










My SciELO
 Custom services
Custom servicesServices on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkCultivos Tropicales
Print version ISSN 0258-5936
cultrop vol.33 no.3 La Habana July-Sept. 2012
Efecto del preacondicionamiento a la salinidad en las relaciones hídricas, el intercambio gaseoso y la conductividad hidráulica de las raíces en plantas de tomate (Solanum lycopersicum L. cv. Amalia)
Salinity preconditioning effects on water relations, gas exchange and root hydraulic conductivity in tomato plants (Solanum lycopersicum L. cv. Amalia)
Dr.C. Donaldo Morales Guevara I y Dr.C. José M. Dell’Amico Rodríguez, I Dr.C. Pedro Rodríguez Hernández, II Dra. María de J. Sánchez-Blanco, III y Dr. Arturo Torrecillas Melendreras III
IInvestigadores Titulares del departamento de Fisiología y Bioquímica, Instituto Nacional de Ciencias Agrícolas, Gaveta Postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32 700.
IIInvestigador Titular del departamento de Fisiología y Bioquímica, Instituto Nacional de Ciencias Agrícolas, Gaveta Postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32 700.
IIICentro de Edafología y Biología Aplicada del Segura (CEBAS), Murcia, España.
RESUMEN
El presente trabajo se realizó con el objetivo de estudiar el comportamiento del crecimiento, la tasa fotosintética neta, la conductancia estomática, la conductividad hidráulica de las raíces y los potenciales hídrico foliar, osmótico y de turgencia en plántulas de tomate sometidas a incrementos progresivos de NaCl en el medio, para ello, plántulas de tomate (Solanum lycopersicum L. Cv. Amalia) se sometieron a siete tratamientos salinos diferentes: control (C) en el que se mantuvieron en solución nutritiva sin NaCl; preacondicionamiento (PC) en el que recibieron cada cuatro días un aumento progresivo de 25 mM de NaCl y los otros cinco tratamientos en los que se sometieron a 25, 50, 75, 100 y 125 mM de NaCl durante todo el periodo experimental. La acumulación de masa seca en el sistema radical y en el tallo se afectó en todos los tratamientos excepto en el que las plantas se preacondicionaron, mientras que la masa seca de las hojas y el área foliar se afectaron en todos los tratamientos respecto al control. El estrés salino disminuyó la tasa fotosintética neta, la conductancia estomática y la conductividad hidráulica de las raíces en las plántulas no preacondicionadas, mientras que las plantas PC mostraron un buen ajuste osmótico que les permitió mantener la turgencia de la hoja disminuyendo el potencial hídrico foliar y el osmótico a plena turgencia mientras mantuvo una adecuada actividad biológica.
Palabras clave: fotosíntesis, conductancia estomática, potencial hídrico foliar, conductividad hidráulica, crecimiento.
ABSTRACT
The present work was conducted with the objective of studying the behavior of growth, the net photosynthetic rate, stomatal conductance, hydraulic conductivity of roots and leaf water, osmotic and turgor potencials in seedlins of tomato plants submitted to progressive increase to NaCl in the medium, for this seedlins of tomato plants (Solanum lycopersicum L. Cv. Amalia) were submitted to seven different treatments: control (C) plants were maintained in nutrient solution without NaCl; adapted plants (PC) were submitted to progressive increase of 25 mM of NaCl each four days in nutritious solution, in the others treatments were submitted to 25, 50, 75, 100 and 125 mM of NaCl during all experimental period. The inhibition of dry mass accumulation in root system and stem was observed in non adapted plants, while the leaf dry mass and leaf area were affected in all treatments in relation to control plants. Saline stress decreased net photosynthetic rate, stomatal conductance and root hydraulic conductivity in non adapted plants, however, PC plants showed good osmotic adjustment, which enabled them to maintain leaf pressure potential, decreased leaf water potential and osmotic potential at full turgor while the biologic activity was appropriate.
Key words: photosynthesis rate, stomatal conductance, leaf water potential, hydraulic conductivity, growth.
