










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCultivos Tropicales
versión impresa ISSN 0258-5936
cultrop vol.32 no.3 La Habana jul.-set. 2011
Revisión Bibliográfica
LAS ACUAPORINAS, SU RELACIÓN CON LOS HONGOS MICORRÍZICOS ARBUSCULARES Y EL TRANSPORTE DE AGUA EN LAS PLANTAS.
Aquaporins, their relationship with arbuscular mycorrhizal fungi AND WATER TRANSPORT IN PLANTS.
Michel Ruiz Sánchez1, Juan Manuel Ruiz - Lozano2 , Yaumara Muñóz Hernández3
1 Investigador Agregado, Profesor Asistente, Unidad Científico Tecnológica de Base “Los Palacios” Km 1 ½ Carretera La Francia, Los Palacios, Pinar del Río, 29200. Cuba. E-mail: mich@inca.edu.cu
2 Profesora Asistente. Universidad Hermanos Saiz Montes de Oca Sede Universitaria Municipal “Los Palacios” Calle 22, entre 19 y 21. Los Palacios, Pinar del Río, 29200. Cuba.
3 DrC.Juan Manuel Ruiz Lozano. Profesor Titular del CSIC, España. Departamento de Microbiología del Suelo y Sistemas Simbióticos, Estación Experimental del Zaidín (CSIC), Profesor Albareda no. 1, E-18008 Granada, Spain.
RESUMEN
El descubrimiento de las acuaporinas en plantas ha originado un cambio muy importante en la forma de estudiar las relaciones hídricas de las plantas, por lo que en los últimos años se han realizado esfuerzos considerables para investigar la funciones y regulaciones de estas, puesto que, en la naturaleza las plantas están continuamente expuestas a condiciones de estrés que retrasan su crecimiento y desarrollo, siendo la sequía, la salinidad y las bajas temperaturas los estreses más comunes a los que deben hacer frente. El objetivo de este trabajo es compilar información actualizada, acerca del descubrimiento y tipos de acuaporinas, su funcionalidad en el transporte de agua de las plantas, su interacción con los hongos micorrízicos arbuscular, así como, algunas investigaciones realizadas en el cultivo del arroz.
Palabras claves: proteinas, hongos, micorrizas arbusculares vesiculares, relaciones planta agua, estrés de sequía, arroz.
ABSTRACT
The discovery of aquaporins in plants has caused a major change in the way of studying the water relations of plants, so that in recent years considerable efforts have been made to investigate the functions and regulations of these, since in nature the plants are continuously exposed to stress conditions that retard its growth and development, being drought, salinity and low temperatures common stresses to which they face. The aim of this work is to compile updated information about the discovery and types of aquaporins, their function in water transport of plants, their interaction with arbuscular mycorrhizal fungi, and some research in rice cultivation.
key words: proteins, fungi, vesicular arbuscular mycorrhizae, plant water relations, drought stress, rice.
INTRODUCIÓN
El agua es la molécula más abundante en la superficie terrestre, su disponibilidad es el factor que limita la presencia (cantidad y variedad) de seres vivos, desde la escala global de los ecosistemas terrestres, hasta la local (productividad de un suelo) (1). Las plantas están constantemente expuestas a limitaciones ambientales de orígenes bióticos y abióticos. Estreses abióticos como la sequía, la salinidad y temperaturas extremas son los factores ambientales más comunes experimentados por las plantas terrestres (2), este conjunto de estreses comparte un componente osmótico común, ya que todos ellos originan una deshidratación de los tejidos en las plantas (3). De hecho, el déficit hídrico originado por estreses de tipo osmótico afecta al desarrollo y crecimiento normal de la planta y tiene un efecto negativo importante sobre la supervivencia y productividad de las plantas. En la actualidad, se estima que alrededor de un 28 por ciento de los suelos terrestres están demasiado secos para soportar un desarrollo y productividad adecuada de las plantas (3). Además de existir un consenso amplio en cuanto a los efectos del cambio climático que conduce a estreses bióticos y abióticos extremos, los cuales van a continuar y se incrementarán en los próximos años, dificultando el crecimiento y desarrollo de las plantas en muchas regiones de la tierra, en especial en las actuales zonas semi-áridas (4).
La disminución del potencial hídrico en los suelos afectados por limitación hídrica obliga a las plantas a hacer frente al problema de captar cantidades suficientes de agua a través de sus raíces (5), un proceso en el que participan las acuaporinas (6).
El descubrimiento de las acuaporinas en las plantas ha provocado un cambio significativo en la comprensión de las relaciones hídricas de las plantas. En los últimos años, mucho esfuerzo se ha concentrado en investigar la función y regulación de las acuaporinas. Los niveles altos de expresión de acuaporinas se muestran en los tejidos con flujos altos de agua a través de membrana, por ejemplo, en las regiones de rápido crecimiento, en los brotes y en las hojas, también en las raíces donde se produce la absorción de agua (7). Las acuaporinas parecen tener una función especialmente importante en el control del transporte transcelular de agua en los tejidos vegetales (8). La expresión de tantas isoformas de acuaporinas en plantas induce a preguntarse si su capacidad de conducir agua entre membranas es realmente su función esencial (9, 10) y la relevancia fisiológica del transporte a través de acuaporinas de solutos pequeños, gases y, probablemente iones, comienza a ser un aspecto de máximo interés, en especial en el caso de las plantas micorrizadas, donde hay un intercambio de nutrientes y de agua a través de membranas entre el hongo y la planta (11). Partiendo de que las acuaporinas fueron descubiertas recientemente y se han iniciado investigaciones en las plantas, el objetivo de esta revisión bibliográfica es resumir información actualizada sobre los tipos de acuaporinas, funcionalidad en el transporte de agua, su interacción con los hongos micorrízicos arbuscular, así como, algunas investigaciones realizadas en el cultivo del arroz.
LAS ACUAPORINAS (AQPs).
Hasta hace algunos años se pensaba que el paso del agua a través de las membranas era por simple difusión, cruzando la bicapa lipídica, sin embargo, existían evidencias que sugerían que la entrada y salida de agua de algunas células debería de estar mediado por un sistema que selectivamente dejaban pasar el agua a través de las membranas (12). El agua es el componente principal de células y tejidos de todos los organismos, y la manera de cómo se regula la entrada y salida del agua en las células debe de ser uno de los aspectos más importante en el estudio de la fisiología de los organismos en general (13).
