










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCultivos Tropicales
versión impresa ISSN 0258-5936
cultrop vol.33 no.1 La Habana ene.-mar. 2012
EFECTO DE LA 24-EPIBRASINÓLIDA EN ELCRECIMIENTO, LOS NIVELES DE PROLINA Y DE MALONDIALDEHIDO DE PLÁNTULAS DE ARROZ (Oryza sativa L.) SOMETIDAS A ESTRÉS SALINO
Effect of 24-epibrassinolide on growth, proline and malondialdehide levelsin rice seedlings (Oryza sativa L.) under salt stress
Yanelis Reyes Guerrero, I Lissy Rosabal Ayan,I LisbelMartínez González,I Dr.C. Luis M. Mazorra Morales,I Dra.C. Miriam Núñez Vázquez,I Dr. Alejandro PietersII
I Departamento de Fisiología y Bioquímica Vegetal, Instituto Nacional de Ciencias Agrícolas, gaveta postal 1, San José de las Lajas,Mayabeque, CP 32700.
II Investigador Asociado delLaboratorio de Ecofisiología Vegetal, Centro de Ecología. InstitutoVenezolano de Investigaciones Científicas, carretera Panamericana,km 11, Altos de Pipe, Estado de Miranda, Venezuela.
RESUMEN
La salinización de los suelos es uno de los fenómenos más alarmantes a nivel mundial, por lo que es de gran importancia encontrar productos naturales que induzcan tolerancia al estrés salino. En este trabajo se evaluó el efecto de la 24-epibrasinólida (24-EBL), un brasinoesteroide natural, en el crecimiento, los niveles de prolina y la peroxidación lipídica en plántulas de arroz (Oryza sativa L.) var. J-104, (susceptible) sometidas a estrés por NaCl durante siete días. Para ello, las semillas de arroz se sumergieron en agua destilada durante 24 horas. Posteriormente se germinaron en placas Petri, a las 48 horas se trasplantaron a potes con suelo Hidromórfico Gley Nodular Ferruginoso, los cuales se colocaron en una cesta plástica con solución nutritiva Hoagland diluida para propiciar el crecimiento normal de las plantas. Al emerger la tercera hoja verdadera (5-6 semanas después del trasplante) se impuso el tratamiento de salinidad (100mM de NaCl en solución nutritiva de Hoagland) por 7 días y en este mismo momento se realizó la aspersión foliar de 24-EBL (0.5, 1 y 2 µmolesL-1). Finalmente las plantas se mantuvieron en solución nutritiva de Hoagland por 14 días para su recuperación. Los muestreos de la longitud y la biomasa del sistema radical y de la parte aérea se realizaron al iniciar el tratamiento salino, al finalizar el período de estrés, a los siete días de recuperación y al finalizar este último período. Además se midieron los niveles de prolina y de malondialdehído (MDA), medida de la peroxidación lipídica, al finalizar la recuperación. Se evidenció que tanto el efecto del tratamiento con NaCl como el de la aspersión foliar con EBL en el crecimiento de las plántulas y la concentración de MDA foliar dependió de las temperaturas en las cuales estaban creciendo las mismas. El incremento que el estrés por NaCl produjo en el nivel de prolina foliar a los 14 días de la recuperación disminuyó significativamente por la aplicación exógena de 1 y 2 µmolesL-1 de EBL en ambas repeticiones.
Palabras clave: brasinoesteroides, estrés salino, arroz.
ABSTRACT
Salinization of soils is one of the most alarmingfacts worldwide, it remarks the necessity of find naturalproducts which omdice the saline stress tolerance on plants.On this paper, the effect of 24 epibrasinolide (EBL), a naturalbrasinoesteroid, on growth, proline level and lipidycperoxidation on rice plantlets, var. J-104 (susceptible) understress caused NaCl during seven days, was evaluated. Riceseeds were sowing on distilled water during 24 hours, thenwere germinated on Petri dishes and 48 hours later, were plantedon flasks containing a Gley nodular ferritic hydromorphic soil,whichs were placed on a plastic basket containing Hoaglandnutritive solution, to allow the normal growth of plants. At themoment of the appearance of the 3th true live (5 to 6 weeksafter planting), the treatment of salinity was imposed with NaCI(100 mM) on Hoagland solution during 7 days and in thismoment leaves were sprinkled with 24 epibrasinolide (0,5; 1and 2 uM/L-1). Plants were maintained on Hoagland solutionduring 14 days to their recovering. At the moment of the salinitytreatment application, at the end of the stress period, 7 daysafter beginning of recovering period and at the end of thisperiod, the length and biomass of radical and aereal parts ofplants were measured. Proline and malondialdehyde levelswere also evaluated, as well as lipidyc peroxidation at the endof the recovering period. The results showed that the effectsof NaCI and EBL on the growth and malondialdehyde level onleaves of plantlets, depends on the temperature during thegrowing period. The increase of proline levels, caused by thestress due to NaCI after 14 days of recovering, diminishedsignificatively with the application of 1 to 2 uML-1 de EBL onboth repetitions.
Key words: brasssinosteroids, salt stress, rice.