INTRODUCCIÓN
La salinidad es un estrés abiótico complejo que simultáneamente presenta componentes osmóticos e iónicos (1). Estos autores consideran que una concentración alta de sales en el medio radical afecta negativamente el desarrollo de la planta, debido fundamentalmente a los efectos hiperosmóticos y hiperiónicos del estrés.
Se ha señalado que la pérdida de homeostasis hídrica e iónica, tanto a escala celular como a nivel de la planta causa serios daños moleculares que detienen el crecimiento de la planta (2). Por una parte, la disminución del potencial hídrico del medio restringe la absorción del agua por las raíces y la absorción de los iones salinos específicos aumentan en los tejidos de la planta en concentraciones que terminan siendo tóxicas, o al mismo tiempo pueden inducir desequilibrios nutricionales debidos a modificaciones en la absorción y distribución de los nutrientes esenciales.
El estudio de la tolerancia a la salinidad es en extremo complicado y el intento de resolver, o por lo menos mitigar los efectos dañinos de las sales, incluye la valoración de cómo estas afectan los procesos fisiológicos, bioquímicos y moleculares del metabolismo de las plantas, así como de los diferentes mecanismos que las plantas utilizan para enfrentar el estrés1.
El estrés salino altera las relaciones hídricas de la planta a través de los estrés osmótico e hídrico. En respuesta a estos estrés, las plantas desarrollan el mecanismo de ajuste osmótico el que aun en condiciones de déficit hídrico mantiene una adecuada turgencia que les permite el crecimiento, el transporte, la acumulación y la compartimentación de los iones orgánicos e inorgánicos fundamentalmente en las plantas superiores (3).
Estudios realizados por algunos autores han estimado que el 6 % de los suelos del mundo y el 30 % de las áreas irrigadas presentan problemas de salinidad (4). La expansión de la agricultura hacia regiones áridas y semi-áridas utilizando la fertilización y el regadío intensivos, provocan un incremento de la salinización secundaria debido a la acumulación progresiva de las sales en el suelo como consecuencia de la disolución de la sal en el agua aplicada, asociada con una tasa elevada de evapotranspiración (5).
Según diversos autores, la fotosíntesis y el crecimiento de las plantas son los procesos que primero se afectan por la salinidad (6). Al respecto otros señalaron que el estrés hídrico y el salino pueden afectar la fotosíntesis directa o indirectamente por el decrecimiento de la disponibilidad de CO2 causada por limitaciones en la difusión de este compuesto (7).
El objetivo de este trabajo consistió en determinar la respuesta del crecimiento, la tasa neta de fotosíntesis, la conductancia estomática, la conductibilidad hidráulica de la raíz y los potenciales hídrico, osmótico y de presión en plantas de tomate (Solanum lycopersicum L.) preacondicionadas y no preacondicionadas al estrés salino.
MATERIALES Y MÉTODOS
Para el desarrollo del trabajo se utilizaron plantas de tomate (Solanum lycopersicum L. cv. Amalia) obtenidas y crecidas en recipientes de 0.5 dm3 en las que se colocó como sustrato arena sílice lavada, todos los recipientes se situaron en una cámara de crecimiento con un fotoperiodo de 13-h, una radiación fotosintéticamente activa de 380 µmol.m-2.s-1, con una temperatura media y una humedad relativa día/noche de 25/18oC y 60/70 %, respectivamente.
Desde la segunda semana posterior a la siembra todos los recipientes fueron colocados en bandejas con solución nutritiva y cuando las plantas alcanzaron su cuarto par de hojas, se seleccionaron 152 plantas de apariencia similar y se sometieron a siete tratamientos salinos: un control sin sal (C), un tratamiento en el que a las plantas se les fue incrementando el nivel de salinidad cada cuatro días a razón de 25 mM de NaCl hasta llegar a 150 mM (PC) y otros tratamientos en los que las plantas permanecieron durante 24 días a niveles de 25, 50, 75, 100 y 125 mM NaCl.