El descubrimiento de las acuaporinas surgió de una observación accidental que permitió el conocimiento de proteínas que actúan regulando la circulación del agua (14). La proteína fue aislada y se determinó su secuencia de aminoácidos. En el año 1992 Peter Agre logró identificar esta proteína tan buscada, a la que bautizó con el nombre de acuaporinas (AQPs). Dicho investigador demostró su hipótesis al inyectar material genético extraído de glóbulos rojos humanos en el ovocito de un anfibio, célula que normalmente no expresa canales y observó un aumento espectacular de la permeabilidad al agua. Cuando los ovocitos se colocaron en una solución acuosa, las células que tenían acuaporinas pudieron absorber agua por ósmosis y se hincharon (14).
La comprobación de la existencia de las acuaporinas (15) o canales de agua ha creado un reto para poder definir su importancia en la biología celular y fisiología de todos los organismos. Tan grande ha sido el impacto del descubrimiento y estudio de estas proteínas que en el año 2003 se concedió el premio Nobel a Agre (14, 15), investigador que identificó por primera vez a estas proteínas y quien las ha estudiado en profundidad, obteniendo recientemente la estructura cristalina.
En las plantas, a diferencia de los demás organismos, las AQPs forman una gran familia génica que tiene una función primordial en los procesos biológicos de las plantas en general. Las acuaporinas son canales de agua, de naturaleza proteica, que facilitan y regulan el movimiento pasivo de agua siguiendo un gradiente decreciente de potencial hídrico (16). No obstante, el agua no es la única molécula que puede transitar a través de las acuaporinas, también solutos de bajo peso molecular como glicerol, urea, amonio, además del (CO2) que pueden atravesar estas proteínas (17).
TIPOS Y FUNCIONES DE LAS ACUAPORINAS EN LAS PLANTAS
Las acuaporinas pertenecen a una familia de grandes proteínas intrínsecas de membrana (MIP en inglés) y están presentes en animales, plantas y bacterias (18). En las características de sus secuencias se han establecido cuatro tipos de acuaporinas en plantas. Las dos más numerosas se localizan en la membrana plasmática (PIPs) o en el tonoplasto (TIPs). No obstante, existen otros dos tipos de acuaporinas, las homólogas a la nodulina 26 de soja (NIPs) y las pequeñas proteínas básicas e intrínsecas (SIPs) (19). La información actual sobre la localización y función de estos dos últimos tipos de acuaporinas es muy escasa, en contraste con la existente acerca de PIPs y TIPs (6). No obstante, en la literatura se describe la funcionalidad de cada uno de los tipos de acuaporinas (9).
1. Las PIPs, presentan una alta permeabilidad al agua, aunque con diferencias entre cada una de ellas. También permiten el paso de (CO2) y de glicerol.
2. Las TIPs, transportan agua a través de la membrana de la vacuola así como urea, glicerol y amonio, por lo que se les ha atribuido una función en la osmoregulación del citosol celular.
3. Para algunas NIPs, se ha demostrado que pueden transportar agua, aunque en menor cantidad que las PIPs, así como glicerol y otros solutos no cargados.
4. Las SIPs, se localizan en la membrana del retículo endoplásmico y algunas presentan también una cierta permeabilidad al agua (20).
Se han encontrado niveles elevados de expresión de genes de acuaporinas en tejidos con tasas importantes de flujo de agua a través de membranas, como es el caso de las regiones de crecimiento rápido, tallos, hojas y en las raíces, donde ocurre la captación del agua del suelo (7).
Otros estudios con plantas transgénicas que tienen inhibida la expresión de una o más PIPs presentan una capacidad menor de captación de agua por sus raíces (22, 23). Quedando claro que las acuaporinas tienen una función específica e importante en el control del transporte de agua célula a célula a través de los distintos tejidos de la planta y, por lo tanto, en la conductividad hidráulica de estos (23). No obstante, cuando las plantas crecen bajo condiciones de déficit hídrico, la transpiración es baja. El movimiento de agua en la planta se produce fundamentalmente a través de la llamada vía célula a célula, que es precisamente en la que las acuaporinas pueden regular el movimiento del agua a través de membranas, es por lo que es de esperar, que dichas proteínas tengan una función destacada regulando la conductividad hidráulica radical bajo condiciones de limitación hídrica (24).
En cualquier caso, a pesar de la investigación intensiva en este campo, la relación existente entre las AQPs y las respuestas de la planta al déficit hídrico permanece aun sin establecer y con resultados contradictorios (25, 26).
Actualmente, hay dos descripciones contrapuestas de la función de las acuaporinas en respuesta a estrés hídrico. La primera esta relacionada con la expresión de algunas acuaporinas, las cuales se inducen tras un estrés que produce deshidratación (27), lo que resultaría en una mayor permeabilidad al agua de las membranas y un transporte más fácil. La segunda se basa en el hecho de que la expresión de algunas AQPs es inhibida bajo condiciones de deshidratación, lo que debería de disminuir la permeabilidad al agua de membranas y puede permitir una conservación del agua de las células en esas condiciones (10, 28).
REGULACIÓN DE LAS RELACIONES HÍDRICAS DE LA PLANTA A TRAVÉS DE LA SIMBIOSIS MICORRÍZICA ARBUSCULAR.
La mayoría de las plantas terrestres establecen una asociación simbiótica con un grupo de hongos del suelo llamados hongos micorrízicos arbusculares (HMA). La simbiosis micorrízica arbuscular está presente en todos los ecosistemas naturales, incluso en aquellos con condiciones ambientales más adversas (29). Los estudios ecofisiológicos que han investigado la función de la simbiosis micorrizíca arbuscular (MA) en la protección de las plantas frente a la sequía, han demostrado que la simbiosis origina una alteración de la tasa de movimiento de agua dentro y hacia fuera de las plantas hospederas, con efectos sobre las relaciones hídricas y la fisiología de la plantas (30, 31). Por lo tanto, se acepta que la simbiosis micorrízica arbuscular protege a la planta hospedadora frente al déficit hídrico a través de una combinación de efectos físicos, nutritivos y celulares (32, 33).
