










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCultivos Tropicales
versión impresa ISSN 0258-5936
cultrop vol.33 no.4 La Habana oct.-dic. 2012
Resistencia al Tomato Yellow Leaf Curl Virus (TYLCV-IL [CU]) en poblaciones F2 de tomate (S. lycopersicum, L.), segregantes para los genes de resistencia Ty-1 y Ty-2
Resistance to Tomato Yellow Leaf Curl Virus (TYLCV-IL [CU]) in F2 tomato (S. lycopersicum, L.) populations segregating for Ty-1 y Ty-2 genes
Dra.C. Marta Álvarez Gil,I y Ms.C. Francisco Dueñas Hurtado,II Dr.C Alejandro Fuentes,III Dra.C. Yamila Martínez Zubiaur,IV Dr. Rafael Fernández MuñozV
IInvestigador Titular del departamento de Genética y Mejoramiento Vegetal, Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, Mayabeque.
IIInvestigador Agregado del departamento de Genética y Mejoramiento Vegetal, Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, Mayabeque
IIIInvestigador Titular, Centro de Ingeniería Genética y Biotecnología, La Habana
IVInvestigador Titular, Centro Nacional de Sanidad Agropecuaria (CENSA), Mayabeque, Cuba. CP 32 700
VCientífico Titular, Instituto de Hortofruticultura Subtropical y Mediterránea «La Mayora», IHSM-UMA-CSIC, Málaga, España.
RESUMEN
La enfermedad ‘Encrespamiento amarillo de la hoja de tomate’, causada por Tomato Yellow Leaf Curl Virus (TYLCV-IL [CU]) ha ocasionado cuantiosos daños a las cosechas de tomate en Cuba. Existen diversos genes de resistencia a esta enfermedad que han sido introgresados a tomate desde especies silvestres de Solanum secc. Lycopersicon. El objetivo de este trabajo fue evaluar la segregación de la resistencia conferida por dos de estos genes, Ty-1 y Ty-2, como base para la selección en la mejora para la resistencia a TYLCV-IL [CU]. Se obtuvieron dos poblaciones F2 de los cruces entre el cultivar susceptible ‘Amalia’ y dos cultivares resistentes,’H-24' (Ty-2) y ‘LA3473’ (Ty-1), derivándose las poblaciones TY-2F2 y TY-1F2, respectivamente. A las plantas, con 25 días tras la siembra, se les expuso a poblaciones de moscas blancas (Bemisia tabaci, biotipo B) virulíferas (aislado cubano, TYLCV-IL [CU]) durante 48 horas, antes del trasplante a campo. A los 30 días después de la inoculación, se les evaluó el índice de severidad de los síntomas de la enfermedad mediante una escala de 0-4 y el contenido de ADN viral en hojas apicales, mediante hibridación molecular con sondas específicas para TYLCV («dot blot»). Los resultados indicaron que la distribución de la frecuencia de plantas F2 según el índice de severidad de la enfermedad y la concentración de ADN viral fue diferente en las poblaciones TY-2F2 y TY-1F2. Se sugiere seleccionar aquellas plantas que tengan valor de escala 1 para la población TY-1F2 y valor 0 para la TY-2F2 para incorporar los genes de resistencia Ty-1 y Ty-2, respectivamente.
Palabras clave: Tomato YellowLeafCurlVirus, Begomovirus, TYLCV, Solanum lycopersicum, tomate.