Evaluaciones: Antes de iniciar los tratamientos se les determinó la masa seca a cada parte de las plantas, así como, la superficie foliar usando un sistema de análisis de imagen (D-T Devices Ltd., Cambridge, UK).
Al finalizar el periodo experimental, se determinó el potencial hídrico foliar (Yw) con una cámara de presión (Soil Moisture Equipment Co, Santa Barbara USA). Las hojas usadas para las medidas de Yw se congelaron en nitrógeno líquido, después se descongelaron y se les determinó el potencial osmótico (Ys), utilizando un osmómetro de presión de vapor Wescor 5500 (Wescor, Logan, EE.UU.).
El potencial de presión (Yp) se estimó como la diferencia entre Yw y Ys.
Para la medida del potencial osmótico a máxima saturación (Yos) las hojas fueron hidratadas durante 24 horas colocando sus peciolos en un recipiente con agua destilada, cubiertos con papel parafilm y situados a bajas temperaturas en un refrigerador doméstico, posteriormente se congelaron en nitrógeno líquido y se siguió el mismo procedimiento aplicado para determinar el Ys.
La conductancia estomática (gs) y la tasa de fotosíntesis neta (Pn) se midieron al mediodía cuando la radiación era de 380 µmol.m-2.s-1, para ello se utilizó un medidor portátil de fotosíntesis LICOR LI-6200 (LI-COR, Lincoln, EE.UU.).
Para medir la conductividad hidráulica de las raíces, las plantas fueron cortadas al nivel del cuello de la raíz, extraídas de las macetas y lavado cuidadosamente su sistema radical. Posteriormente se colocaron en un recipiente con solución nutritiva de Hoagland e introducidas en la cámara de presión quedando hacia el exterior una parte del tallo en el que se ajustó un pequeño tramo de manguera tipo capilar. A partir de ese momento se comenzó a incrementar la presión dentro de la cámara hasta que se obtuvo un flujo constante, a partir de entonces esta se fue incrementando a una taza de 0.4 MPa.min-1 hasta alcanzar una presión final de 1 MPa. A cada planta se les extrajeron tres exudados, midiéndose el volumen extraído cada tres minutos. La conductividad hidráulica de la raíz se calculó usando la fórmula:
Lp = J/(P x L)
donde
Lp: expresada en mg.m-1.s-1.MPa-1
P: presión hidrostática aplicada (MPa)
L: largo de la raíz (m)
J: tasa de flujo de agua a través de todo el sistema radical (mg.s-1).
Se utilizó un diseño de bloques completamente aleatorizados. Cada cuatro días se evaluaron cuatro plantas en los tratamientos control y el de adaptación a la salinidad (PC) y cuatro plantas en los restantes tratamientos al finalizar el período. Los datos obtenidos fueron analizados según un modelo de clasificación simple (8) comparándose las medias a través de la prueba de rangos múltiples de Duncan.
RESULTADOS Y DISCUSIÓN
En la Tabla I se presentan las masas secas de raíz, tallo y hojas, así como la superficie foliar al finalizar el periodo en el que las plantas estuvieron sometidas a los diferentes tratamientos.
Como se puede apreciar, en la medida en que se fueron incrementando los niveles de NaCl en el medio, hubo un decrecimiento significativo de las diferentes variables evaluadas, pero destaca el hecho de que las plantas en las que el contenido salino del medio se les fue incrementando paulatinamente, mostraron un comportamiento similar al de las plantas control tanto en la masa seca de las raíces como de los tallos y aunque en las demás variables presentaron diferencias estadísticamente significativas con las plantas control, los valores alcanzados se tornaron bastante cercanos a los de las plantas que no fueron tratadas con NaCl.
Estos resultados parecen indicar que este cultivar es capaz de adaptarse a condiciones de cultivo en las que la salinidad se vaya incrementando durante su desarrollo, habilidad que le permitiría cultivarse bajo condiciones de riego con aguas contaminadas con contenidos moderados de cloruro de sodio.