A partir de los años 90 se comenzaron a desarrollar en Cuba trabajos que permitieron establecer las bases para el manejo de las asociaciones micorrízicas, partiendo de tres premisas fundamentales: la inoculación de especies eficientes de Hongos Micorrízicos Arbusculares (HMA), la importancia del ambiente edáfico en la selección de cepas eficientemente y la influencia del suministro de nutrientes sobre la efectividad de la simbiosis (34).
Debido a que las asociaciones micorrízicas desarrollan múltiples funciones entre las que se destacan: un aprovechamiento más eficiente de la zona radical a partir de un aumento en el volumen de suelo explorado, una mayor resistencia a las toxinas (metales pesados), incremento de la translocación y solubilización de elementos nutritivos esenciales, aumento de la tolerancia a condiciones abióticas adversas (seguía, salinidad, etc.), así como cierta protección contra patógenos (35).
Por otra parte, se ha demostrado que la infección micorrizógena de las raíces producen un efecto beneficioso en las relaciones hídricas de las plantas, lo que se manifesta a través de una mayor conductividad hidráulica de las raíces por unidad de masa seca; esto favoreció considerablemente la conductancia estomática y por tanto una mayor tasa fotosintética, por lo que se vio reflejado un mayor crecimiento en biomasa seca de la parte aérea de las plantas y en el área foliar (36).
Se ha demostrado que las micorrizas arbusculares (AM) en simbiosis puede proteger a las plantas contra los efectos perjudiciales del déficit de agua (37). Los estudios realizados hasta la fecha, han sugerido varios mecanismos por los que la simbiosis puede aliviar la sequía en las plantas hospederas. Los más importantes son: la absorción directa y la transferencia de agua a través de las hifas fúngicas de la planta huésped, cambios en las propiedades de retención de agua del suelo (38), mejor ajuste osmótico de las plantas micorrizadas, la mejora del intercambio de la planta de gases y uso eficiente del agua, la protección contra el daño oxidativo generado por la sequía (39).
La mejora de la tolerancia al estrés suele estar relacionado con la mejora del contenido de compuestos antioxidantes en las plantas. Dada la toxicidad de las especies reactivas de oxígeno (ROS) en las plantas necesitan disponer de sistemas adecuados de desintoxicación en el lugar que permita la eliminación rápida de estos compuestos. Estos sistemas incluyen varias enzimas antioxidantes y compuestas enzimáticas como el ascorbato, glutatión, los flavonoides, carotenoides y tocoferoles (40).
Investigaciones resientes realizadas en el cultivo del arroz han demostrado que el arroz se micorriza en condiciones aeróbicas, la cual se incrementa a medida que las plantas van creciendo (41, 42). La simbiosis micorrízica arbuscular incrementa en un 40% la eficiencia fotosintética de la planta de arroz bajo estrés hídrico inducido, la acumulación de glutatión como mecanismo antioxidante y reduce la acumulación de peróxido de hidrógeno y el daño oxidativo a los lípidos en estas plantas (42).
La micorrización es un buen ejemplo de las alteraciones morfológicas que las células de la raíz de las plantas deben acometer para acomodar la presencia de un simbionte. Dado que la mayoría de los cambios inducidos por la micorriza en las células de la raíz de la planta conciernen a las membranas citoplasmáticas o vacuolares (43), por lo que se propuso que la simbiosis originaría una alteración en los patrones de expresión de genes que codifican proteínas asociadas a membranas como es el caso de las acuaporinas. En el siguiente acápite se resume la información existente en la actualidad relativa a la alteración de genes que codifican acuaporinas por parte de la simbiosis micorrízica arbuscular.
REGULACIÓN DE ACUAPORINAS POR LA SIMBIOSIS MICORRÍZICA ARBUSCULAR Y SU RELACIÓN CON EL TRANSPORTE DE AGUA EN LAS PLANTAS.
Los primeros indicios de la alteración de la expresión de acuaporinas por parte de la simbiosis micorrízica arbuscular (MA) fueron propuestos en el año 2000, encontrándose la inducción de la expresión de acuaporinas tipo TIP en plantas de perejil y otras, por lo que se plantea que los genes de acuaporinas debido a la simbiosis micorrízica arbuscular serviría probablemente para optimizar el intercambio de nutrientes y agua entre ambos simbiontes (43). No obstante, estos dos estudios se realizaron bajo condiciones hídricas óptimas y no estudiaron la expresión de esas acuaporinas en plantas micorrizadas sometidas a déficit hídrico.
Otros estudios demuestran que el efecto de la disminución de la expresión del gen PIP NtAQP1 en plantas de tabaco mutantes antisentido micorrizadas para dicho gen (44), no afectó a la colonización de la raíz por parte de dos hongos micorrízicos arbusculares diferentes, lo que sugería que o bien la función del gen NtAQP1 no es relevante para el proceso de colonización radical o que la disminución de la expresión de este gen ha sido compensada por cambios en la abundancia o actividad de otras acuaporinas. Por el contrario, cuando se evaluó la eficiencia simbiótica de los dos hongos micorrízicos arbusculares, se observó que bajo condiciones de sequía, la eficiencia simbiótica de ambos hongos micorrízicos arbusculares era mayor en plantas de fenotipo salvaje que en las plantas antisentido y que el transporte de agua mediado por la acuaporina NtAQP1 es importante para la eficiencia de la simbiosis en condiciones de sequía (44).
Se han realizado investigaciones sobre el clonado de genes PIPs en soja y lechuga, donde se analizaron su patrón de expresión en plantas micorrizadas y no micorrizadas cultivadas en condiciones óptimas y de sequía (45). La hipótesis de partida era que si los hongos micorrízicos arbusculares pueden transferir agua a la raíz de la planta hospedadora, esperando que la planta incremente su permeabilidad al agua y que los genes de acuaporinas sean inducidos con el fin de permitir un flujo mayor transcelular de agua (23). En contra de esta hipótesis, los resultados obtenidos mostraron que los genes PIPs estudiados resultaban inhibidos por el déficit hídrico tanto en soja como en lechuga y que esa inhibición era aún más severa en las plantas micorrizadas que en las no micorrizadas (45). Un resultado similar fue obtenido para algunos genes PIP y TIP en raíces de plantas de tomate colonizadas y sometidas a salinidad (5). Por lo que se plantea que la disminución de la expresión de genes PIP en plantas micorrizadas en condiciones de déficit hídrico puede ser un mecanismo de regulación para limitar la perdida de agua de las células (45).