ABSTRACT
The ‘Tomato yellow leaf curl disease’, caused by Tomato Yellow Leaf Curl Virus (TYLCV-IL [CU]) has caused extensive damage to tomato crops in Cuba. There are several resistance genes that have been introgresed to tomato from wild Solanum species. The objective of this study was to assess the segregation of two such genes, Ty-1 and Ty-2, as the basis for selection in improving resistance to TYLCV-IL [CU]. Two F2 populations, susceptible cultivar ‘Amalia’ x resistant cultivar ‘H-24’ (Ty-2), TY-2F2 and ‘Amalia’ x resistant cultivar ‘LA3473’ (Ty-1), TY-1F2, were obtained. Plants F2 and plants of susceptible controls, cvs. ‘Amalia’ and ‘STY-1’. The plants, 25 days after sowing, were inoculated with viruliferous (Cuban isolate TYLCV-IL [CU]) whiteflies (Bemisia tabaci, biotype B) for 48 hours before transplantation to the field. The disease severity index (DSI, 0-4 scale) and viral DNA (dot blot hybridization) were evaluated 30 days after the inoculation. The plant frequency distribution of disease severity and viral DNA concentration changed according to TY-2F2 and TY-1F2 plants. Therefore, we should select as resistant plants to TYLCV those with 1 score in TY-1F2 and 0 scored in TY-2F2, when the improvement program is performed by selecting phenotypic resistance TYLCV-IL [CU] conferred by Ty-1 and Ty-2 resistance genes, respectively.
Key words: Tomato Yellow Leaf Curl Virus, Begomovirus, TYLCV, Solanum lycopersicum, tomato.
INTRODUCCIÓN
La enfermedad conocida como ‘Encrespamiento amarillo de la hoja de tomate’ (TYLCD, «Tomato Yellow Leaf Curl Disease») es causada por un complejo de virus de los denominados virus del ‘Encrespamiento amarillo de la hoja de tomate’ (TYLCV, Tomato yellow leaf curl virus), que pertenecen a dos especies del género Begomovirus (1) y que son transmitidos por la mosca blanca (Bemisia tabaci Genn.) (Homoptera: Aleyrodidae) (2). Estos virus causan serios daños en todas las principales regiones productoras de vegetales del mundo; que comprenden la cuenca Mediterránea, el África tropical, el Sureste asiático, América del Norte y del Sur y la región del Caribe (3, 4).
En Cuba, el TYLCV se ha distribuido ampliamente desde 1980, causando grandes pérdidas en la producción de tomate, que han alcanzado hasta el 100 % al cultivarse únicamente variedades susceptibles (5). Asimismo, se determinó que el aislado cubano, TYLCV-IL [CU], es homólogo al aislado de Israel en un 98 %1 (6), disponiéndose en principio solo de un cultivar resistente (‘Vyta’, IIHLD, Instituto de Investigaciones Hortícolas Liliana Dimitrova) adaptado a las condiciones cubanas, lo cual es insuficiente para la producción de tomate en el país.
A nivel mundial se conoce que la mejor estrategia para evitar los daños y la diseminación de esa virosis es el empleo de cultivares resistentes (7, 8), por lo cual se han encaminado esfuerzos hacia la introgresión hacia el tomate cultivado (Solanum lycopersicum L.) de genes de resistencia desde especies silvestres emparentadas del género Solanum secc. Lycopersicon.
Entre las especies empleadas como fuente de resistencia a virus causantes de TYLCD se encuentran: S. peruvianum L., S. chilense (Dunal) Reiche, S. habrochaites S. Knapp & D.M. Spooner, S. cheesmaniae (L. Riley) Fosberg y S. pimpinellifolium L. (9, 10), diseñándose estrategias para producir cultivares e híbridos resistentes a partir de las líneas primarias de tomate a las que se les ha introducido la resistencia. De este genofondo, las resistencias mas explotadas han sido aquellas provenientes de S. chilense, S. peruvianum y S. habrochaites (11), produciéndose numerosos cultivares e híbridos resistentes capaces de la supresión de la acumulación del virus y/o su diseminación, bajo condiciones ambientales que favorezcan la infección viral (8).