Cuando se analizó la respuesta de las plantas control y las sometidas a incrementos paulatinos de la salinidad en medio (Figura 1) a través del periodo experimental, se apreció cómo el solo hecho de someterlas durante cuatro días a un tratamiento de 25 mm de NaCl, afectó la acumulación de masa seca del sistema radical y de los tallos; sin embargo, esta última variable se recuperó inmediatamente e igualó su comportamiento a los cuatro días posteriores cuando ya había recibido un incremento de 25 mM de NaCl en la solución nutritiva, mientras que el sistema radical de este tratamiento se equilibró con el de las plantas control después de permanecer durante cuatro días a 100 mM de NaCl en el medio. Es en este momento cuando se observó un efecto desfavorable en la acumulación de materia seca en las hojas, mientras que la superficie foliar no manifestó diferencias estadísticas con el control.

Este resultado revela la habilidad notable de las raíces y los tallos a la adaptación cuando las plantas son expuestas gradualmente a concentraciones crecientes de NaCl, mientras la masa foliar es afectada quizás por la reducción observada del tamaño de la hoja.
El efecto observado en el crecimiento de las plantas expuestas a la salinidad pudiera estar asociado, según lo señalado por algunos, a daños oxidativos como resultado de un desbalance entre la producción de especies reactiva de oxígeno (ROS) y la defensa por sustancias antioxidantes (9, 10).
Se han reportado efectos adversos del estrés salino en el crecimiento de las plantas al informar acerca del comportamiento de diferentes tipos de plantas en condiciones salinas (11, 12), quienes además señalaron que la salinidad ejerce su efecto perjudicial principalmente mediante desajustes del equilibrio iónico y osmótico de la célula siendo bien conocido que la señal del estrés es primeramente recibida por los receptores a nivel de membrana y después es traducida en la célula.
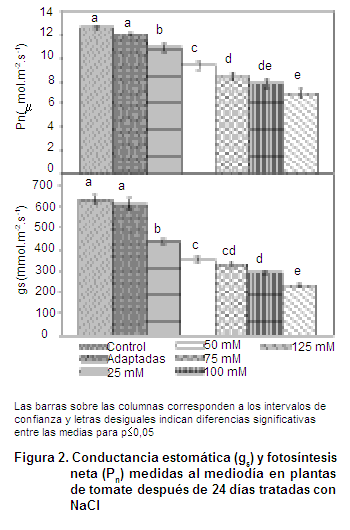
La Figura 2 muestra el comportamiento de la actividad fotosintética y la conductancia estomática en plantas de tomate después de permanecer durante 24 días sometidas a diferentes niveles de salinidad en el medio en que se desarrollaron.
En la Figura 2 se denota como estas variables manifestaron un comportamiento similar entre las plantas que no recibieron aportes de NaCl en la solución nutritiva y las que las recibieron de manera paulatina cada cuatro días. De igual forma indica que en este cultivar el intercambio gaseoso se ve afectado cuando es sometido durante 24 días a niveles constantes de salinidad, observándose la tendencia a disminuir sus valores con el incremento de la concentración salina.
Estos resultados concuerdan con los informados por otros autores quienes indicaron que esta reducción puede estar asociada a la pérdida de capacidad de las plantas para absorber el agua en condiciones salinas provocando una regulación estomática (13, 14).
De igual forma, también se ha señalado que en las plantas expuestas, tanto a sequía como a salinidad, manifiestan respuestas fisiológicas, bioquímicas y moleculares similares, tales como un cierre estomático y el decrecimiento de la tasa de intercambio gaseoso, de la acumulación de osmolitos, regulaciones de los sistemas enzimáticos antioxidantes y cambios en sus hormonas (15, 16, 17)
De acuerdo con lo planteado por otros, este comportamiento es consecuencia de limitaciones estomáticas y no estomáticas que inhiben los procesos fotoquímicos que suceden cuando se induce la salinidad provocando por tanto, un decrecimiento de la actividad fotosintética (18).
La Figura 3 refleja el comportamiento de los potenciales hídrico foliar, osmótico a plena turgencia, de turgencia y la conductividad hidráulica de las raíces después de las plantas permanecer durante 24 días expuestas a diferentes niveles de NaCl en la solución nutritiva en que se desarrollaron.