INTRODUCCIÓN
La salinidad de los suelos se ha convertido en uno de los problemas más alarmantes en la actualidad. Las frecuentes variaciones del clima debido al cambio climático, la erosión en zonas costeras que permite la entrada del agua del mar, así como la inadecuada irrigación de los suelos, hacen que este fenómeno se agrave cada día más [1-3]. Existen reportes que demuestran que el área de nuestro planeta afectada por la salinización es de alrededor de 800 millones de hectáreas [4]. En nuestro país, la superficie agrícola está afectada en un 14% y otro 15% más presenta peligros potenciales de salinización [5]. Este fenómeno tiene una incidencia directa en la producción de alimentos y en la economía del agricultor que vive en dichas zonas [3].
El arroz (Oryza sativa L.) es una de las especies más cultivadas en el planeta pues constituye la fuente principal de alimento de más de un tercio de la población mundial. Este cultivo, por sus características culturales, ve seriamente afectada su productividad por la salinidad en cada etapa del desarrollo, especialmente en la etapa de plántula, cuando es más susceptible al estrés salino [6].
Los brasinoesteroides son potentes reguladores del crecimiento vegetal de naturaleza esteroidal. Estas hormonas tienen efectos pleiotrópicos como son: estimulación del alargamiento celular y de la desdiferenciación de protoplastos, la regeneración de la pared celular, la regulación de la diferenciación de elementos traquearios, el incremento de la biomasa y del rendimiento [7].
Además, se ha informado ampliamente el efecto protector de los brasinoesteroides ante distintos tipos de estrés como altas y bajas temperaturas [8], sequía y salinidad [9]. Además disminuyen los efectos del estrés causado por la falta de nutrientes y por exceso de metales pesados y también incrementan la resistencia a herbicidas y a agentes patógenos [7].
Específicamente en arroz, existen algunos estudios sobre la protección de los brasinoesteroides ante el estrés salino.
Anuradha y Rao han informado una recuperación en el crecimiento y en algunos aspectos metabólicos en semillas de arroz tratadas con EBL [10; 11]. Otros autores han reportado que el tratamiento con EBL aumentó el crecimiento de plántulas de arroz sometidas a estrés salino y disminuyó la peroxidación lipídica y la acumulación de prolina causada por este estrés [12].
Se ha demostrado, además, que el tratamiento por siete días con EBL incrementó las velocidades de crecimiento de las raíces y la parte aérea de plántulas de arroz en medio salino [13].
Como se puede apreciar, todos estos autores refieren la utilización del tratamiento a las semillas para sus evaluaciones. Sin embargo, la información que aparece en la literatura internacional acerca de la posible protección de los brasinoesteroides cuando es utilizada la aspersión foliar es bastante limitada.
Por estas razones, se decidió acometer el siguiente trabajo, cuyo objetivo central fue evaluar el efecto de la aspersión foliar con 24-epibrasinólida (24-EBL) en el crecimiento, los niveles de prolina y la peroxidación lipídica en plántulas de arroz var. J-104 sometidas a un período corto de estrés por NaCl.
MATERIALES Y MÉTODOS
Para dar cumplimiento al objetivo propuesto, se utilizaron semillas de arroz (Oryza sativa L.) de la variedad Jucarito-104 (J-104) considerada como susceptible al estrés salino [14] y se sumergieron en agua destilada durante 24 horas. Posteriormente, se colocaron en placas Petri; en la oscuridad, para propiciar la germinación y a las 48 horas, las semillas germinadas se trasplantaron a potes que contenían suelo Hidromórfico Gley Nodular Ferruginoso [15] y se introdujeron en cestas plásticas que contenían agua corriente. Se dejaron crecer 3 plantas por pote y se colocaron 20 potes por cesta. Las cestas se colocaron dentro de un umbráculo para evitar el efecto de las precipitaciones. En el momento de emergencia de la tercera hoja verdadera (19 días después del trasplante), se realizó la aspersión foliar con la 24-epibrasinólida (EBL). Las concentraciones utilizadas fueron 0.5, 1 y 2 µmolesL-1 . Se asperjaron 120 plantas por tratamiento y se dejaron 120 como tratamiento control. En ese momento, las plantas se dividieron en dos grupos de 60 plantas, uno se colocó en cestas a las cuales se les adicionó solución nutritiva Hoagland diluida y el otro se colocó en cestas que contenían solución nutritiva Hoagland diluida suplementada con 100 mmolesL-1 de NaCl. A los siete días, las plantas que estaban sometidas a tratamiento salino, se colocaron en solución nutritiva Hoagland diluida, durante 14 días, para evaluar la recuperación.
Las evaluaciones de crecimiento (altura, longitud de las raíces, masa seca de la parte aérea y de las raíces) se realizaron a 12 plantas por tratamiento, al iniciar y finalizar el tratamiento salino y a los siete y 14 días de recuperación. La longitud del tallo se midió con una regla desde la base del mismo al ápice de la hoja más alargada. De igual forma se determinó la longitud de la raíz.
En el caso de las masas secas, con las medias de cada tratamiento a los 0, 7 y 14 días de recuperación, se calcularon los porcentajes de variación que el tratamiento con NaCl provocó en estos indicadores utilizando la fórmula:
%= -(1 – T2/T1)*100
T2= Valor de masa seca de cada tratamiento en condiciones salinas
T1= Valor de masa seca de cada tratamiento en solución nutritiva
Las concentraciones de prolina y de malondialdehido (MDA) en las hojas se determinaron al finalizar el período de recuperación. La prolina se realizó mediante el método de Bates et al. [16] y el contenido de MDA se determinó por el método del ácido tiobarbitúrico [17]. En ambos casos se realizaron 3 extracciones con 3 réplicas cada una por tratamiento.