El mejor aprovechamiento de estas ‘resistencias’ lo determina su base genética, la cual puede variar, desde un gen simple dominante, hasta un patrón cuantitativo recesivo (12). Hasta el presente, se han identificado muchos loci de resistencia, aunque solo han sido mapeados cinco loci mayores que confieren resistencia a TYLCV (Ty-1 - Ty-5): Ty-1 y Ty-3 son dos loci semi-dominantes introgresados desde S. chilense y localizados en el brazo corto del cromosoma 6 (13, 14); el locus Ty-4, proveniente también de S. chilense, ubicado en el brazo largo del cromosoma 3 (15); el locus Ty-2, originado en S. habrochaites, con un efecto parcialmente dominante, mapeado en el brazo largo del cromosoma 11 (16) y, recientemente, el locus Ty-5, proveniente de S. peruvianum, que fue localizado en el cromosoma 4 (17, 18).
Para que los mejoradores decidan qué gen o genes incorporar a sus cultivares susceptibles a mejorar, es muy necesaria la información sobre el comportamiento de esas ‘resistencias’ frente a los aislados locales de TYLCV. También, determinar la forma de seleccionar los fenotipos resistentes en las plantas segregantes a partir de los cruzamientos entre los cultivares susceptibles y resistentes.
Como antecedentes, se ha demostrado (5, 19) que los genes Ty-1 y Ty-2 confieren resistencia al aislado cubano de TYLCV2. A partir de esta información, se realizaron cruzamientos entre el cultivar susceptible ‘Amalia’, plenamente adaptado a las condiciones de cultivo de Cuba, y dos cultivares resistentes (‘LA3473’, portador de Ty-1; y ‘H-24’, portador de Ty-2), con la finalidad de incorporar estos genes de resistencia al cultivar susceptible. El objetivo de este trabajo fue establecer un criterio de selección fenotípico para incorporar la resistencia a TYLCV cuando no se disponga de facilidades para aplicar la selección asistida por marcadores.
MATERIALES Y MÉTODOS
Cultivares y poblaciones F2
Los cultivares que se tomaron como progenitores fueron: ‘Amalia’, cultivar comercial en la producción de tomate en Cuba (19), susceptible a TYLCV; ‘LA3473’, accesión tolerante a TYLCV, portadora del gen de resistencia Ty-1, introgresado desde la accesión ‘LA1969’ de S. chilense, (14) y ‘H-24’, proveniente de una línea cuya mejora se inició en la India (20), portadora del gen de resistencia Ty-2, introgresado desde la accesión B6013 de S. habrochaites f. glabratum (21). Se cruzó el cv. ‘Amalia’ con ‘LA3473’ y ‘H-24’ para obtener plantas F1, que luego de autofecundadas, derivaron en dos poblaciones F2 (TY-1F2, segregante para Ty-1; y TY-2F2, segregante para Ty-2). Se incluyó en el ensayo el cv. ‘STY1’, como control susceptible a TYLCV (22).
Virus, mantenimiento de la mosca e infección viral
El cultivo del aislado cubano, TYLCV-IL [CU] (7) fue mantenido en tomate donde se criaron las colonias de mosca blanca (B. tabaci, biotipo B), en una casa de cristal protegida contra insectos.
El procedimiento para la inoculación viral de las plantas se realizó según lo descrito por Lapidot et al (22); más de 50 moscas adultas que habían sido mantenidas en plantas infectadas con TYLCV-IL [CU] fueron colocadas sobre plantas de 25 días de sembradas, creciendo en cepellones que contenían sustrato a partir de suelo Nitisol ródico (23) y cachaza de caña de azúcar, como materia orgánica (2:1). Las plantas estuvieron expuestas a la inoculación del virus por las moscas virulíferas durante un período de 48 horas.
Las plantas inoculadas y sus controles (no expuestas a las moscas virulíferas) fueron sembradas en un sustrato de suelo Nitisol ródico y cachaza de caña de azúcar (3:1) contenido en canaletas de asbesto cemento, a una distancia de siembra de 0,25m entre plantas y 0,50m entre surcos. El control de los insectos fue mediante aplicaciones periódicas con el insecticida Confidor.
