 25ºC (conductímetro), acidez (por valoración con NaOH 0,1 N y como indicador fenolftaleína), contenido de Cl- (autoanalizador de cloruros) y potencial osmótico del jugo (osmómetro).
Todos los indicadores antes mencionados se realizaron por técnicas convencionales de laboratorio. Para el procesamiento de los datos y la comparación de medias, se utilizó el programa estadístico SPSS 10.0 para Windows. La graficación de los resultados se realizó mediante el programa SIGMA PLOT 10.0
RESULTADOS Y DISCUSIÓN
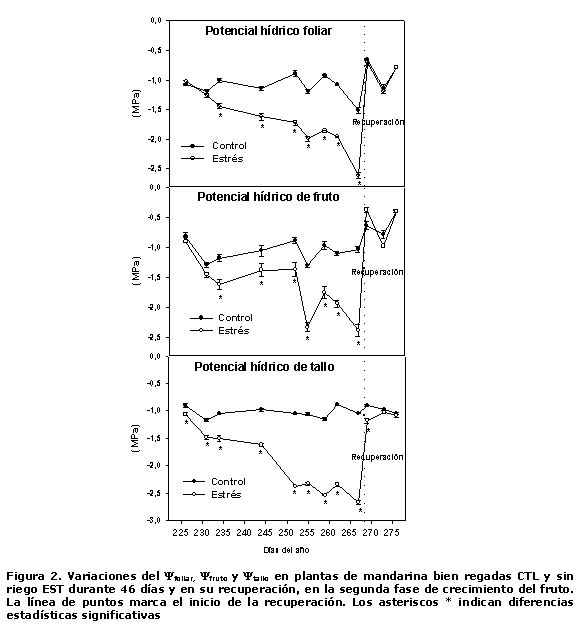
En relación con el estado hídrico de las plantas (Figura 2), se encontró que el ?foliar de las plantas del CTL se mantuvo alrededor de -1.0 MPa entre los días 225 y 262 (primeros 37 días de ensayo) y el valor más bajo -1.5 MPa se alcanzó a los 267 días. Mientras que los valores de este indicador en las plantas del tratamiento EST comienzan a diferenciarse de las del CTL a partir de los 234 días (nueve días de iniciado el ciclo de sequía). Hasta el día 244 (primeros 19 días de estrés), las diferencias entre tratamientos se mantienen en el orden de -0.5 MPa a favor de las plantas control y a partir de este momento se incrementan paulatinamente hasta alcanzar valores de -1.1 MPa al final del ciclo de sequía.
Al analizar las variaciones en el ?fruto, se encontró que las diferencias entre los frutos de las plantas de ambos tratamientos se presentaron el día 234 (siete días de estrés) a favor de las plantas bien regadas. Sin embargo, estas diferencias hasta el día 255 no sobrepasaron los -0.5 MPa. Posteriormente, el estrés hídrico provocó un descenso significativo en los valores de ?fruto hasta el final del ciclo.
Cabe señalar que el hecho de encontrarse pocas diferencias en el ?fruto entre los días 225 y 252 está estrechamente ligado a que los frutos cítricos son órganos compuestos por diferentes tipos de tejidos y el movimiento del agua a través de estos tiene lugar fundamentalmente por vías no vasculares y al analizar la relación estructura función de estos tejidos (12), se encontró poca especialización para el transporte; de hecho se registró una densidad de plasmodesma en estos tejidos no vasculares similar a la densidad presente en el parénquima de muchos sistemas.
Anteriormente, se señaló que no existen conexiones vasculares entre las vesículas (sacos de jugo) y el floema o el xilema del pericarpio (13), lo que permite considerar que los factores que afectan la transpiración y traslocación ejercen un efecto mínimo en las relaciones hídricas de las vesículas.
En el caso del ?tallo desde el inicio de los tratamientos, se presentaron diferencias significativas a favor de las plantas del CTL, las que se mantuvieron hasta el día 244 con valores de -0.7 MPa. Posteriormente, se produjo un descenso brusco en los valores de este indicador en las plantas estresadas, que oscilaron entre -2.3 y -2.4 MPa, que prevalecieron hasta los 42 días, momento en el que se sobrepasan los -2.5 MPa (valor prefijado como culminación del ciclo de sequía e inicio de la recuperación).
Resulta interesante destacar que a diferencia de las hojas y los frutos, en el ?tallo fue donde primero se detectaron diferencias entre las plantas de ambos tratamientos, pero fue el que más tardó en recuperarse.
Estos resultados demuestran que la sensibilidad a la pérdida o ganancia de agua en estos órganos (hojas, frutos y tallo), dadas sus propias características, se manifiesta a través de tasas específicas diferentes. Es por ello que muchos (14, 15) reconocen el ?tallo como un indicador confiable del estado hídrico de los frutales, incluso ante condiciones ligeras de estrés y ha sido frecuentemente utilizado como referencia en la programación del riego en diferentes cultivos: melocotonero (6), almendra (7), entre otros.