En los últimos años se ha encontrado que la respuesta de genes de acuaporinas a la simbiosis micorrízica arbuscular es bastante compleja, ya que un mismo gen de acuaporina responde de forma muy diferente dependiendo de las características intrínsecas del estrés osmótico aplicado y esa respuesta depende, a su vez, de la presencia o no del hongo micorrízico arbuscular en las raíces de la planta hospedadora (46, 47). La inducción o inhibición de genes de acuaporinas por sequía ha sido descrita en raíces de muchas especies de plantas (23, 44 y 45).
Recientemente, han detectado la inducción por la simbiosis micorrízica arbuscular de isoformas específicas de acuaporinas tipo PIP y NIP en Medicago truncatula (11). La acuaporina tipo PIP resultó muy permeable al agua, mientras que la tipo NIP mostró permeabilidad al amonio. Los autores han sugerido que ambas acuaporinas pueden estar implicadas en los procesos de intercambio (agua y amonio) entre el hongo y la planta, lo que incrementa el interés de estudiar las acuaporinas en la simbiosis micorrízica arbuscular (11).
La simbiosis micorrízica arbuscular modula la expresión de algunos genes de acuaporinas en la planta hospedadora, sin embargo, hasta ahora el número de genes de acuaporinas estudiados ha sido muy limitado y en plantas diferentes (32). Es por eso que se planta que para poder obtener conclusiones fiables sobre la función de las acuaporinas frente a condiciones ambientales particulares se debe de analizar el comportamiento de todas las isoformas de acuaporinas presentes en la planta (46). Por lo tanto, falta aún mucho por conocer acerca de la posible modulación por la simbiosis micorrízica arbuscular de la mayoría de los otros genes de acuaporinas y la repercusión que ello pueda tener sobre en variables como la conductividad hidráulica radical, las relaciones hídricas de las plantas y sobre la tolerancia al déficit hídrico de la planta hospedadora (32).
Investigadores de la Estación Experimental del Zaídín, Granada, España, han sido los primeros en impulsar estudios sobre la expresión de acuaporinas en plantas micorrizadas en condiciones de déficit hídrico. En primer lugar estudiaron el efecto de la disminución de la expresión del gen PIP NtAQP1 en plantas micorrizadas de tabaco mutantes antisentido para dicho gen (45), demostrando que el transporte de agua mediado por la acuaporina NtAQP1 es importante para mantener la eficiencia de la simbiosis en condiciones de sequía (44), por lo que la disminución de la expresión de genes PIP en plantas micorrizadas en condiciones de déficit hídrico puede ser un mecanismo de regulación para limitar la perdida de agua de las células (49).
Se ha demostrado que el ácido abscísico (ABA) modula la expresión de algunos genes PIP en raíces y hojas, además de modular tanto la expresión de acuaporinas como la propia conductividad hidráulica radical (27, 50, 51, 52). Estudios en plantas de tomate con un mutante deficiente en (ABA) indicaron que la micorrización reguló de forma diferente la expresión de cuatro genes PIP en condiciones de sequía y tras la aplicación de ABA exógeno, y además de que, esta regulación dependió del fenotipo de la planta estudiada y, por lo tanto, del contenido endógeno de ABA de la planta hospedadora (53, 54). Esto concuerda con resultados obtenidos con anterioridad, donde se encontró que genes PIP de arroz responden de forma diferente al déficit hídrico y al ABA. Indicando que durante el déficit hídrico la regulación de genes PIP sigue dos rutas, una dependiente de ABA y otra independiente de ABA (26). En su conjunto, estos resultados demostraron que la expresión de ciertos genes de acuaporina estarían reguladas tanto por ABA como por la simbiosis micorrízica arbuscular (26, 29, 53 y 54).
LAS ACUPORINAS EN LA PLANTAS DE ARROZ.
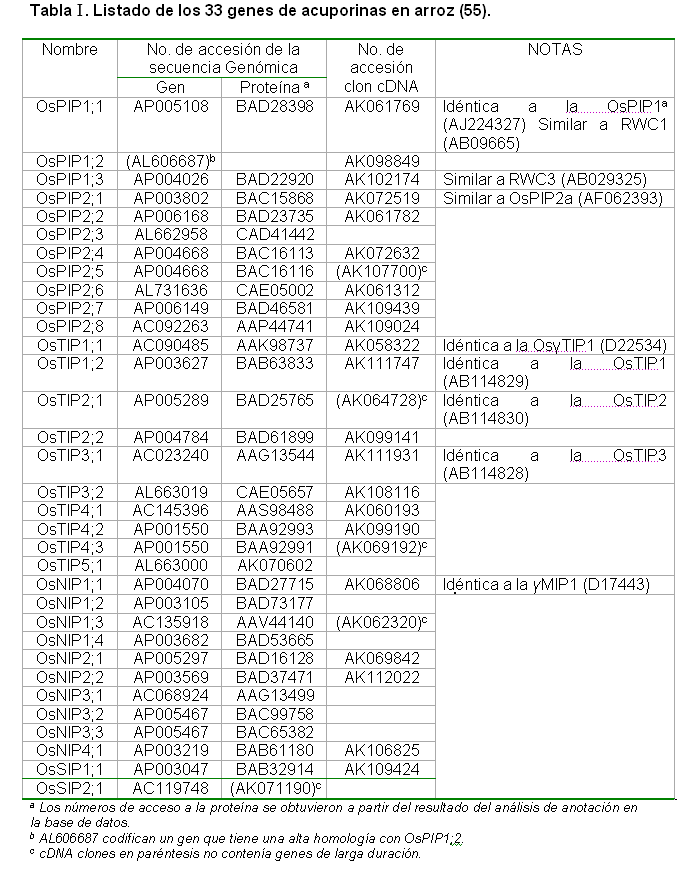
La literatura reporta que en el genoma del arroz existen las cuatro familias de acuaporinas (PIPs, TIPs, NIPs y las SIPs), nomenclatura que en el caso de este cultivo le anteceden las siglas Os, que significa el nombre de la especia (Oryza sativa), por lo tanto las acuaporinas llevan por nomenclatura (OsPIPs, OsTIPs, OsNIPs y OsSIPs) ver tabla 1 (55), en el resto de las plantas se sigue la misma nomenclatura.