Evaluación de la severidad de los síntomas de TYLCV
La evaluación de los síntomas de la enfermedad causada por TYLCV se efectuó en 10 plantas de los cultivares (progenitores) y 75 plantas de las dos poblaciones F2. En los progenitores, a los 15 y 30 días posteriores a la inoculación y en las F2 a los 30 días. Se evaluó el índice de severidad de la enfermedad (DSI, «Disease Severity Index»), mediante una escala de 0-4 grados, propuesta por Friedmann et al. 1998, citados por Lapidot et al. (22):
0 ningún síntoma visible, las plantas inoculadas muestran el mismo crecimiento y desarrollo que las plantas no inoculadas.
1 muy ligero amarilleamiento de los márgenes de los foliolos de la hoja apical.
2 algún amarilleamiento y ligero encrespamiento de los extremos de los foliolos.
3 amplio rango de amarillaemiento de las hojas, encrespamiento y acucharamiento, con alguna reducción del tamaño, aunque las plantas continúan desarrollándose.
4 severa atrofia de la planta y amarilleamiento, encrespamiento y acucharamiento pronunciado de las hojas, detención del crecimiento de la planta.
Evaluación del contenido de ADN viral
Los begomovirus son virus de ADN (Ácido desoxirribonucleico) (24). Se extrajo el ADN de hojas jóvenes de las plantas, a los 30 días después de la inoculación. Para determinar la presencia y concentración de ADN viral se utilizó la técnica de hibridación de ácidos nucleicos (HAN) por la metodología de dot blot. Se aplicó el extracto de ADN sobre una membrana Hybond N+, con ayuda de un dispositivo comercial (Hibrid-Dot manifold, Gibco, EE.UU) y se fijó con luz UV (UV-crosslinker, E=120 mJ/cm2, Amersham Pharmacia Biotech, Reino Unido). La sonda empleada para la hibridación consistió en una secuencia de 500 nucleótidos del genoma de TYLCV, que incluye la región intergénica y el extremo 5’ del gen C1. Para la interpretación de los resultados se realizó una curva patrón a partir de concentraciones conocidas del genoma total de TYLCV-IL [CU] (10 ng.ìL-1, 1 ng.mL-1, 0.1 ng.mL-1 y 0.01 ng.mL-1) clonado en el plásmido PTY59 que sirvió de referencia para estimar visualmente el contenido del ADN viral en el tejido foliar de los cultivares inoculados3.
Los resultados fueron analizados mediante estadísticos descriptivos y distribución de frecuencia en las poblaciones TY-1F2 y TY-2F2 (SPSS Statistical Package).
RESULTADOS Y DISCUSIÓN
Evaluación de la resistencia a TYLCV en los progenitores
Transcurridos solo 15 días después de la inoculación, el cv. ‘STY1’, empleado como control susceptible, ya tenía síntomas intensos de TYLCD (grado 4, valor máximo de la escala), al igual que el cv. ‘Amalia’, de conocida susceptibilidad a la enfermedad causada por este virus. La rapidez en la aparición de los síntomas en estos confirmó la efectividad del procedimiento empleado para la inoculación con TYLCV.
En cambio, el cv. ‘H24’ no mostró ninguna de las sintomatologías descritas para la enfermedad causada por TYLCV (grado 0, valor mínimo de la escala). Sus foliolos apicales mantuvieron el color verde oscuro, característico de las hojas de las plantas sanas. Esta carencia de síntomas de la enfermedad se mantuvo luego de 30 días de efectuada la inoculación, al igual que la no detección de ADN viral en el tejido foliar apical.
Otros autores habían demostrado que la respuesta de ‘H-24’ ante TYLCV dependía del aislado. Ji et al (25) se refirieron a la resistencia específica de este cultivar a algunos aislados de TYLCV y el también begomovirus ToLCV (Tomato Leaf Curl Virus). ‘H-24’ mostró tolerancia a los aislados de TYLCV y ToLCV de Taiwán, el norte de Vietnam, el sur de la India e Israel; en cambio, fue susceptible a los aislados de TYLCV del norte de la India, Tailandia y Filipinas. Por su buen comportamiento frente a los aislados de Taiwán, ‘H-24’ ha sido empleado como progenitor resistente en los programas de mejora del tomate que se desarrollan en el AVRDC (26), aunque su respuesta de resistencia no fue informada como ausencia total de síntomas.