Al observar la Figura 3, se puede apreciar cómo los diferentes potenciales y la conductividad hidráulica de las raíces disminuyeron significativamente respecto al tratamiento control con el incremento de la salinidad, excepto el potencial de turgencia que no presentó diferencias significativas entre las plantas que recibieron un incremento paulatino de la salinidad y el control.
Se han observado efectos depresivos del potencial hídrico en condiciones salinas en Portulaca oleoracea L. (19) y en Capsicum annum L. (20).
Por otra parte, al observar el comportamiento del potencial osmótico a plena turgencia, se puede inferir que este cultivar es capaz de desarrollar el mecanismo de ajuste osmótico, lo que le permitió mantener una adecuada turgencia foliar y por tanto, un comportamiento muy similar a las plantas sin adiciones de NaCl en el medio, ya sea en su intercambio gaseoso como en su crecimiento.
Diversos autores han coincidido en señalar que la tolerancia de las plantas puede ser debida a la presencia del ión homeostasis, a mecanismos de ajuste osmótico o un sincronismo eficiente de la acción de varios componentes enzimáticos y no enzimáticos de los sistemas antioxidantes de defensa (21, 22, 23, 24).
Resultados que concuerdan con los informados, en relación con la conductividad hidráulica de las raíces, han sido reportados en plantas de Arbutus unedo (25).
CONCLUSIONES
Los resultados antes expuestos, indican la capacidad de adaptación de este cultivar a incrementos progresivos de la salinidad, hecho que sugiere la conveniencia de profundizar en su comportamiento en condiciones de campo dadas sus potencialidades para ser utilizada cuando las aguas disponibles para el riego poseen un determinado contenido de NaCl.
REFERENCIAS
1. Sam, O.; Ramírez, C.; Coronado, M. J.; Testillano, P. S. y Risueño, M. del C. Changes in tomato leaves induced by NaCl stress: leaf organization and cell ultrastructure. Biologia Plantarum, 2003, vol. 47, no. 3. p. 361-366.
2. Zhu, J. K. Plant salt tolerance. Trends Plant Science, 2001, vol. 6, p. 66-71.
3. Zhu, J. K. Salt and drought Stress Signal Transduction in Plants. Annu. Rev. Plant Biol., 2002, vol. 53, p. 247-273.
4. Munns, R. y Tester, M. Mechanisms of salinity tolerance. Annual Review Plant Biology, 2008, vol. 59, p. 651-681.
5. Chaves, M. M.; Flexas, J. y Pinheiro, C. Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell. Annals of Botany, 2008, vol. 125, p. 1-10.
6. Munns, R.; James, R. A. y Läuchli, A. Approaches to increasing the salt tolerance of wheat and other cereals. Journal of Experimental Botany, 2006, vol. 57, p. 1025-1043.
7. Flexas, J.; Diaz-Espejo, A.; Galmés, J.; Kaldenhoff, R.; Medrano, H. y Ribas-Carbo, M. Rapid variations of mesophyll conductance in response to changes in CO2 concentration around leaves. Plant Cell and Environment, 2007, vol. 30, p. 1284-1298.
8. Snedecor, G. Métodos estadísticos. G. Snedecor, W. Cochran. C. México : Ed. Continental, 1971, 720 p.
9. Manchanda, G. y Garg, N. Salinity and its effect on the functional biology of legumes. Acta Physiol. Plant, 2008, vol. 30, p. 595-618.
10. Vital, S. A.; Fowler, R. W.; Virgen, A.; Gossett, D. R.; Banks, S. W. y Rodriguez, J. Opposing roles for superoxide and nitric oxide in the NaCl stress-induced upregulation of antioxidant enzyme activity in cotton callus tissue. Environ. Exp. Bot., 2008, vol. 62, p. 60-68.
11. Goykovic Cortés, V. y Saavedra del Real, G. Algunos efectos de la salinidad en el cultivo del tomate y prácticas agronómicas de su manejo. IDESIA, 2007, vol. 25, no. 3, p. 47-58.