En la Figura 3 se presentan los resultados alcanzados en ?OR y ?Osat de hojas, donde se pudo apreciar que en el caso del ?OR, los valores entre los días 225 y 255 (primeros 30 días de iniciado el tratamiento EST), tanto en las plantas estresadas como en las del tratamiento CTL, se mantuvieron entre -2.0 y -2.5 Mpa con algunas diferencias significativas.
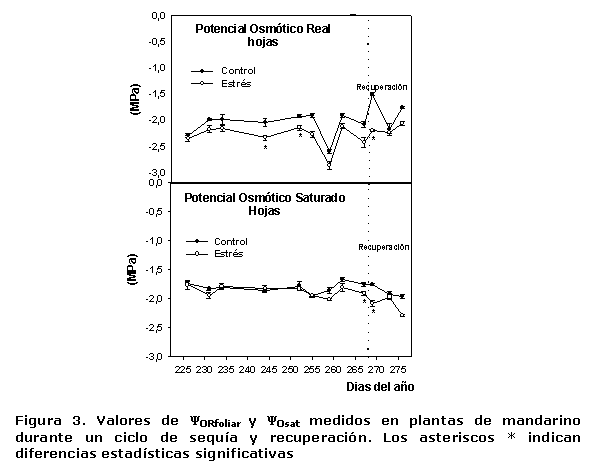
Entre los días 255 y 260 se produjo un descenso brusco del ?ORfoliar en las plantas de ambos tratamientos y, posteriormente, los valores vuelven a ubicarse en el entorno de -2.0 y -2.5 Mpa. Solamente a las 24 h de iniciarse la recuperación, se observaron diferencias estadísticas significativas. Los valores de ?Osat fueron muy similares en las plantas de los tratamientos en estudio, hasta los 267 días de iniciado el estrés, los que oscilaron entre -1.5 y -2.0 Mpa. Al final del periodo de estrés, entre los 267 y 270 días del ciclo de sequía, puede decirse que existió una tendencia al ajuste osmótico, al detectarse diferencias estadísticamente significativas en los valores de esta variable entre las plantas de los dos tratamientos. Sin embargo, fue precisamente a los 268 días que se alcanzó el valor más bajo de ?P en las hojas (Figura 5), lo que indica que la disminución de los valores de ?Osat en estos días no se debió a la acumulación de sustancias osmóticamente activas. Es de señalar que la no ocurrencia del proceso de ajuste osmótico en estas evaluaciones, se deba a fundamentalmente a la aclimatación de los árboles que durante 18 años se han cultivado en esas condiciones.
Las variaciones en los ?ORcorteza y ?ORjugo se presentan en la Figura 4, donde se puede apreciar que en el caso de ?ORcorteza prácticamente en todas las evaluaciones se encontraron diferencias significativas y los valores mayores siempre se presentaron en las plantas de TCL y se mantuvieron entre -1.26 y -1.45 MPa, mientras que en EST estos oscilaron entre -1.32 y -2.17 MPa durante el ciclo de sequía. Por otra parte, el comportamiento de los valores del ?ORjugo fue acentuadamente variable en las plantas de los tratamientos estudiados y, en general, alcanzaron magnitudes mucho más negativas a los ?ORcorteza (valores entre -2.0 y -4.0 MPa).
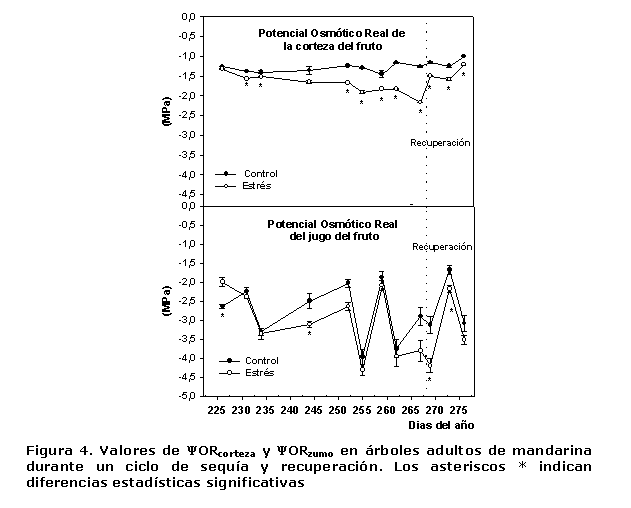
Atendiendo a la estructura y composición de los frutos cítricos, se conoce que en su interior, muchas sustancias solubles en agua se encuentran en concentraciones de medias a altas, como la sacarosa que puede ser mayor de 250 mM (16) en la etapa de rápido crecimiento, lo que tiene un efecto importante en el potencial osmótico, fundamentalmente del jugo. Como la concentración de estas sustancias osmóticamente activas varía entre los distintos tejidos del fruto en desarrollo, justifica las variaciones en sus diferentes partes y el establecimiento de gradientes de potencial osmótico, que favorece la traslocación de la sacarosa hacia el interior de las vesículas de jugo (17).