En la línea de mejora ‘LA3473’, que porta el gen de resistencia Ty-1, se observó solo un ligero amarillamiento de los foliolos apicales, similar a la descripción del grado 1 de la escala, valor que se mantuvo a los 30 días posteriores a la inoculación, siendo bajo el contenido de ADN viral (0,06 ng.mL-1, como valor promedio). Este gen fue el primero que se reportó para la resistencia a TYLCD, luego de ser introgresado a S. lycopersicum desde S. chilense (14), y el que le confirió la resistencia a los primeros híbridos resistentes de tomate que salieron al mercado, y es seguro que ha sido el más explotado comercialmente. En Cuba, el cv. ‘Vyta’, portador de Ty-1, lo que se ha confirmado mediante el marcador molecular TG97 (RFLP, «Restriction Fragment Length Polymorphism») (27), ha mostrado un buen nivel de resistencia, a pesar de haberse detectado una pequeña cantidad de ADN viral en sus plantas.
Evaluación de la resistencia a TYLCV en poblaciones F2
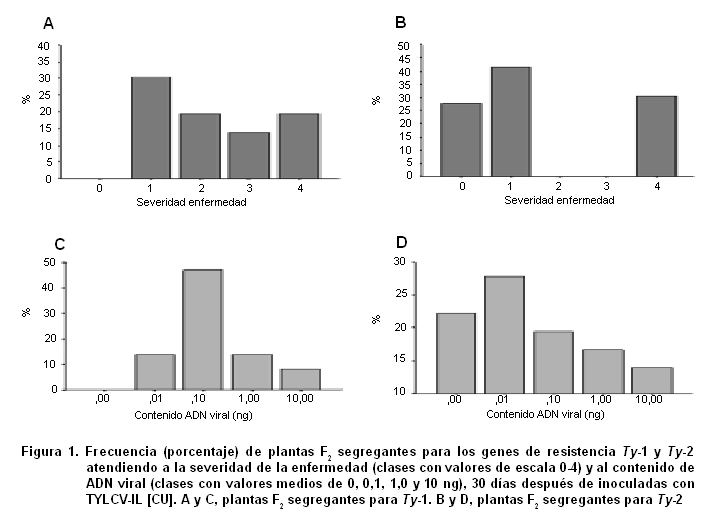
La expresión fenotípica de la sintomatología de la enfermedad fue diferente en las plantas de las poblaciones F2 segregantes para los genes Ty-1 y Ty-2. Las plantas segregantes para Ty-1 tuvieron la sintomatología que corresponde a un rango de valores desde 1 a 4 grados de la escala (Figura 1A), mientras que las plantas segregantes para Ty-2 mostraron una distribución discreta, con los síntomas correspondientes a los valores 0, 1 y 4 (Figura 1B).
Si se tiene en cuenta que el gen Ty-1 confiere resistencia del tipo dominante parcial (14), ello explicaría que la expresión de la resistencia abarcara todos los valores de la escala, excepto el grado 0 (que por otra parte tampoco lo alcanza el parental donante de la resistencia). En cambio, las plantas segregantes para Ty-2 tuvieron solamente los dos fenotipos parentales, 0 y 4, con una frecuencia, aproximada de ¼ y solamente un fenotipo nuevo, el grado 1, cerca del 42 % (1/2), supuestamente heterocigóticas, con un ligero amarilleamiento en las hojas apicales, apenas perceptible, que las distingue de las plantas del progenitor resistente, cv. ‘H-24’, que no mostraron cambios de coloración.
Scott (28) se refirió a que ‘H-24’ mostró resistencia dominante, de parcial a completa, frente a los aislados de Taiwán, y algunos síntomas de virosis ante los aislados de la India. En este trabajo, no se cuenta con todas las poblaciones que se requieren para un estudio de herencia más completo; sin embargo, se evidenció una expresión de dominancia casi completa en la F2 ante el aislado cubano, lo cual evidencia que la expresión genética de este gen difiere ante aislados de diferentes localidades.