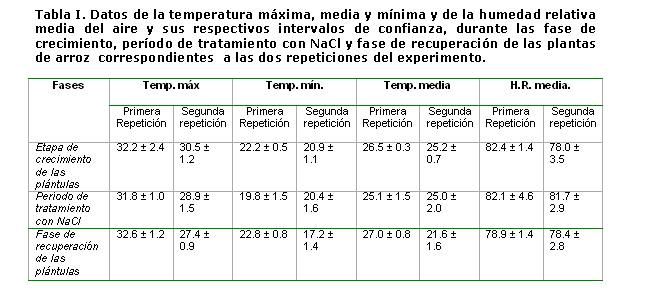
Se ejecutaron dos repeticiones del experimento, una se inició el 8 de septiembre y la otra el 12 de octubre del 2009, respectivamente. Se registraron las temperaturas mínimas, medias y máximas y las humedades relativas medias durante la etapa de crecimiento inicial de las plántulas (desde el trasplante hasta la imposición del tratamiento con NaCl), durante el período de tratamiento con NaCl (siete días) y durante la fase de recuperación (14 días) en las dos repeticiones del experimento (Tabla I).
Los datos de cada repetición se procesaron mediante el cálculo de las medias y los intervalos de confianza a p=0.05.
RESULTADOS Y DISCUSIÓN
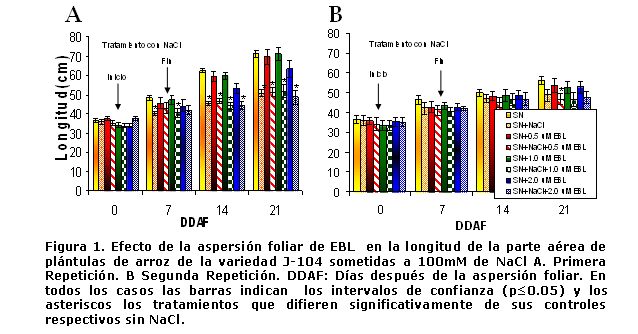
En la figura 1 se muestran los resultados de la longitud de la parte aérea de ambas repeticiones.
Como se puede apreciar en la primera repetición del experimento, el tratamiento con NaCl afectó la longitud de la parte aérea; aunque la 24-epibrasinólida (EBL), a las concentraciones de 0,5 y 2 µmolesL-1, tuvo un efecto protector durante el período de estrés. Este efecto del estrés salino en el crecimiento es ampliamente conocido ya que tanto el estrés hídrico como la acumulación de iones retrasan los procesos de división y diferenciación celular [18]. Además se ha demostrado que en condiciones salinas disminuye la síntesis de proteínas, ARN y ADN [19]. Lo anteriormente planteado unido a la síntesis de solutos compatibles para restablecer el potencial osmótico celular, estrategia costosa para la célula, hacen que se inhiban los procesos de crecimiento. Además se observa que en la recuperación en solución nutritiva sin NaCl todavía las plantas mostraban afectaciones en el crecimiento, las cuales no pudo revertir la EBL. Esta disminución en la velocidad de crecimiento de las plantas tratadas, durante este período, pudiera deberse en parte a que las afectaciones en la etapa inicial del crecimiento son muy difíciles de recuperar, ya que en este período ocurren en las plantas mecanismos adaptativos de defensa en el intercambio de sustancias, que conllevan al paro temporal del crecimiento por la falta de materiales de constitución o de reserva, que al normalizarse, se restablece el funcionamiento del organismo (incluidos los procesos de crecimiento), pero a un nivel menos intenso (16).
Algunos autores han señalado el hecho de que una vez concluido el período de estrés y sometidas las plantas durante siete días al tratamiento control, estas no lograron recuperarse, lo que demuestra el carácter irreversible de los daños ocasionados [20].
No obstante, en la segunda repetición del experimento (Figura 1B), el tratamiento con NaCl no afectó significativamente la longitud de la parte aérea de las plántulas hasta el final de la etapa de recuperación, donde se observa un efecto de protección de la EBL a las concentraciones de 1 y 2 µmolesL-1.
La causa de esta diferencia en la respuesta de las plantas ante el tratamiento con NaCl en ambas repeticiones, es desconocida por lo que deberemos seguir investigando para dilucidarla. La diferencia observada en la etapa de recuperación pudiera estar relacionada con las condiciones meteorológicas imperantes durante la ejecución de ambas repeticiones. Como se puede observar en la Tabla I, en la segunda repetición, tanto la temperatura máxima, como la mínima y la media durante la etapa de recuperación de las plántulas fue aproximadamente 5 grados más baja que en la primera repetición y es bien conocido, el efecto que la temperatura ejerce en el crecimiento del arroz, pues la temperatura óptima para la fase vegetativa de este cultivo es de 25-30ºC y temperaturas menores ocasionan un retraso en el crecimiento [21]. Es de destacar, que en la etapa de 4-5 hojas, que es en la que se realiza el experimento, el arroz es muy sensible a las variaciones de temperatura, ya sea disminución o aumento de su temperatura óptima [22].