El ?P calculado por la diferencia entre el potencial hídrico y el osmótico, en el caso particular del fruto, se asumió el mismo potencial hídrico del fruto para el cálculo del potencial de turgencia de la corteza y el jugo.
En la Figura 5 se presentan los resultados de esta variable, donde se encontró que tanto en las hojas como en la corteza el estrés hídrico provocó que se alcanzaran valores negativos de turgencia, lo que fue mucho más notorio en la corteza del fruto. Mientras que en el ?P del jugo, aun cuando los valores en las plantas de ambos tratamientos presentaron una gran variabilidad durante el periodo de estrés, en ningún momento se alcanzaron valores negativos.
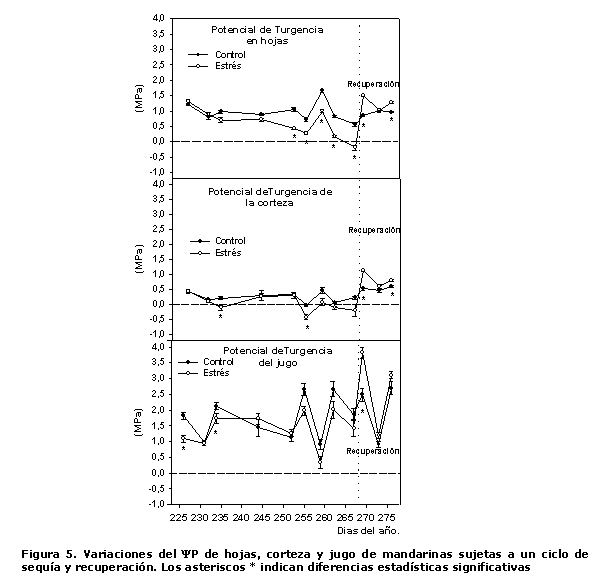
Este comportamiento está relacionado con el hecho de que tanto el tejido foliar como el de la corteza del fruto están vinculados al torrente transpiratorio y ejercen poca resistencia relativa a la pérdida de vapor de agua por transpiración y por el movimiento del agua desde la corteza del fruto a las hojas adyacentes (13). Mientras que en el caso de las vesículas del jugo, las características anatómicas de la cutícula que las cubre las hace completamente impermeables, por lo que el movimiento del agua y los solutos tiene lugar únicamente a través de los pedúnculos de las vesículas. Por otra parte, se ha señalado que las vesículas de jugo para su crecimiento en extensión requieren de una turgencia relativamente elevada (12), al igual que ocurre con el fruto del Litchi (18), capaz de vencer la barrera física de la corteza. Además, la turgencia celular en el interior de las vesículas es conservada por la corteza, aun cuando existan condiciones que propicien una transpiración elevada. Así, la transpiración de las hojas próximas al fruto afectará a la corteza, pero no a las vesículas de jugo.
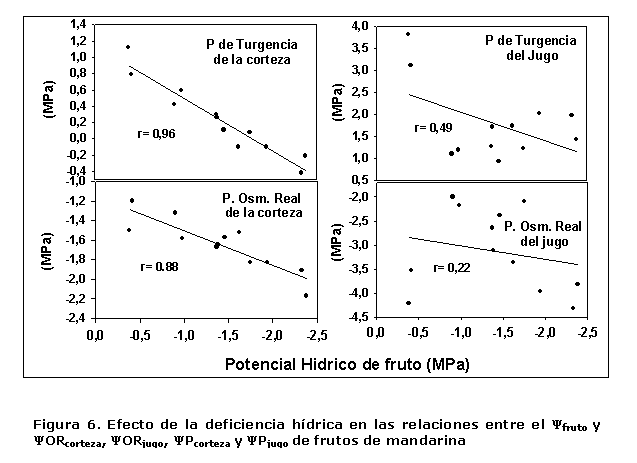
Una mayor claridad en este sentido se obtuvo al relacionar los valores del ?fruto con ?ORcorteza, ?ORjugo, ?P de la corteza y el jugo (Figura 6), donde se encontró una buena relación entre el ?fruto, ?ORcorteza y el ?P de corteza, r= 0,88 y r= 0,96, respectivamente. Mientras que en las relaciones con ?ORjugo y ?Pjugo, los valores fueron relativamente bajos.