Al analizar la distribución de frecuencia de las plantas de ambas poblaciones F2, de acuerdo a la concentración de ADN viral (Figura 1C y D), se observó una distribución desigual a la mostrada para la severidad de los síntomas. Esto pudiera explicarse porque el contenido de ADN viral es un carácter cuantitativo, con el que se expresa el nivel de replicación y/o movimiento sistémico del virus en la planta, mientras que la sintomatología es un carácter cualitativo, mediante el cual se evalúa la enfermedad, la afectación que provoca el virus en el crecimiento y desarrollo de la planta.
Se estableció el rango de variación del contenido de ADN viral que le corresponde a cada uno de los valores discretos de la escala (Tabla I), con el fin de determinar las plantas a seleccionar en las F2, atendiendo a ambos caracteres. Se aprecia que aquellas plantas con valor 1 y 4 de la escala tuvieron rangos semejantes, ya sean éstas de la población TY-2F2 o TY-1F2. El valor 0, ausencia total de mancha, se correspondió con ausencia de acumulación viral, al igual que su parental resistente, el cv. ‘H-24’ (Figura 2).

Este alto nivel de resistencia o respuesta de inmunidad en el cv. ‘H-24’ ante el aislado cubano de TYLCV, que ya había sido informada con anterioridad (Dueñas 2009, comunicación personal) en plantas evaluadas a los 45 días posteriores a la inoculación. Anteriormente, la «inmunidad» en el tomate ante TYLCV-IL [CU] había sido informada (29) en plantas genéticamente transformadas con una construcción genética asociada al gen de la proteína C1 en el fondo genético del cv. ‘Campbell-28’, que inducía el silenciamiento génico en la replicación temprana del TYLCV. En este trabajo se demuestra que este alto nivel de expresión de la resistencia ante el aislado cubano se puede encontrar en fuentes naturales de resistencia y que esta se hereda a ¼ de la población F2, lo que hace posible la selección fenotípica efectiva para este carácter.
Lapidot y Friedmann (30) se refirieron a la disparidad entre los fitopatólogos y mejoradores al definir el término ‘resistencia’. Estos autores consideran resistente: «aquella planta huésped que puede suprimir la multiplicación de un virus y, consecuentemente, suprimir el desarrollo de los síntomas de la enfermedad’.
Al igual, para Vidavsky (31), son ‘resistentes’ aquellas plantas sin síntomas ni ADN viral detectable; en cambio, son ‘tolerantes’ las que no muestran síntomas de la enfermedad, pero se detecta el ADN viral. Existen otros que consideran mas importante el efecto sobre el rendimiento de la planta (32), al clasificar las plantas acorde a los síntomas evaluados por la escala de 0-4 aquí también utilizada; siendo consideradas como resistentes parcialmente, aquellas con síntomas ligeros (<2), por no ocasionar pérdidas en el rendimiento, mientras que las plantas con 2 o más, como susceptibles.
Si tenemos en cuenta que los genotipos homocigóticos resistentes (progenitores) para Ty-2 y Ty-1 tuvieron valores 0 y 1 en la escala, respectivamente, las plantas a seleccionar en las poblaciones TY-2F2 y TY-1F2, deberán tener esos mismos valores, respectivamente. La fiabilidad en el empleo de la sintomatología para clasificar o seleccionar para la resistencia ha sido discutida. Con anterioridad, algunos autores (24, 33) encontraron una correlación positiva entre la severidad de los síntomas y la acumulación de ADN viral, sin embargo, no recomiendan emplear este último, como un solo indicador de la resistencia.
En este trabajo se evidenció una alta correlación positiva y altamente significativa** (P£0,01) entre la severidad de la enfermedad y el contenido de ADN viral; 0,726** en las plantas segregantes para Ty-1 y 0,932** en las segregantes para Ty-2 y se definieron los rangos de variación del contenido de ADN viral para cada grado de la escala. A consideración de los autores, existe una alta correlación fenotipo-genotipo, lo cual hace posible emplear la escala de severidad de síntomas en las primeras etapas de la selección de los genotipos homocigóticos resistentes, si se sigue cuidadosamente la misma metodología de inoculación y evaluación que se empleó en este trabajo.