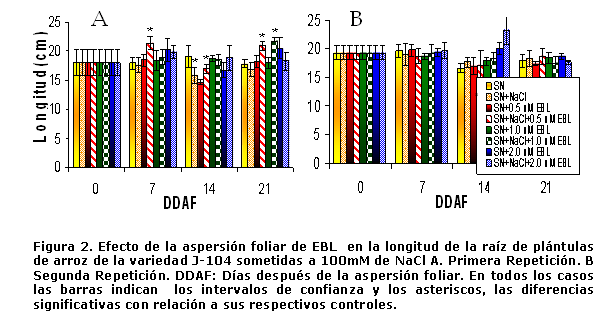
En cuanto a la longitud de la raíz en la primera repetición se observa un incremento significativo con la concentración de 0,5 µmolesL-1 de EBL tanto en el período de estrés como en la recuperación. Al final de este período se observa que la concentración de 1 µmolL-1 también logra aumentar la longitud de la raíz. (Figura 2A).
Sin embargo, en la segunda repetición del experimento (Figura 2B) no se observa este efecto, tan sólo se evidencia en la etapa de recuperación, una disminución significativa de la longitud radical en las plántulas tratadas con 2 µmolesL-1 de EBL y sometidas a 100mM de NaCl. Estas diferencias entre las dos repeticiones del experimento también pueden obedecer a las diferencias en las temperaturas registradas entre ambas repeticiones.
En el caso del arroz existe poca información sobre el efecto de los brasinoesteroides en la reducción de los efectos negativos del estrés salino.
Algunos autores han demostrado que el tratamiento por siete días con 24-EBL (1µM) fue capaz de incrementar significativamente las velocidades de crecimiento de las raíces y la parte aérea de las plántulas de arroz variedad INCA LP-7 en medio salino (100 mM NaCl), mientras que en la variedad J-104 el tratamiento con EBL (0.1µM) fue el mejor aunque sólo aumentó la masa fresca de las raíces [13].
Anuradha y Rao informaron una recuperación en el crecimiento y un incremento de los niveles de proteínas solubles y RNA en semillas de arroz var IR-64 tratadas durante 24h con NaCl (150mM) y EBL (3µM) [11]. Estos mismos autores en un trabajo posterior demostraron que la imbibición de las semillas con brasinólida (BL), EBL y homobrasinólida (HBL) (3µM) restauró considerablemente el crecimiento, los niveles de clorofila y la actividad de la nitrato reductasa ante estrés salino (150mM) [10]. Sin embargo, no utilizaron los controles respectivos sin NaCl y con BR, por lo que el efecto observado pudiera ser producto de la capacidad de estas hormonas para inducir el crecimiento y no de una verdadera protección ante el estrés salino.
Por otra parte, en Turquía, otros autores evaluaron la germinación de las semillas y el crecimiento inicial de plántulas de arroz variedad IR-28 ante estrés salino (120mM) con 24-EBL (3µM). Los resultados obtenidos fueron que los tratamientos con BR aumentaron el crecimiento de las plántulas pero no la germinación.
Como se puede observar estos reportes se refieren al tratamiento de las semillas de arroz, por lo que la aspersión foliar con brasinoesteroides está muy poco documentada.
Sin embargo, en otro cereal como es el trigo, se evaluaron los efectos de la aspersión foliar con EBL en plantas de la variedad S-24 (tolerante a la sal) y MH-97 (moderadamente sensible a la sal), dos días después de comenzado el tratamiento de estrés (suplemento de 150 mM de NaCl a la solución nutritiva Hoagland, 41 días después la siembra). Se encontró que, 45 días después de la aplicación, la EBL solamente mejoró el crecimiento en la variedad tolerante, aunque la inhibición de la actividad fotosintética provocada por la sal fue contrarrestada significativamente por este compuesto en ambos cultivares. La EBL estimuló la eficiencia del fotosistema II en ambos cultivares medido como la relación Fv/Fm; sin embargo, esta hormona no tuvo influencia en el rendimiento del grano [23].
En otro estudio, con estas dos mismas variedades, se demostró que el suministro continuo de EBL por las raíces, a través de la solución nutritiva suplementada o no con 120 mM de NaCl, mejoró no solo el crecimiento de las plantas de ambos cultivares en condiciones salinas, sino que además estimuló el rendimiento total del grano y la masa de 100 granos, siendo la concentración de 0.052 µM la más efectiva [24]. Los autores sugirieron que el incremento en el tamaño del grano inducido por la EBL pudo deberse al incremento en la translocación de más fotoasimilados hacia el grano, mientras que el mejoramiento en el crecimiento de ambos cultivares pudo estar asociado al incremento observado en la capacidad fotosintética, debido a la supresión de las limitaciones estomáticas inducidas por dicho estrés. Se conoce que, en cereales, los niveles de lectinas se incrementan en condiciones de estrés.
Por otra parte, recientemente, se demostró que el tratamiento de semillas con 3 µM de EBL disminuyó significativamente el índice mitótico de plantas de cebada crecidas en medio que contienen 300, 350 ó 400 mM de NaCl; sin embargo, esta hormona fue capaz de mejorar las anormalidades cromosómicas que se presentan en los ápices de las raíces en estas condiciones [25].
Otros autores encontraron que el tratamiento a las semillas de garbanzo con HBL, antes o después de la inmersión en NaCl, era capaz de revertir los efectos dañinos que el tratamiento salino ejerce en el crecimiento y rendimiento de estas plantas [26].
En otra especie, Brassica napus, se ha informado que la presencia de 2 µM en el medio estimuló significativamente la germinación y el crecimiento inicial de las plántulas ante condiciones salinas impuestas por 200, 250 y 300 mM de NaCl [27].