En el genoma del arroz aparecen los 33 genes de acuaporinas los que están compuestos por 11 OsPIPs, 10 OsTIPs, 10 OsPINs y dos OsSIPs (Tabla I). Mediante el uso de transcripción inversa-PCR (RT-PCR) se demostró que en las plantas de arroz se expresan 12 genes PIPs y 10 TIPs en las raíces y en las hojas respectivamente (56).
Sin embargo, estos resultados de la localización específica de las acuaporinas y sus actividades de transporte de agua han sido erróneos, puesto que no se tuvo en cuenta las vías de transporte de agua célula – célula en las plantas y para eso se debió de utilizar anticuerpos de isoforma-específicas (57).
El transporte del agua en las plantas es muy dependiente de la expresión y actividad de los canales de transporte, llamados acuaporinas (12, 13, 14, 55 y 58), debido a que existen tejidos y células específicas en las que se localizan las diferentes familias de acuaporinas en plantas de arroz (57).
La mayoría de las plantas tienen más de 30 familias de acuaporinas: 35 en Arabidopsis (19), 33 en plantas de maíz (9), y 33 en el arroz (56, 57). Aunque se reportan que son 38, basándose en su secuencia de aminoácidos (59). La mayoría de los PIPs del arroz están situadas en la membrana plasmática, con la excepción notable de OsPIP1;2, que se supone que a de existir en el cloroplasto o las mitocondrias. Algunos de los genes aparecen estrechamente vinculados, como por ejemplo, los cuatro genes de tipo PIPs (OsPIP2;1, OsPIP2;4, OsPIP2;5, y OsPIP2;9) aproximadamente a 15,4 pares de Mb en el cromosoma 7, lo que sugiere eventos de duplicación de genes (60).
Estudio que se corroboró a través del uso de inmunotransferencia y inmunocitoquímica, utilizando siete isoformas específicas y anticuerpos de acuaporinas, observándose que en hoja y raíces existes una mayor expresión de las OsPIP1, OsPIP2;1, OsTIP1;1, y OsTIP2;2, mientras que las OsPIP2;3, OsPIP2;5 y OsTIP2;1 sólo en las raíces (58).
En las raíces, cantidades grandes de acuaporinas se acumulan en la región adyacente al ápice radical (en torno a 1,5 y 4 mm del ápice). En esta región, específica se localizan varias familias de las acuaporinas OsPIP1 y OsTIP2;2, predominantemente en la endodermis y el cilindro central, respectivamente. OsTIP1;1 mostró localización específica en la rhizodermis y exodermis. Se observó que en toda la raíz se acumulan las OsPIP2;1, OsPIP2;3 y OsPIP2;5, con mayor porcentaje de acumulación en la endodermis que en otras células. En la región apical de la raíz a 35 mm, donde se desarrolla el tejido aerénquima, se observó niveles bajo de acumulación de acuaporinas. En hojas, las familias OsPIP1:1 y OsPIP2;1 se ubicaron principalmente en las células del mesófilo. Las OsPIP2;2, OsPIP2,3, OsPIP2;5 y OsTIP2;2, expresados en levaduras mostraron alta actividad de transporte de agua en esta región (55).
Se ha observado que existe una estrecha relación entre los genes de acuaporinas del trigo y los del arroz, específicamente en las acuaporinas PIPs y las TIPs (61), los cuales forman una gran familia con características muy similares, que se consideran importantes en la estructura y función.
Otra investigación se llevó a cabo en condiciones de hidropónico con estrés hídrico, simulado mediante la adición de Polyethilene Glycol un déficit hídrico, con el fin de demostrar la participación de las acuaporinas en la asimilación de nitrógeno en forma de amonio y nitrato (NH4+, NO3-) respectivamente, su influencia en el crecimiento y desarrollo de las plantas de arroz, así como su tolerancia al estrés hídrico. Los resultados mostraron que, en comparación con las plantas no estresadas, el masa fresca de las plántulas de arroz aumentó un 14% con la nutrición NH4+, mientras que había disminuido en un 20% con cualquiera de NO3- o con la nutrición nitrogenada mixta, mientras que el flujo del xilema disminuyó en un 57% y 24% bajo estrés hídrico en NO3- y la nutrición mixta y conductividad hidráulica raíz disminuyó un 29% y 54% en las plantas en NH4+ y NO3- condiciones de nutrición, respectivamente. Resultado similares fueron encontrados con anterioridad (62, 63). Aunque la capacidad de absorción de agua disminuyó en ambos nutrientes, la actividad acuaporinas fue mayor en NH4+ que en NO3-, independientemente de estrés hídrico (64). Comportamiento que puede mejorar el manejo del agua en las plántulas de arroz y posteriormente aumentar su resistencia a la seguía (64).
Las acuaporinas están relacionadas con la inhibición de la absorción de agua, inhibida por las bajas temperaturas en la raíz (65, 66, 67, 68). Los niveles de expresión de estos genes en las raíces de arroz disminuyeron notablemente a baja temperatura (4ºC) durante 3 días, excepto OsPIP1;3, que aumentaron (55). Resultados similares fueron reportados en otras variedades de arroz (68, 69) y también en el maíz (67). Estudios posteriores a este, revelaron que la actividad de transporte de agua de las acuaporinas es regulada a través del mecanismo de compuerta, como la fosforilación de residuos específicos de serina (71).
A modo de conlución, las acuaporinas estan presente en todos los sere vivos y son las proteinas que regulan el transporte del agua en la membrana, además intracelular y extracelular, así como de otros compuestos, su comportamiento es regulado por los estreses biotico y abioticos.
En el caso de las plantas agregar que la simbiosis micorrízica influye y puede determinar el funcionamiento de las acuaporinas bajo diversas condiciones estresantes en cultivos como el maíz, arabidosis, tabaco, arroz, entre otros. Especificamente en las plantas de arroz no se reporta variación en la expreción de las acuporinas por efecto de las micorrizas, sin embargo se a demostrado su expresión tanto en hojas como en raíz, su localización a nivel de tejidos y posición, su funcionalidad con diferentes formas nitrogenadas aplicadas, así como su inhibición cuando imperan bajas temperaturas en las raíces del arroz.