Teniendo en cuenta los resultados, en el programa de retrocruzamiento y selección que se está llevando a cabo para la incorporación de los genes Ty-2 y Ty-1 al cultivar susceptible ‘Amalia’, para el primero solamente se deberán seleccionar aquellas plantas con grado 0, y para el gen Ty-1, deberán seleccionarse las que alcancen el valor 1 en la escala. Con esto, se garantizaría estar seleccionando las plantas resistentes homocigóticas, lo cual ahorra tiempo en la mejora para la resistencia por este método, cuando no sea posible emplear la selección asistida por marcadores ligados a estos genes.
REFERENCIAS
1. Briddon R. W.; Markham P. G. Family Geminiviridae. In: Murphy, F. A.; Fauquet, C. M.; Bishop, D. H. L.; Ghabrial, S. A.; Jarvis, A. W.; Martelli G. P.; Mayo, M. A.; Summers MD (eds), Virus Taxonomy: Classification and Nomenclature of Viruses. Sixth Report of the International Committee on Taxonomy of Viruses. Springer, Wien-NewYork, 1995, p. 158–165.
2. Cohen, S. y Antignus, Y. Tomato yellow leaf curl virus, a whitefly-borne geminivirus of tomatoes. Adv. Dis. Vector Res., 1994, vol. 10, p. 259-288.
3. Czosnek, H. y Laterrot, H. A worldwide survey of tomato yellow leaf curl viruses. Arch Virol., 1997, vol. 142, p. 1391–1406.
4. Moriones, E. y Navas-Castillo, J. Tomato yellow leaf curl virus, an emerging virus complex causing epidemics worldwide. Virus Res., 2000, vol. 71, p. 123-134.
5. Gómez, O.; Pinón, M.; Martýínez, Y.; Quiñónes, M.; Fonseca, D. y Laterrot H. Breeding for resistance to begomovirus in tropic-adapted tomato genotypes. Plant Breeding, 2004, vol. 123, p. 275-279.
6. Fauquet, C. M.; Briddon, R. W.; Brown, J. K.; Moriones, E.; Stanley J.; Zerbini M.; Zhou X. Geminivirus strain demarcation and nomenclature. Arch. Virol., 2008, vol. 153, p. 783-821.
7. Cohen, S. y Lapidot, M. Appearance and expansion of TYLCV:a historical point of view. En: Tomato Yellow Leaf Curl Virus Disease. Springer, Czosnek H. (ed.), 2007, p 3-12.
8. Gómez, P.; Rodríguez-Hernández, A. M.; Moury, B. y Aranda, M. A. Genetic resistance for the sustainable control of plant virus diseases: breeding, mechanisms and durability. Eur. J. Plant Pathol., published online: 2009, 09 April.
9. Scott, J. W.; Stevens, M. R.; Barten, J. H. M.; Thome, C. R.; Polston, J. E.; Schuster, D. J. y Serra, C. A. Introgression of resistance to whitefly-transmitted geminiviruses from Lycopersicon chilense to tomato. En: D. Gerling and R.T. Mayer (eds.) Bemisia, Taxonomy, Biology, Damage Control and Management. Intercept Press, Andover, UK. 1995, p. 357-367.
10. Vidavsky, F. y Czosnek, H. Tomato breeding lines immune and tolerant to tomato yellow leaf curl virus (TYLCV) issued from Lycopersicon hirsutum. Phytopathology, 1998, vol. 88, p. 910-914.
11. Vidavsky, F.; Czosnek, H.; Gazit, S.; Levy, D. y Lapidot, M. Pyramiding of genes conferring resistance to Tomato yellow leaf curl virus from different wild tomato species. Plant Breeding, 2008, vol. 127, p. 625-631.