Se ha constatado que, en plántulas de mostaza expuestas a estrés por NaCl, la aspersión foliar con 1 µM de 24-EBL, 15 días después de la siembra, incrementó significativamente el crecimiento, el nivel de pigmentos y la actividad fotosintética; además de disminuir el eflujo de electrolitos y la peroxidación lipídica, lo que favorece el índice de estabilidad de las membranas [28].
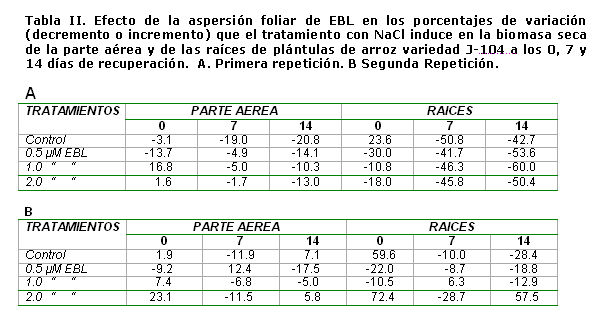
Como se puede observar en la tabla II, el tratamiento con NaCl provocó, al final del período de estrés, un incremento en la biomasa seca de las raíces que se hizo más evidente en la segunda repetición y muy poca variación en la biomasa de la parte aérea. Se debe destacar, como en este momento, sobre todo en la segunda repetición la aspersión foliar con 2 µmolesL-1 de EBL estimuló la formación de biomasa seca de las plántulas en condiciones salinas. Sin embargo, en las evaluaciones realizadas a los 7 y 14 días de la recuperación, se puede observar, como el tratamiento con NaCl indujo, de forma general, en ambas repeticiones, una reducción en la acumulación de biomasa seca tanto de la parte aérea como de las raíces de las plántulas no asperjadas con EBL. Se destaca, en la segunda repetición, el tratamiento de 2 µmolesL-1 de EBL, el cual provocó una estimulación significativa en la acumulación de biomasa seca de las raíces a los 14 días de la recuperación.
Nuestros resultados muestran que la biomasa de la raíz aumenta con el tratamiento salino, este resultado se repite en numerosos experimentos y hemos encontrado varios autores que observan un comportamiento similar [29; 30]. A pesar de que las raíces se encuentran en contacto directo con la sal, su crecimiento se ve menos afectado que el de la parte aérea, aumentando la razón raíz: vástago [30]. Esto pudiera ser una adaptación de la planta para aumentar el área de absorción de agua y disminuir el área de transpiración.
En general, los niveles de brasinoesteroides, en raíces son más bajos que en los meristemos y, al parecer, los requerimientos en este órgano también son menores [31]. Los primeros reportes mostraron que la 24-EBL tenía efectos inhibitorios en la formación de las raíces en maíz, tomate, Arabidopsis y plántulas de soya [32]. Sin embargo, otros autores han informado incrementos en la longitud de la raíz en plantas de berro (Lepidium sativum) crecidas en la oscuridad y raíces primarias de maíz [33]. Mussig y colaboradores en el 2003 demostraron que esta respuesta es dependiente de la concentración. Bajas concentraciones de BRs externos (0.1nM de EBL y 10 nM de Epicastasterona (EpiCS)) estimularon el crecimiento de la raíz de plantas de genotipos silvestres y restablecieron los valores normales en mutantes deficientes de brasinoesteroides. Las altas concentraciones de estos compuestos resultaron inhibitorias (10nM de EBL y 500 nM de EpiCS), probablemente, por estimulación de la biosíntesis de etileno, el cual es bien conocido que inhibe la elongación de las raíces [34].
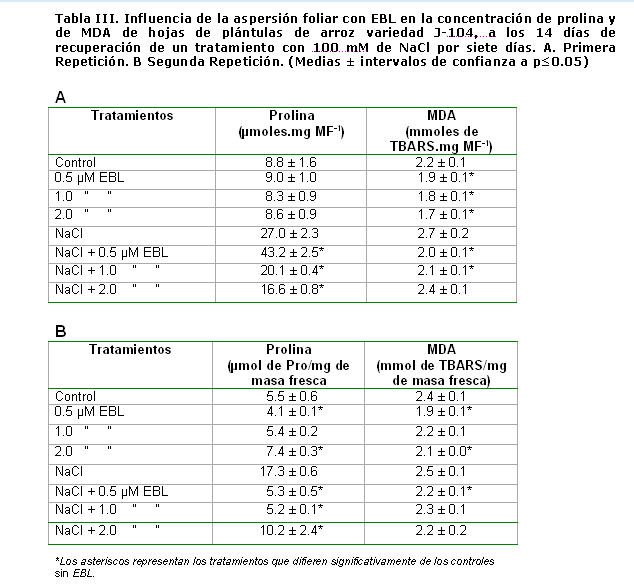
En la tabla III se muestran los niveles de malondialdehído (MDA) y prolina.
El MDA se produce cuando los ácidos grasos poliinsaturados se oxidan por efecto de los radicales libres del oxígeno Esta peroxidación lipídica da una medida indirecta del daño celular inducido por el estrés oxidativo que provoca la salinidad. Como se puede observar, en la primera repetición del experimento (Tabla III A), los niveles de MDA aumentaron levemente ante el estrés salino. Sin embargo, la EBL disminuyó significativamente estos niveles en los tratamientos sin NaCl, y en los tratamientos con estrés las concentraciones de 0,5 y 1 µmolL-1 también redujeron los niveles de peroxidación lipídica. En la segunda repetición, no hubo un aumento de los niveles de MDA ante el estrés salino. No obstante, la EBL (0,5 y 2 µmolesL-1) disminuyó los niveles de peroxidación lipídica en los tratamientos sin estrés, mientras que la concentración de 0,5 µmolesL-1 fue la única que disminuyó significativamente los niveles de MDA en las plantas tratadas con NaCl.