REFERENCIAS
1. González, S. Las acuaporinas o quién capta y distribuye el agua en las plantas. [En línea] Curso avanzado de Bioquímica del Suelo, 2007. [Consultado: 01/2009]. Disponible en: <http://www.madrimasd.org/blogs/universo/2007/03/09/60821>
2. Seki, M, Kamei, A, Yamaguchi-Shinozaki, K, Shinozaki, K. Molecular responses to drought, salinity and frost: common and different paths for plant protection. Curr. Op. Biotechnol., 2003, vol. 14, p.194-199.
3. Bray, EA. Genes commonly regulated by water-deficit stress in Arabidopsis thaliana. J. Exp. Bot., 2004, vol 55, p.2331-2341.
4. Denby K, Gehring, C. Engineering drought and salinity tolerance in plants: lessons from genome-wide expression profiling in Arabidopsis. Trends Biotechnol., 2005, vol. 23, p.547-552.
5. Ouziad, F, Wilde, P, Schmelze, E, Hildebrandt, U, Bothe, H. Analysis of expression of aquaporins and Na+/H+ transporters in tomato colonized by arbuscular mycorrhizal fungi and affected by salt stress. Environ. Exp. Bot., 2006, vol. 57, p. 177-186.
6. Luu, D. T, Maurel, C. Aquaporins in a challenging environment: molecular gears for adjusting plant water status. Plant, Cell Environ. 2005. vol. 28, p.85-96.
7. Zhao, C-X, Shao, H-B and Chu, L-Y. Aquaporin structure-function relationships: water flow through plant living cells. Colloid Surface. 2008, vol. 62, p.163-172.
8. Otto, B, Kaldenhoff, R. Cell-specific expression of the mercury-insensitive plasma-membrane aquaporin NtAQP1 from Nicotiana tabacum. Planta. 2000, vol. 211, p.167-172.
9. Maurel, C. Plant aquaporins: novel functions and regulation properties. FEBS Lett. 2007, vol.581, p.2227-2236.
10. Kaldenhoff, R, Fischer, M. Functional aquaporin diversity in plants. Biochimica and Biophysica Acta. 2007, vol.1758, p.1134-1141.
11. Pettersson, N, Hagström, J, Bill, RM, Hohmann, S. Expression of heterologous aquaporins for functional analysis in Saccharomyces cerevisiae. Curr. Genet. 2006, vol.50, p.247-255.
12. Uehlein, N, Fileschi, K, Eckert, M, Bienert, GP, Bertl, A, Kaldenhoff, R. Arbuscular mycorrhizal symbiosis and plant aquaporin expression. Phytochemistry. 2007, vol.68, p.122-129.
13. Vera–Estrella, R., Barkla, B. and Pantoja, O. La biología y la fisiología de las acuaporinas en plantas.[En línea] Biojournal, 2006, vol.1, p.1–15. [Consultado: 30/1/2010] Disponible en: <http://www.biojournal.net/pdf/05.pdf.Biojournal.net>
14. Daniels, MJ, Wood, MR, Yeager, M. In vivo functional assay of a recombinant aquaporin in Pichia pastoris. Appl. Environ. Microbiol. 2006, vol. 72, p. 1507-1514.
15. Knepper, MA, Nielsen, S. Peter, Agre, 2003 Nobel Prize winner in chemistry. Journal of the American Society of Nephrology. 2004, vol.15, p.1093-1095.
16. Agre, P., Sasaki, S., Chrispeels, MJ. Aquaporins: a family of water channel proteins. American. Journal of Physiology, 1993, vol. 261, f 461.
17. 17 Kruse, E, Uehlein, N, Kaldenhoff, R. The aquaporins. Genome Biol. 2006, vol.7, p. 206. ISSN:1465-6914
18. Flexas, J, Ribas-Carbó, M, Hanson, DT, Bota, J, Otto, B, Cifre, J, McDowell, M., Medrano, H., Kaldenhoff, R. Tobacco aquaporin NtAQP1 is involved in mesophyll conductance to CO2 in vivo. Plant J. 2006, vol.48, p.427-439.
19. Maurel, C. Aquaporins and water permeability of plant membranes. An. Rev. Plant Physiol. Plant Mol. Biol. 1997, vol.48, p.399-429.
20. Johanson, U., Karlsson, M., Johanson, I., Gustavsson, S., Sjövall, S., Fraysse, L., Weigh, AR, Kjellbom, P. The complete set of genes encoding major intrinsic proteins in Arabidopsis provides a framework for a new nomenclature for major intrinsic proteins in plants. Plant Physiol. 2001, vol.126, p.1358-1369.
21. Ishikawa, F., Suga, S., Uemura, T., Sato, MH, Maeshima, M. Novel type aquaporin SIPs are mainly localized to the ER membrane and show cell-specific expression in Arabidopsis thaliana. FEBS letters, 2005, vol.579, no.25, p.5814-5820.
22. Siefritz, F., Tyree, M.T., Lovisolo, C., Schubert, A. and Kaldenhoff, R. PIP1 plasma membrane aquaporins in tobacco: from cellular effects to function in plants. Plant Cell. 2002, vol.14, p.869-876.
23. Javot, H. and Maurel, C. The role of aquaporins in root water uptake. Ann. Bot. 2002, vol.90, p.301-313.
24. Hachez, C., Moshelion, M., Zelazny, E., Cavez, D., Chaumont, F. Localization and quantification of plasma membrane aquaporin expression in maize primary root: a clue to understanding their role as cellular plumbers. Plant Mol. Biol. 2006, vol. 62, p.305-323.
25. Aharon, R., Shahak, Y., Wininger, S., Bendov, R., Kapulnik, Y., Galili, G. Overexpression of a plasma membrane aquaporins in transgenic tobacco improves plant vigour under favourable growth conditions but not under drought or salt stress. Plant Cell. 2003, vol.15, p.439-447.
26. Lian, HL, Yu, X, Ye, Q, Ding, XS, Kitagawa, Y, Swak, SS, Su, WA, Tang, ZC. The role of aquaporin RWC3 in drought avoidance in rice. Plant Cell Physiol. 2004, vol.15, p.481. 489.