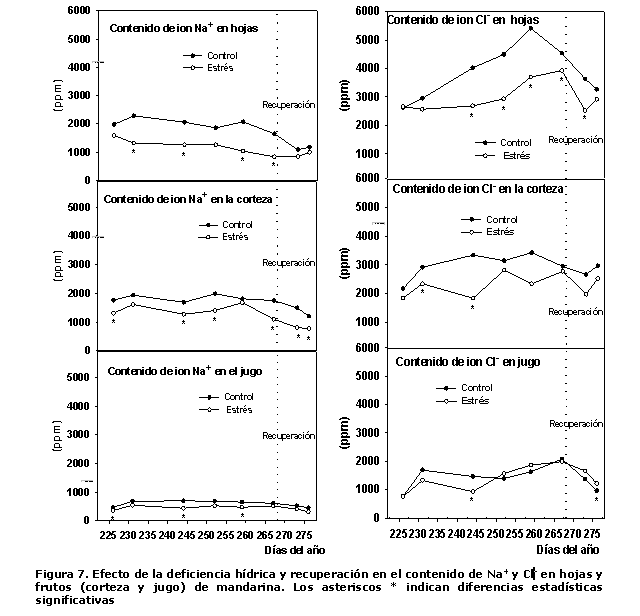
Contenido iónico. Los valores de contenido de los iones Na+ y Cl- en hojas, corteza y jugo del fruto se presentan en la Figura 7, donde se encontró que, en general, presentaron una tendencia similar, mayor concentración en hojas y menor en zumo, con valores intermedios en la corteza de los frutos.
En las plantas del tratamiento CTL, los contenidos de Na+ y Cl- en el fruto dependieron en un 67 % (r= 0.67) y 60 % (r= 0.60), respectivamente, de los contenidos en las hojas. Mientras que en las plantas sujetas a estrés hídrico fueron del -4 % (r= -0.04) y 71 % (r= 0.71), respectivamente de los contenidos en las hojas. Cabe señalar que las concentraciones de Na+ y Cl- en las hojas, la corteza y el jugo fueron significativamente mayores en las plantas del tratamiento CTL.
Por otra parte, los contenidos de Cl- en hojas y frutos fueron superiores a los de Na+ en ambos tratamientos. El hecho de encontrarse mayores contenidos de Na+ y Cl- en hojas que en el jugo de las plantas de ambos tratamientos, se debe fundamentalmente a que no existen conexiones vasculares entre las vesículas de jugo y el floema o xilema del pericarpio, lo que indica su desvinculación de la vía de transporte hídrico y transpiración dominantes en las hojas y corteza. Por otra parte, las características anatómicas de las vesículas hacen que el flujo de masa en su interior sea relativamente bajo, en relación con la corteza y las hojas, por lo que se consideran sumideros débiles Na+ y Cl- comparado con las hojas, aspecto señalado con anterioridad (8). Los mayores contenidos de Na+ y Cl- en hojas, corteza y jugo se presentaron en las plantas del TCL, lo que indica que condiciones favorables de abastecimiento hídrico y transpiración propiciaron una mayor absorción y transporte de estos iones hacia el interior de las plantas. Como ya ha sido señalado (19, 20), los procesos de absorción y acumulación de los iones Na+ y Cl- en los cítricos son dependientes de la transpiración.
En el cultivo de los cítricos la mayoría de los desórdenes fisiológicos y anatómicos provocados por la salinidad son asociados con mayor frecuencia a la mayor concentración de Cl- en sus hojas, que en muchas ocasiones supera ampliamente las de Na+ (20), por lo que la mayor o menor tolerancia a la salinidad de los cítricos está vinculada con la mayor o menor capacidad de las plantas de limitar la absorción de Cl- y su acumulación en las hojas (10, 20).
En otras investigaciones (8, 20) se obtuvieron evidencias sólidas de que en variedades de cítricos sensibles o tolerantes a la salinidad, la absorción de Cl- y agua están directamente relacionadas y que las cantidades de Cl- en las hojas están determinadas muy probablemente por las tasas de transpiración. También se señala que en condiciones de salinidad, la absorción de Cl- es fundamentalmente pasiva (11, 21, 22). Otros plantean que aunque la redistribución de Cl- en el perfil del suelo (23) probablemente esté dominada por flujos de masa y procesos de convección asociados con el movimiento del agua, en este caso un movimiento de difusión importante puede ocurrir en suelos donde el gradiente de concentración de Cl- y la humedad sean altos.
A diferencia de otras especies, algunas plantas leñosas (perennes) y particularmente en los cítricos, el Na+ es retenido en la parte maderable de las raíces y basal del tallo, y es el Cl- el que mayor acumulación tiene en la parte aérea, resultando mucho más perjudicial para las plantas (24).
En general, se puede concluir que las condiciones de abastecimiento hídrico y transpiración propiciaron mayores contenidos de Cl- y Na+ en hojas, corteza y jugo, por considerarse las vesículas un sumidero pobre de solutos inorgánicos con escaso flujo de masa. Los mayores contenidos de Cl- en las hojas, corteza y sumo respecto al Na+ se deben fundamentalmente a que los cítricos son mucho más sensibles al Cl- que al Na+, por ser este último retenido en la parte maderable de raíces y tallo. Todo lo anterior indica que las plantas del EST fueron favorecidas, en cuanto a los efectos dañinos de la salinidad se refiere, al propiciar el estrés hídrico menores contenidos foliares principalmente de Na+ y Cl-.