Este incremento de los niveles de MDA ante estrés salino ha sido informado por otros autores pero en raíces de arroz de variedades susceptibles y ha sido correlacionado con decremento en las actividades de enzimas antioxidantes como catalasa y glutationa reductasa y una actividad basal de superóxido dismutasa y ascorbato peroxidasa. Sin embargo en variedades tolerantes ha sucedido todo lo contrario [19; 35].
Además, la EBL (3µM) logró disminuir la peroxidación lipídica y la acumulación de prolina causada por el estrés salino (120mM) de plántulas de arroz variedad IR-28. Entre las enzimas antioxidantes sólo se incrementó la ascorbato peroxidasa ante el tratamiento con esta hormona [12].
En maíz, el tratamiento de semillas con otro BR, la homobrasinólida, incrementó la actividad de las enzimas antioxidantes, disminuyó la peroxidación lipídica e incrementó la concentración de proteínas solubles de plantas crecidas en condiciones salinas [36].
En cuanto a la prolina, se puede observar el incremento que experimentó este indicador a los 14 días de la recuperación en las plántulas tratadas con NaCl (Tabla III). Las plantas superiores pueden acumular L-prolina (Pro) en respuesta a diversos estrés medioambientales como sequía y salinidad, bajas temperaturas o radiación ultravioleta [37]. La acumulación de prolina estabiliza las membranas y los componentes subcelulares, incluyendo el complejo II de la cadena de transporte electrónico mitocondrial. Existen estudios que proponen a la prolina como aceptor final de los radicales libres y estabilizador del potencial redox por reaprovisionamiento de la provisión de NADP+ [38]. Además, la prolina induce la expresión de genes de respuesta a estrés salino que tienen elementos de respuesta a prolina (PRE, ACTCAT) en sus promotores [39].
Aunque la acumulación de prolina ha sido observada en arroz expuesto a estrés salino, su rol exacto es muy controversial, pues es considerada más un síntoma de daño que un indicador de tolerancia al estrés. La acumulación de prolina ha sido correlacionada con el grado de deterioro inducido por el estrés y hay autores que plantean que el aumento de este aminoácido podría deberse a una necesidad de la planta de eliminar el exceso de amonio [40].
En la primera repetición, la aspersión foliar con 0,5 µmolesL-1 de EBL aumentó aún más los niveles de prolina en las plántulas sometidas a estrés salino, mientras que las concentraciones de 1 y 2 µmolesL-1 redujeron estos niveles. En el segundo experimento, las tres concentraciones estudiadas redujeron los niveles de prolina en las plántulas tratadas con NaCl.
En plantas, la prolina es sintetizada del glutamato o la ornitina, y la primera es considerada la vía predominante en el estrés salino, ya que en estas condiciones disminuyen las actividades de la alanina y aspartato aminotransferasas acumulándose glutamato [41]. En esta vía, la biosíntesis de la prolina comienza con la fosforilación y reducción del glutamato a glutamil-5-semialdehído (G5SA) por la enzima bifuncional ?1-pirrolina-5-carboxilato sintasa (P5CS). La actividad ?-glutamil quinasa de P5CS representa el paso limitante de la velocidad de reacción de esta vía y es sensible a retroinhibición por Pro [42]. La expresión de P5CS fue fuertemente inducida por el estrés salino. Esto sugiere que P5CS en la biosíntesis de la prolina juega un papel protector bajo las condiciones de estrés, quizás controlando el potencial redox y estimulando la vía de las pentosas fosfato para la regeneración del NADP+. Algunos autores sugirieron que la acumulación de prolina inducida por el estrés salino puede ser causada por la expresión de P5CS [43].
Por otro lado, en Arabidopsis thaliana, la sequía y el estrés salino activan diferencialmente la expresión de dos genes de P5CS: AtP5CS1 (At2g39800) y AtP5CS2 (At3g55610). AtP5CS1 es responsable de la acumulación de prolina durante el estrés salino y la sequía. La expresión de AtP5CS1 es activada por una vía de traducción de señales dependiente de ABA y modulada por la luz y los brasinoesteroides [44; 45]. Esto pudiera sugerir un posible mecanismo por el cual los BR disminuyen los efectos adversos provocados por la salinidad.
A modo de conclusión, se puede plantear que la respuesta del crecimiento de las plántulas de arroz variedad J-104 al tratamiento con 100 mmolesL-1 de NaCl por siete días y a la aplicación exógena de 24-epibrasinólida estuvo muy relacionada con las temperaturas en las cuales estaban creciendo las mismas. Esto último es de gran interés científico-técnico, pues la mayoría de los resultados que se han informado en cuanto a la inducción de tolerancia al estrés salino por la 24-epibrasinólida en el cultivo del arroz se han obtenido mediante el tratamiento a la semilla (22, 23, 24) y se ha evaluado el crecimiento inicial de las plántulas en condiciones controladas; sin embargo, no hay información anterior de aplicaciones mediante aspersión foliar en esta etapa de crecimiento del cultivo en condiciones abiertas; por lo que se hace necesario continuar profundizando en este sentido, pues al parecer la respuesta a determinadas dosis de la hormona dependerá en gran medida de las condiciones de crecimiento de las plantas.