27. Jang, JY, Kim, DG, Kim, YO, Kim, JS, Kang, H. An expression analysis of a gene family encoding plasma membrane aquaporins in response to abiotic stresses in Arabidopsis thaliana. Plant Mol. Biol. 2004, vol. 54, p.713-725.
28. Smart, LB, Moskal, WA, Cameron, KD, Bennett, AB. Mip genes are down-regulated under drought stress in Nicotiana glauca. Plant Cell Physiol. 2001, vol.42, p. 686-693.
29. Smith, S.E., Read, D.J. Mycorrhizal symbiosis. 3ra. ed. San Diego, CA: Academic Press, 2003. 3v, 787 p.
30. Ruiz-Lozano, JM. Arbuscular mycorrhizal symbiosis and alleviation of osmotic stress. New perspectives for molecular studies. Mycorrhiza. 2003, vol.13, p. 309-317.
31. Ruiz-Lozano, JM, Porcel, R, Aroca, R. Does the enhanced tolerance of arbuscular mycorrhizal plants to water deficit involve modulation of drought-induced plant genes? New Phytol. 2006, vol.171, p.693-698.
32. Ruiz-Lozano, JM, Aroca, R. Modulation of aquaporin genes by the arbuscular mycorrhizal symbiosis in relation to osmotic stress tolerance. In: Seckbach J, Grube M. (Eds.) Symbioses and Stress: Joint Ventures in Biology, cellular origin, life in extreme habitats and astrobiology. Springer Science Business Media, 2010, v.17, part.4, p.359-374.
33. Porcel, R, Aroca, R, Cano, C, Bago, A, Ruiz-Lozano, J.M. A gene from the arbuscular mycorrhizal fungus Glomus intraradices encoding a binding protein is up-regulated by drought stress during the mycorrhizal symbiosis. Environ. Exp. Bot. 2007, vol.60, p.251-256.
34. Rivera-Espinosa, R.A. and Fernández-Suárez, K. El manejo eficiente de la simbiosis micorrízica, una vía hacia la agricultura sostenible, estudio de caso: El Caribe. La Habana: Instituto Nacional de Ciencias Agrícolas (INCA), 2003. 171 p. ISBN 959-7023-24-5.
35. Fernández, F. Avances en la producción de inoculantes micorrízicos arbusculares. En: Rivera-Espinosa, R.A. and Fernández-Suárez, K. (Eds.) El manejo eficiente de la simbiosis micorrízica, una vía hacia la agricultura sostenible Estudio de caso: El Caribe. La Habana: Instituto Nacional de Ciencias Agrícolas (INCA), 2003. p 97-110. ISBN 959-7023-24-5.
36. Dell'amico, J., Torrecillas, A., Rodríguez, P., Morte, A., Sánchez-Blanco, M.J. Responses of tomato plants associated with the arbuscular mycorrhizal fungus Glomus clarum during drought and recovery. Journal of Agricultural Science. 2002, vol.138, no.4, p.387-393.
37. Augé RM. Arbuscular mycorrhizae and soil/plant water relations. Can J Soil Sci. 2004. vol.84, p.373-81.
38. Augé, RM. Water relations, drought and vesicular-arbuscular mycorrhizal symbiosis. Mycorrhiza. 2001, vol.11, p.3-42.
39. Porcel, R. and Ruiz-Lozano, JM. Arbuscular mycorrhizal influence on leaf water potential, solute accumulation and oxidative stress in soybean plants subjected to drought stress. J. Exp.Bot. 2004, vol.55, p.1743-1750.
40. Ma, YH, Ma, FW, Zhang, JK, Li, MJ, Wang, YH, Liang, D. Effects of high temperature on activities and gene expression of enzymes involved in ascorbate-glutathione cycle in apple leaves. Plant Sci. 2008, vol.175, p.761-766.
41. Vallino, M., Greppi, D., Novero, M., Bonfante, P., Lupotto, E. Rice root colonisation by mycorrhizal and endophytic fungi in aerobic soil. Annals of Applied Biology. 2009. vol.154, no.2, p.195-204.
42. Ruiz-Sánchez, M., Aroca, R., Muñoz, Y., Polón, R., Ruiz-Lozano, J.M. The arbuscular mycorrhizal symbiosis enhances the photosynthetic efficiency and the antioxidative response of rice plants subjected to drought stress. Journal of Plant Physiology. 2010, vol.167, p.862–869.
43. Krajinski, F, Biela, A, Schubert, D, Gianinazzi-Pearson, V, Kaldenhoff, R, Franken P. Arbuscular mycorrhiza development regulates the mRNA abundance of Mtaqp1 encoding a mercury-insensitive aquaporin of Medicago truncatula. Planta. 2000, vol. 211, p.85-90.
44. Porcel, R, Gómez, M, Kaldenhoff, R, Ruiz-Lozano, JM. Impairment of NtAQP1 gene expression in tobacco plants does not affect root colonization pattern by arbuscular mycorrhizal fungi but decreases their symbiotic efficiency under drought. Mycorrhiza. 2005, vol.15, no.6, p.417-23
45. Porcel, R, Aroca, R, Azcón, R, Ruiz-Lozano, JM. PIP aquaporin gene expression in arbuscular mycorrhizal Glycine max and Lactuca sativa plants in relation to drought stress tolerance. Plant Mol. Biol. 2006, vol.60, p.389-404.
46. Jahromi, F, Aroca, R, Porcel, R, Ruiz-Lozano, JM. Influence of salinity on the in vitro development of Glomus intraradices and on the in vivo physiological and molecular responses of mycorrhizal lettuce plants. Microb. Ecol. 2008, vol.55, no.1, p.45-53.
47. Aroca, R, Porcel, R, Ruiz-Lozano, JM. How does arbuscular mycorrhizal symbiosis regulate root hydraulic properties and plasma membrane aquaporin in Phaseolus vulgaris under drought, cold or salinity stresses? New Phytol. 2007, vol.173, p.808-816.
48. Alexandersson, E, Fraysse, L, Sjöval-Larsen, S, Gustavsson, S, Fllert, M, Karlsson, M, Johanson, U, Kjelbom, P. Whole gene family expression and drought stress regulation of aquaporins. Plant Mol. Biol. 2005, vol.59, p.469-484.