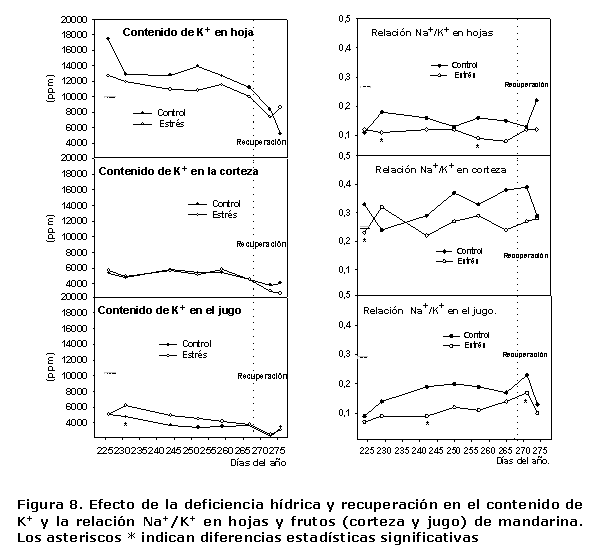
Los valores obtenidos en los contenidos de K+ y la relación Na+/K+ en hojas, corteza y jugo, se presentan en la Figura 8, donde se encontró que en general los valores de concentración de este ión en las diferentes partes analizadas fueron relativamente muy superiores a los de Na+ y Cl-. Tanto en las hojas como en la corteza, no se apreciaron diferencias estadísticamente significativas entre las plantas del TCL y el EST, lo que indica que el ciclo de sequía impuesto a las plantas no ejerció un efecto negativo acentuado, en cuanto a la absorción y concentración de este ión en hojas y corteza, aunque es de señalar que en las hojas los valores fueron ligeramente más altos en las plantas que tuvieron un mayor abastecimiento hídrico durante esa etapa. Al analizar este indicador en el jugo, aunque los contenidos fueron muy similares en las plantas de ambos tratamientos, su comportamiento fue diferente al de las hojas y los valores más altos de concentración estuvieron a favor de las plantas estresadas.
Respecto a la relación Na+/K+ (Figura 8), aunque solamente en determinadas evaluaciones se encontraron diferencias estadísticamente significativas, principalmente a favor de las plantas del TCL, este comportamiento ratifica que ellas estuvieron más afectadas negativamente por la salinidad, al presentar los valores más altos de esta relación.
Es bien conocido que la utilización de aguas con altos contenidos de sales para el riego de los cultivos, además de provocar limitaciones en la absorción hídrica y el crecimiento, también puede causar desbalances nutricionales y efectos de toxicidad por iones específicos. La presencia de altos contenidos de Na+ y Cl- puede limitar la toma de otros nutrientes, mediante interacciones competitivas o por alteración de la selectividad de las membranas. Sin embargo, en el sistema radical de los cítricos, como en muchas glicofitas, se ha comprobado que ante determinadas concentraciones externas de K+ y Na+, existe una mayor selectividad por el K+ que por el Na+, lo que justifica en este caso su mayor contenido con respecto al Na+.
En trabajos realizados en mandarina (25), se encontró que en plantas tratadas con 60 y 90 mM de NaCl hubo un incremento significativo del K+ foliar, lo que podría ser un mecanismo de regulación y mantenimiento del balance osmótico ante niveles altos de Cl- propiciados por el estrés salino.
Otros trabajos realizados en cebada (26) señalan que cálculos termodinámicos demostraron que el K+ fue transportado activamente al interior de las vacuolas de células epidérmicas y del mesófilo tanto en plantas afectadas o no por la salinidad. Analógicamente, con el mantenimiento de bajos contenidos de Na+ (24), se mantienen altos contenidos foliares de K+ y para muchas especies resulta más importante mantener bajas relaciones Na+/K+ que simplemente mantener bajos contenidos de Na+ (26).
Los resultados del análisis por espectrometría de fluorescencia de rayos X de muestras foliares a los 40 días del restablecimiento del riego a las plantas del EST, se presentan en la Tabla I, donde se encontró que en general los contenidos de Na+ fueron inferiores a los de Cl- y K+.

Solamente se apreciaron diferencias estadísticamente significativas en los contenidos de Na+ y los valores de la relación Na+/K+, en ambos casos a favor de las plantas del TCL. Sin embargo, aunque sin diferencias significativas, los contenidos más altos de Cl- y K+ se encontraron en las plantas de EST. Estos resultados indican que el restablecimiento del riego propició una mayor absorción y acumulación foliar de Cl- y K+, siendo mucho más limitadas en cuanto al Na+, comportamiento que se debe a las limitaciones que tiene el Na+ en cuanto a su transporte hacia la parte aérea de las plantas y que, además, propició valores más altos de la relación Na+/K+ en las plantas de TCL, lo que indica su mayor afectación por el estrés salino.