REFERENCIAS
1 IPCC. Climate Change : Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. En: 2007: University Press, Cambridge).
2 García, J.A., Gallego, M.C., Serrano, A., et al. Trends in block-seasonal extreme rainfall over the Iberian Peninsula in the second half of the twentieth century. J. Climate. 2007, Vol.20, p.113-130.
3 Ritzema, H.P., Satyanarayana, T.V., Raman, S., et al. Subsurface drainage to combat waterlogging and salinity in irrigated lands in India: Lessons learned in farmers’ fields. Agric. Water Manage. 2008, Vol. 95, p.179-189.
4 FAO. FAO Land and Plant Nutrition Management Service. . 2008. Disponible en: <http://www.fao.org/ag/agl/agll/spush>
5 Lau, A., Garea, E. and Ruiz, M.E. Estimación de la salinidad de los suelos utilizando una imagen espectrozonal y el sistema de información geográfica TELEMAP. Revista Ciencias Técnicas Agropecuarias. 2005, Vol.14, no.1, p.47-54.
6 Sahi, C., Singh, A., Kumar, K., et al. Salt stress response in rice: genetics, molecular biology, and comparative genomics. Functional and Integrative Genomics. 2006, Vol. 6, p.263-284.
7 Bajguz, A. and Hayat, S. Effects of brassinosteroids on the plant responses to environmental stresses. Plant Physiology and Biochemistry. 2009, Vol.47, no.3, p.1-8.
8 Singh, I. and Shono, M. Physiological and molecular effects of 24-epibrassinolide, a brassinosteroid on thermotolerance of tomato. . Plant Growth Regulation. 2005, Vol.47, p.111-119.
9 Kagale, S., Divi, U.K., Krochko, J.E., Keller, W.A., et al. Brassinosteroid confers tolerance in Arabidopsis thaliana and Brassica napus to a range of abiotic stresses. Planta. 2007, Vol. 225, p.353-364.
10 Anuradha, S. and Rao, S. Application of brassinosteroids to rice seeds (Oryza sativa L.) reduced the impact of salt stress on growth, prevented photosynthetic pigment loss and increase nitrate reductase activity. Plant Growth Regul. 2003, Vol.40, p.29-32
11 Anuradha, S. and Rao, S. Effect of brassinosteroids on salinity stress induced inhibition of seed germination and seedling growth of rice (Oryza sativa L.). Plant Growth Regul. 2001, Vol.33, p.151-153.
12 Ozdemir, F., Bor, M., Demiral, T., et al. Effects of 24-epibrassinolide on seed germination, seedling growth, lipid peroxidation, proline content and antioxidative system of rice (Oryza sativa L.) under salinity stress. Plant Growth Regul 2004, Vol.42, p.203-211
13 Núñez, M., Mazorra,L.M., Martínez,L., González, M. C., Robaina,C. Influencia de la 24-epibrasinólida y un análogo espirostánico de brasinoesteroides en el crecimiento de plántulas de dos variedades de arroz (Oryza sativa l.) en medio salino. Cultivos Tropicales. 2006, Vol.27, no.1, p.75-82.
14 Alfonso, R.l. Determinación de parámetros genético-fisiológicos indicadores del estrés hídrico para su empleo en el mejoramiento genético del arroz (Oryza sativa L.) y la estabilidad varietal [Tesis de grado]. IIA, 1998.
15 Instituto de Suelos. Nueva clasificación de los suelos de Cuba. La Habana : Agrinfor. Cuba. MINAGRI. 1999, p.64.
16 Bates, L., Waldren, RP,Teare, ID Rapid determination of free proline for water stress studies. Plant Soil. 1973, Vol.39, p.205-207.
17 Hodges, M.D., DeLong, J.M., Forney, C.F., et al. Improving the thiobarbituric acid-reactive-substances assay for lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta. 1999, Vol.207, p.604-611.
18 Duan, D.Y., Li, W.Q., Liu, X.J., et al. Seed germination and seedling growth of Suaeda salsa under salt stress. Ann. Bot. Fennici. 2007, Vol.44, no.161-169.
19 Demiral, T. and Turkan, I. Comparative lipid peroxidation, antioxidant defense systems and proline content in roots of two rice cultivars differing in salt tolerance. Env Exp Bot 2005, Vol. 53, p.247-257.
20 Morales, D., Bolarín, M.C. and Cayuela, E. Respuesta de plantas de arroz (Oryza sativa L.) a la aplicación de diferentes niveles de NaCl. I. Crecimiento y relaciones hídricas. Cultivos Tropicales. 2006, Vol.27, no.4, p.27-32.
21 Yoshida, S. Fundamentals of rice crop science En: (1981: Los Banos, Philippines.). International Rice Research Institute
22 Sánchez, S. and Socorro, M. 2008. Tecnología del cultivo del arroz en pequeña escala. . Oliva., E.M. (Ed.). Biblioteca ACTAF. [Consultado: Disponible en:
23 Shahbaz, M., Ashraf, M. and Athar, H.R. Does exogenous application of 24-epibrassinolide ameliorate salt induced growth inhibition in wheat (Triticum aestivum L.)? Plant Growth Regul 2008, Vol.55, p.51-64.
24 Ali, Q., Atar, H.-R. and Ashraf, M. Modulation of growth, photosynthetic capacity and water relations in salt stressed wheat plants by exogenously applied 24-epibrassinolide. . Plant Growth Regul. 2008, Vol.56, p.107-116.