49. Porcel, R, Aroca, R, Azcón, R, Ruiz-Lozano, JM. PIP aquaporin gene expression in arbuscular mycorrhizal Glycine max and Lactuca sativa plants in relation to drought stress tolerance. Plant Mol. Biol. 2006, vol.60, p.389-404.
50. Zhu, C, Schraut, D, Hartung, W, Schäffner, AR. Differential responses of maize MIP genes to salt stress and ABA. J. Exp. Bot. 2005, vol.56, p.2971-2981.
51. Aroca, R, Ferrante, A, Vernieri, P, Chrispeels, MJ. Drought, abscisic acid, and transpiration rate effects on the regulation of PIP aquaporin gene expression and abundance in Phaseolus vulgaris plants. Ann. Bot. 2006, vol.98, p.1301-1310.
52. Lian, HL, Yu, X, Lane, D, Sun, WN, Tang, ZC, Su, WA. Upland rice and lowland rice exhibit different PIP expression under water deficit and ABA treatment. Cell Res. 2006, vol.16, p.651-660.
53. López-García, F, Aroca, R, Alguacil, MM, Vernieri, P, Ruiz-Lozano, JM. Plant responses to drought stress and exogenous ABA application are differently modulated by mycorrhization in tomato and an ABA-deficient mutant (sitiens). Microb. Ecol. 2008, vol.56, no.4, p.704-719.
54. Aroca, R. Exogenous catalase and ascorbate modify the effects of abscisic acid (ABA) on root hydraulic properties in Phaseolus vulgaris L. plants. J. Plant Growth Regul. 2006, vol.25, no.1, p.0-17.
55. Sakurai, J., Ishikawa, F., Yamaguchi, T., Uemura, M. and Maeshima, M. Identification of 33 rice aquaporin genes and analysis of their expression and function. Plant Cell Physiol. 2005, vol.46, p.1568–1577.
56. Katsuhara, M., Hanba, Y., Shiratake, K. and Maeshima, M. Expanding roles of plant aquaporins in plasma membranes and cell organelles. Funct. Plant Biol. 2008, vol.35, p.1–14.
57. Guo, L, Wang, ZY, Lin, H, Cui, WE, Chen, J, Liu, M, Chen, ZL, Qu, LJ, Gu, H. Expression and functional analysis of the rice plasma membrane intrinsic protein gene family. Cell Res. 2006, vol.16, p.277–286.
58. Sakurai, J., Ahamed, A., Mari, M., Masayoshi, M. and Matsuo, U. Tissue and Cell-Specific Localization of Rice Aquaporins and Their Water Transport Activities. Plant Cell Physiol. 2008, vol.49, no.1, p.30–39.
59. Chaumont, F., Barrieu, F., Wojcik, E., Chrispeels, M.J. and Jung, R. Aquaporins constitute a large and highly divergent protein family in maize. Plant Physiol. 2001, vol.125, p.1206–1215.
60. Forrest, K.L. and Bhave, M. Major intrinsic proteins (MIPs) in plants: a complex gene family with major impacts on plant phenotype. Funct. Integr. Genomics. 2007, vol.7, p.263 – 289.
61. Forrest, K. L.; Bhave Mrinal. Physical mapping of wheat aquaporin genes. Theor Appl Genet. 2010, vol.120, p.863–873.
62. Guo, S, Chen, G, Zhou, Y, Shen, Q. Ammonium nutrition increases photosynthesis rate under water stress at early development stage of rice (Oryza sativa L.). Plant Soil. 2007, vol.296, p.115–124.
63. Guo, S, Zhou, Y, Li, Y, Gao, Y, Shen, Q. Effects of different nitrogen form and water stress on water use efficiency of rice plants. Ann Appl Biol. 2008, vol.153, p.127–134.
64. Gao, Y; Li, Y; Xiuxia, Y.; Haijun, L.; Qirong, S. & Guo, S. Ammonium nutrition increases water absorption in rice seedlings (Oryza sativa L.) under water stress. Plant Soil. 2010, vol.331, p.193–201.
65. Lee, S.H., Chung, G.C. and Steudle, E. Gating of aquaporins by low temperature in roots of chilling-sensitive cucumber and chillingtolerant figleaf gourd. J. Exp. Bot. 2005, vol.56, p.985–995.
66. Lee, S.H., Chung, G.C. and Steudle, E. Low temperature and mechanical stresses differently gate aquaporins of root cortical cells of chilling-sensitive cucumber and -resistant figleaf gourd. Plant Cell Environ. 2005, vol.28, p. 191–1202.
67. Aroca, R., Amodeo, G., Fernández-Illescas, S., Herman, E.M., Chaumont, F. and Chrispeels, M.J. The role of aquaporins and membrane damage in chilling and hydrogen peroxide induced changes in the hydraulic conductivity of maize roots. Plant Physiol. 2005, vol.137, p.341– 353.
68. Yu, X., Peng, Y.H., Zhang, M.H., Shao, Y.J., Su, W.A. and Tang, Z.C. Water relations and an expression analysis of plasma membrane intrinsic proteins in sensitive and tolerant rice during chilling and recovery. Cell Res. 2006, vol.16, p.599–608.
69. Murai-Hatano, M., Kuwagata, T., Sakurai, J., Nonami, H., Ahamed, A., Nagasuga,K., Matsunami, T. Effect of Low Root Temperature on Hydraulic Conductivity of Rice Plants and the Possible Role of Aquaporins. Plant and Cell Physiology. 2008, vol.49, no.9, p.1294-1305.
70. Matsumoto, T., Lian, H.-L., Su, W.-A., Tanaka, D., Liu, C.W., Iwasaki, I., Kitagawa, Y. Role of the Aquaporin PIP1 Subfamily in the Chilling Tolerance of Rice. Plant and Cell Physiology. 2009, vol.50, no.2, p.216-229.
71. Törnroth-Horsefield, S., Wang, Y., Hedfalk, K., Johanson, U., Karlsson, M., Tajkhorshid, E., Neutze, R. and Kjellbom, P. Structural mechanism of plant aquaporin gating. Nature. 2006, vol.439, p.688–694.
Recibido 02/07/2010, aceptado 27/05/2011.

{kind=link}