Los valores obtenidos en cuanto al rendimiento y sus componentes se presentan en la Tabla II, donde se encontró que el hecho de aplicar un estrés hídrico severo (hasta alcanzar -2,5 MPa de ?tallo) en la fase de rápido crecimiento del fruto, solamente provocó diferencias estadísticamente significativas en la masa promedio de los frutos a favor de las plantas del TCL.
Por otra parte, es oportuno señalar que los valores del rendimiento en gramos y número de frutos/planta fueron superiores en las plantas del tratamiento EST en un 10,7 y 21,35 %, respectivamente con respecto al TCL, sin que se apreciaran diferencias estadísticas significativas entre tratamientos.
El rendimiento constituye uno de los procesos más complejos de los cultivos y depende en gran medida de la acción de muchos factores, tanto naturales (condiciones edafoclimáticas y de cultivo) como artificiales (fertilización, riego, tratamientos químicos, etc.).
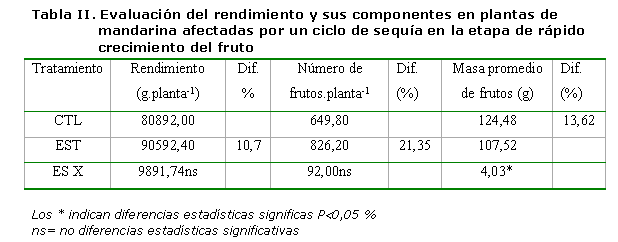
Específicamente en los cítricos, se han realizado muchos trabajos relacionados con la selección de un adecuado esquema de riego y la mejora del rendimiento (27, 28, 29, 30, 31), trabajando en mandarina con diferentes tratamientos de RDC aplicados en diferentes periodos fenológicos del crecimiento del fruto, y se encontró que la reducción del riego en un 75 % en la fase II (crecimiento rápido del fruto) no causó descensos del rendimiento ni en el número de frutos por árbol, solamente diferencias estadísticamente significativas en la masa promedio de los frutos, resultados que coinciden en gran medida con los obtenidos en este trabajo. Otros resultados similares han sido informados (32, 33) en trabajos realizados con naranja dulce cv “Lane late”.
Finalmente, se puede señalar que los cítricos, por sus bondades, dan la posibilidad a los productores, mediante un manejo adecuado del riego, de ahorrar ciertos volúmenes de agua y energía, sin perjudicar el rendimiento del cultivo, aspectos que en gran medida contribuyen a su rentabilidad, además de disminuir en cierta medida los efectos nocivos de la salinidad.
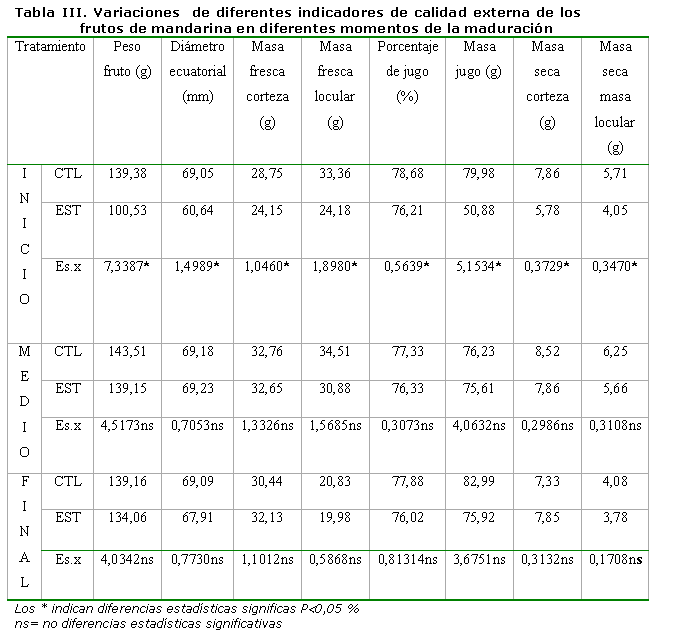
La evaluación de diferentes indicadores de calidad realizados a los frutos, tanto externos como internos, se presenta en las Tablas III y IV, donde se pudo apreciar que en los indicadores de carácter externo (Tabla III) el efecto del tratamiento de estrés hídrico se manifestó solamente al inicio de la maduración, provocando diferencias estadísticamente significativas en todos los indicadores evaluados a favor de las plantas del TCL, que desaparecieron en la medida que avanzó el proceso de maduración en los frutos.
Por otra parte, en los indicadores que aparecen en la Tabla IV, no se apreció una tendencia definida en su comportamiento, de modo que al inicio de la maduración los valores de SST y el potencial osmótico fueron estadísticamente superiores en las plantas del EST y al final de la maduración solamente se encontraron ligeras diferencias en los valores de pH, algo más pronunciadas en el contenido de Cl- a favor de las plantas del CTL, que comenzaron a detectarse desde mediados de la maduración.