25 Tabur, S. and Demir, K. Cytogenetic response of 24-epibrassinolide on the root meristem cells of barley seeds under salinity. Plant Growth Regul. 2009, Vol.58, p.119-123.
26 Ali, B., Hayat, S. and Ahmad, A. 28-Homobrassinolide ameliorates the saline stress in chickpea (Cicer arietinum L.). Environ. Exp. Bot. 2007, Vol.59, p.217-223.
27 Kagale, S., Divi, U.K., Krochko, J.E., et al. Brassinosteroid confers tolerance in Arabidopsis thaliana and Brassica napus to a range of abiotic stresses. Planta 2007, Vol. 225, p.353-364.
28 Ali, B., Hayat, S., Fariduddin, Q., Ahmad, A. 24-Epibrassinolide protects against the stress generated by salinity and nickel in Brassica juncea. Chemosphere 2008, Vol.72 p.1387-1392.
29 Qayyum, B., Shahbaz, M. and Akram, N.A. Interactive effect of foliar application of 24-Epibrassinolide and root zone salinity on morpho-physiological attributes of wheat (Triticum aestivum L.). International Journal of Agriculture & Biology. 2007, Vol.9, no.4, p.584-589.
30 Moud, A.M. and Maghsoudi, K. Salt stress effects on respiration and growth of germinated seeds of different wheat (Triticum aestivum L.) cultivars. World Journal of Agricultural Sciences. 2008, Vol.4, no.3, p.351-358.
31 Shimada, Y., Goda, H., Nakamura, A., et al. Organ-specific expression of brassinosteroid-biosynthetic genes and distribution of endogenous brassinosteroids in Arabidopsis. Plant Physiol 2003, Vol.131, p.287-297.
32 Sasse, J.M. Physiological actions of brassinosteroids: an update. J. Plant Growth Regul. . 2003, Vol.22, p.276-288.
33 Kim, S.K., Chang, S.C., Lee, E.J., et al. Involvement of brassinosteroids in the gravitropic response of primary root of maize Plant Physiol. 2000, Vol.123, p.997-1004.
34 Mussig, C., Shin, G.H. and Altmann, T. Brassinosteroids promote root growth in Arabidopsis. Plant Physiol. 2003, Vol.133, p.1261-1271.
35 Khan, M.H. and Panda, S.K. Alterations in root lipid peroxidation and antioxidative responses in two rice cultivars under NaCl-salinity stress. Acta Physiol Plant 2008, Vol.30, p.81-89.
36 Arora, N., Bhardwaj, R., Sharma, P., et al. 28-homobrassinolide alleviates oxidative stress in salt treated maize (Zea mays L.) plants. Brazilian J. Plant Physiol. 2008, Vol.20, p.153-157.
37 Munns, R. and Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, Vol.59, no.651-681.
38 Hamilton, E., Heckathorn, SA Mitochondrial adaptations to NaCl. Complex I is protected by anti-oxidants and small heat shock proteins, whereas complex II is protected by proline and betaine. Plant Physiol. 2001, Vol.126, p.1266-1274.
39 Satoh, R., Nakashima, K., Seki, M., et al. ACTCAT, a novel cis-acting element for proline and hypoosmolarity-responsive expression of the ProDH gene encoding proline dehydrogenase in Arabidopsis. Plant Physiol. 2002, Vol.130, p.709-719.
40 Amini, F. and Ehsanpour, A.A. Soluble proteins, proline, carbohydrates and Na+/K+ changes in two tomato (Lycopersicon esculentum Mill.) cultivars under in vitro salt stress. Am. J. Biochem & Biotech. . 2005, Vol.1 no.4, p.212-216.
41 Pandey, R., Agarwal, R.M., Jeevaratnam, K., et al. Osmotic stress-induced alterations in rice (Oryza sativa L.) and recovery on stress release. Plant Growth Regulation 2004, Vol.42, p.79-87.
42 Dea-Wook, K., Junko, S., Ganesh, K.A., et al. Gene transcription in the leaves of rice undergoing salt-induced morphological changes (Oryza sativa L.). Mol. Cells. 2007, Vol.24, no.1, p.45-59.
43 Kant , S., Kant , P., Raveh, E., et al. Evidence that differential gene expression between the halophyte Thellungiella halophila, and Arabidopsis thaliana is responsible for higher levels of the compatible osmolyte proline and tight control of Na+ uptake in T. halophila. . Plant Cell Environ. 2006, Vol.29, p.1220-1234.
44 Savouré, A., Hua, XJ, Bertauche, N, Van Montagu, M, Verbruggen, N. Abscisic acid-independent and abscisic acid-dependent regulation of proline biosynthesis following cold and osmotic stresses. Mol Gen Genet. 1997, Vol.254, no.104-109.
45 Strizhov, N., Abraham, E., Okresz, L., Zilberstein, A., Schell, J., Koncz, C., Szabados, L. Differential expression of two P5CS genes controlling proline accumulation during salt-stress requires ABA and is regulated by ABA1,ABI1 and AXR2 in Arabidopsis. Plant J. 1997, Vol.12, no.3, p.557-569.
Recibido 2/07/2010, aceptado 12/07/2011
Yanelis Reyes Guerrero, Departamento de Fisiología y Bioquímica Vegetal, Instituto Nacional de Ciencias Agrícolas, gaveta postal 1, San José de las Lajas,Mayabeque, CP 32700. Email: yanelisrg@inca.edu.cu


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}