12. Mahajan, S. y Tuteja, N. Cold, salinity and drought stresses: An overview. Archives of Biochemistry and Biophysics, 2005, vol. 444, no. 2 , p. 139-158.
13. Khan, N. A.; Nazar, R. y Anjum, N. A. Growth, photosynthesis and antioxidant metabolism in mustard (Brassica juncea L.) cultivars differing in ATP-sulfurylase activity under salinity stress. Scientia Horticulturae, 2009, vol. 122, p. 455-460.
14. Tabatabaei, S. J. Effects of salinity and N on the growth, photosynthesis and N status of olive (Olea europaea L.) trees. Scientia Horticulturae, 2006, vol. 108, no. 4, p. 432-438.
15. Wang, R.; Chen, S.; Zhou, X. /et al./. Ionic homeostasis and reactive oxygen species control in leaves and xylem sap of two poplars subjected to NaCl stress. Tree Physiology, 2008, vol. 28, p. 947-957.
16. Chaves M. M.; Flexas J. y Pinheiro, C. Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell. Annals of Botany, 2009, vol. 103, p. 551-560.
17. Chen, L.; Zhang, S.; Zhao, H.; Korpelainen, H.; Li, Ch. Sex-related adaptive responses to interaction of drought and salinity in Populus yunnanensis. Plant, Cell and Environment, 2010, vol. 33, p. 1767-1778.
18. Steduto, P.; Albrizio, R.; Giorio, P. y Sorrentino, G. Gas exchange response and stomatal and non-stomatal limitations to carbon assimilation of sunflower under salinity. Environ. Exp. Bot., 2000, vol. 44, p. 243-255.
19. Yazici, I.; Turkan, I.; Sekmen, A. H. y Demiral, T. Salinity tolerance of purslane (Portulaca oleoracea L.) is achieved by enhanced antioxidative system, lower level of lipid peroxidation and proline accumulation. Environ. Exp. Bot., 2007, vol. 61, p. 49-57.
20. Silva, C.; Martínez, V. y Carvajal, M. Osmotic versus toxic effects of NaCl on pepper plants. Biol. Plant., 2008, vol. 52, p. 72-79.
21. Radic, S.; Radic-Stoikovic, M. y Pevalek-Kozlina, B. Influence of NaCl and mannitol on peroxidase activity and lipid peroxidation in Centaurea ragusina L. roots and shoots. J. Plant Physiol., 2006, vol. 163, p. 1284-1292.
22. Sumithra, K.; Jutur, P. P.; Carmel, D. y Reddy, A. R. Salinity-induced changes in two cultivars of Vigna radiata responses of antioxidative and proline metabolism. Plant Growth Regul., 2006, vol. 50, p. 11-22.
23. Wahid, A.; Perveen, M.; Gelani, S. y Basra, S. M. Pretreatment of seed with H2O2 improves salt tolerance of wheat seedlings by alleviation of oxidative damage and expression of stress proteins. J. Plant Physiol., 2007, vol. 164, p. 283-294.
24. Yang, Y.; Xu, S.; An, L. y Chen, N. NADPH oxidase-dependent hydrogen peroxide production, induced by salinity stress, may be involved in the regulation of total calcium in roots of wheat. J. Plant Physiol., 2007, vol. 164, p. 1429-1435.
25. Navarro, A.; Bañon, S.; Olmos, E. y Sánchez-Blanco, M. de J. Effects of sodium chloride on water potential components, hydraulic conductivity, gas exchange and leaf ultrastructure of Arbutus unedo plants. Plant Science, 2007, vol. 172, no. 3, p. 473-480.
Recibido: 22 de junio de 2011
Aceptado: 7 de mayo de 2012
Dr.C. Donaldo Morales Guevara, Investigadores Titulares del departamento de Fisiología y Bioquímica, Instituto Nacional de Ciencias Agrícolas, Gaveta Postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32 700. Email: dmorales@inca.edu.cu


{kind=link}
{kind=link}
{kind=link}