Resulta de gran interés resaltar que el estrés hídrico no ejerció efecto negativo en los indicadores de calidad interna de los frutos; por el contrario, se obtuvieron frutos con menor contenido de Cl-.
Al igual que el rendimiento, la calidad de los frutos, tanto externa como interna, depende de muchos factores; en el caso específico de los frutos cítricos, se ha señalado que el manejo adecuado del riego tiene una importancia decisiva en este aspecto, atendiendo principalmente a las etapas fenológicas en que se hacen restricciones en el abastecimiento hídrico al cultivo (34).
En general, los resultados mostraron que en ausencia de estrés hídrico, existe una buena relación entre el potencial hídrico del fruto (?fruto), el potencial osmótico real de la corteza (?ORcorteza) y el potencial de presión de la corteza (?Pcorteza). La mala calidad del agua de riego propició mayores contenidos de Cl- y Na+ en hojas, corteza y jugo, cuando las condiciones de abastecimiento hídrico y transpiración fueron favorables. El estrés hídrico (-2,5 MPa de potencial hídrico de tallo) en la fase de rápido crecimiento del fruto de mandarina Fortune no ejerció un efecto negativo en el rendimiento y la calidad de la cosecha.
AGRADECIMIENTOS
El colectivo de autores agradece a la Fundación SENECA de Murcia y al Departamento de Producción Vegetal de la ETSIA de la Universidad Politécnica de Cartagena (UPCT), por el apoyo brindado para la realización de este trabajo de investigación.
REFERENCIAS
1. Bajaj, S., Jayaprakash, T., Li-Frei, L., Ho, T. H. D. y Wu, R. Transgenic approaches to increase dehydration-stress tolerance in plants, Mol. Breed., 1999, vol. 5, p. 493–503.
2. Correa, H. B., Marura, C. J., Bespalhok, J. C., Kobayashib, A. K., Pileggic, M., Pereira, R., Protasio, L. F. y Esteves, L. Osmotic adjustment in transgenic citrus rootstock Carrizo citrange (Citrus sinensis Osb. x Poncirus trifoliata L. Raf.) overproducing proline. Plant Science, 2004, vol. 167, p. 1375–1138.
3. Consoli, S., O’Connell, N., Snyder, R. Measurement of light interception by navel orange orchard canopies: case study of Lindsay, California. J. Irrig. Drain. Eng., 2006, vol. 132, p. 9–20.
4. Yonemoto, Y., Matsumoto, K., Furukawa, T., Asakawa, M., Okuda, H., Takahara, T. Effects of rootstock and crop load on sap flow rate in branches of ‘Dhirakawa Satsuma’ mandarin (Citrus unshiu Marc.). Sci. Hortic., 2004, vol. 102, p. 295–300.
5. Goldhamer, D. A., Fereres, E., Salinas, M. Can almond trees directly dictate their irrigations needs? Calif. Agric., 2003, vol. 57, p. 138–144.
6. Remorini, D., Massai, R. Comparison of water status indicators for young peach trees. Irrig. Sci., 2003, vol. 22, p. 39–46.
7. Ortuño, M. F., Alarcón, J. J., Nicolás, E. y Torrecillas, A. Sap flow and trunk diameter fluctuations of young lemon trees under water stress and rewatering. Environmental and Experimental Botany, 2005, vol. 54, p. 155–162.
8. Storey, R. y Walker, R. R. Citrus and salinity. Scientia Horticulturae, 1999, vol. 78, p. 39-81.
9. Soil Survey Staff. Claves para la taxonomía de suelos. Dpto. de Agricultura de los Estados Unidos. Décima Edición. Versión al español, 2006, 331 p.
10. Turner, N. C. Measurement of plant water status by the pressure chamber technique. Irrigation Science, 1988, vol. 9, p. 289–308.
11. García-Sánchez, F. y Syvertsen, J. P. Salinity tolerance of Cleopatra mandarin and Carrizo Citrange citrus rootstock seedlings is affected by CO2 enrichment during growth. J. Amer. Soc. Hort. Sci., 2006, vol. 131, no. 1, p. 24-31.
12. Koch, K. E. y Avigne, W. T. Postphloem. Nonvascular transfer in citrus. Kinetics, metabolism and sugar gradients. Plant Physiol., 1990, vol. 93, p. 1405-1416.
13. Kaufmann, M. R. Water potential components in growing citrus fruits. Plant Physiol., 1970, vol. 46, p. 145-149.
14. Intrigliolo, D. S. y Castel, J. R. Performance of various water stress indicators for prediction of fruit size response to deficit irrigation in plum. Agric. Water Management, 2006, vol. 83, p. 173–180.
15. Velez, J. E., Intrigliolo, D. S. y Castel, J. R. Scheduling deficit irrigation of citrus trees with maximum daily trunk shrinkage. Agricultural Water Management, 2007, vol. 90, p. 197-204.
16. Etxeberria, E., González, P. y Pozueto-Romero, J. Sucrose transport into citrus juice cells: evidence for an endocytic transport system. J. Amer. Soc. Hort. Sci., 2005, vol. 130, no. 2, p. 269-274.
17. Barry, G. H., Castle, W. S. y Davies, F. S. Rootstocks and plant water relations affect sugar accumulation of citrus fruit via osmotic adjustment. J. Amer. Soc. Hort. Sci., 2004, vol. 129, no. 6, p. 881-889.
18. Huang, H. y Qiu, Y. Growth correlations and assimilate partitioning in the arilate fruit of Litchi chinensis Sonn. Aust. J. Plant Physiol., 1987, vol. 14, p. 181-188.
19. Walker, R. R., Munns, R. y Tonnet, M. L. Xylem chloride and sodium concentrations of salt treated citrus plants, En: Beilby, M. J., Walker, N. A., Smith, J. R. (Eds.). Membrane Transport in Plants and Fungi. Sydney: University of Sydney, 1990, p. 490-494.
20. Moya, J. L., Gómez-Cadenas, A., Primo-Millo, E. y Talón, M. Chloride absorption in salt-sensitive Carrizo citrange and salt-tolerant Cleopatra mandarin citrus rootstocks is linked to water use. Journal of Experimental Botany, 2003, vol. 54, no. 383, p. 825-833.
21. Moya, J. L., Tadeo, F. R., Gómez-Cadenas, A., Primo-Millo, E. y Talón, M. Transmissible salt tolerance traits identified through reciprocal grafts
between sensitive Carrizo and tolerant Cleopatra citrus genotypes. J. Plant Physiol., 2002, vol. 159, p. 991–998.
22. Moya, J. L., Primo-Millo, E. y Talon, M. Morphological factors determining salt tolerance in citrus seedlings: the shoot to root ratio modulates passive root uptake of chloride ions and their accumulation in leaves. Plant, Cell, Environment, 1999, vol. 22, p. 1425–1433.
23. White, P. J. y Broadley, M. R. Chloride in soils and its uptake and movement within the plant: a review. Annals of Botany, 2001, vol. 88, p. 967-988.
24. Tester, M. y Davenport, R. Na+ tolerance and Na+ transport in higher plants. Annals of Botany, 2003, vol. 91, p. 503-527.
25. García-Sánchez, F. Jifon, J. L., Carvajal, Micaela y Syvertsen, J. P. Gas exchange, chlorophyll and nutrient contents in relation to Na+ and Cl- accumulation in Sunburst mandarin grafted on different rootstocks. Plant Science, 2002, vol. 162, p. 705-712.
26. Cuin, T. A., Miller, A. J., Laurie, S. A. y Leigh, R. A. Potassium activities in cell compartments of salt-grown barley leaves. Journal of Experimental Botany, 2003, vol. 54, no. 383, p. 657-661.
27. Burke, S., Mulligan, M. y Thornes, J. B. Optimal irrigation efficiency for maximum plant productivity and minimum water loss. Agric. Water Management, 1999, vol. 40, p. 377–391.
28. Barragan, J. y Wu, I. P. Optimal scheduling of a microirrigation system under deficit irrigation. J. Agric. Eng. Res., 2001, vol. 80, no. 2, p. 201–208.
29. Alva, A. K., Paramasivam, S., Graham, W. D.y Wheaton, T. A. Best nitrogen and irrigation management practices for citrus production in sandy soil. Water, Air, Soil Pollut., 2003, vol. 143, p. 139–154.
30. Morimoto, T., Ouchi, Y., Shimizu, M. y Baloch, M. S. Dynamic optimization of watering Satsuma mandarin using neural networks and genetic algorithms. Agric. Water Management, 2007, vol. 93, p. 1–10.
31. González-Altozano, P. y Castel, J. R. Riego deficitario controlado en Clementina de Nules. I. Efectos sobre la producción y la calidad de la fruta. Span. J. Agric. Res., 2003, vol. 1, no. 2, p. 81–92.
32. Hutton, R. J., Landsberg, J. J. y Sutton, B. G. Timing irrigation to suit citrus phenology: a means of reducing water use without compromising fruit yield and quality? Aust. J. Exp. Agric., 2007, vol. 47, no. 1, p. 71–80.
33. Pérez-Pérez, J. G., Romero, P., Navarro, J. M. y Botía, P. Response of sweet orange cv Lane late to deficit-irrigation strategy in two rootstocks. II: Flowering, fruit growth, yield and fruit quality. Irrig Sci., 2008, vol. 26, p. 519–529.
34. Treeby, M. T., Henriod, R. E., Bevington, K. B., Milne, D. J. y Storey, R. Irrigation management and rootstock effects on navel orange (Citrus sinensis L. Osbeck) fruit quality. Agricultural Water Management, 2007, vol. 91, p. 24–32.
Recibido: 29 de octubre de 2009
Aceptado: 28 de septiembre de 2010