12. Laterrot, H. Resistance genitors to tomato yellow leaf curl virus (TYLCV), Tomato Yellow Leaf Curl Newsletter, 1992, vol. 1, p. 2-4.
13. Agrama, H. A. y Scott, J. W. Quantitative trait loci for Tomato yellow leaf curl virus and Tomato mottle virus resistance in tomato. J. Am. Soc. Hortic. Sci., 2006, vol. 131, p. 267-272
14. Zamir, D.; Ekstein-Michelson, I.; Zacay, Y.; Navot, N.; Zeidan, M.; Sarfatti, M.; Eshed, Y.; Haret, E.; Pleban, T.; van Oss, H.; Kedar, N.; Rabinowitch, H. D. y Czosnek, H. Mapping and introgression of a tomato yellow leaf curl virus tolerance gene, Ty-1. Theoretical and Applied Genetics, 1994, vol. 88, p. 141-146.
15. Ji, Y.; Scott, J. W. y Maxwell, D. P. Molecular mapping of Ty-4, a new Tomato yellow leaf curl virus resistance locus on chromosome 3 of tomato. J. Amer. Soc. Hort. Sci., 2009, vol. 134, p. 281-288.
16. Hanson, P. M.; Bernacchi, D.; Green, S.; Tanksley, S. D.; Muniyappa, V.; Padmaja, A. S.; Chen, H. M.; Kuo, G.; Fang, D. y Chen, J. T. Mapping a wild tomato introgression associated with tomato yellow leaf curl virus resistance in a cultivated tomato line. J. Am. Soc. Hortic. Sci., 2000, vol. 125, p. 15-20.
17. Anbinder, I.; Reuveni, M.; Azari, R.; Paran, I.; Nahon, S.; Shlomo, H.; Chen, L.; Lapidot, M. y Levin, I. Molecular dissection of Tomato yellow leaf curl virus (TYLCV) resistance in tomato line TY172 derived from Solanum peruvianum. Theor. Appl. Gen., 2009, vol. 119, p. 519-530.
18. Friedmann, M.; Lapidot, M.; Cohen, S. y Pilowsky, M. A novel source of resistance to tomato yellow leaf curl virus exhibiting a symptomless reaction to viral infection. J. Am. Soc. Hortic. Sci., 1998, vol. 123, p. 1004-1007.
19. Alvarez, M.; Moya, C.; Dominí, M. E.; Sánchez, J. A.; Martínez, B.; Pérez, S. y Cuartero Zueco, J. ‘Amalia’, a medium fruit-size, heat-tolerant tomato cultivar for tropical conditions. HortScience, 2004, vol. 39, p. 1503-1504.
20. Kalloo, G.; Banerjee, M. K. Transfer of tomato leaf curl virus resistance from Lycopersicon hirsutum f. glabratum to L. esculentum. Plant Breed., 1990, vol. 105, p. 156-159.
21. Hanson, P. M.; Green, S. K. y Kuo, G. Ty-2, a gene on chromosome 11 conditioning geminivirus resistance in tomato. Rep. Tomato Genet. Coop., 2006, vol. 56, p. 17–18.
22. Lapidot, M.; Ben-Joseph, R.; Cohen, L.; Machbash, Z. y Levy , D. Development of a scale for evaluation of Tomato yellow leaf curl virus resistance level in tomato plants. Phytopathology, 2006, vol. 96, p. 1404-1408
23. Hernández, A.; Ascanio, M. O.; Morales, M. y Cabrera, A. Correlación de la nueva versión de de clasificación internacional de los suelos de Cuba con las clasificaciones internacionales y nacionales: Una herramienta para la investigación, docencia y producción agropecuaria. La Habana, Instituto Nacional de Ciencias Agrícolas (INCA), 2005, p. 60.
24. Rom, M.; Antignus, Y.; Gidoni, D.; Pilowsky, M. y Cohen, S. Accumulation of tomato yellow leaf curl virus DNA in tolerant and susceptible tomato lines. Plant Dis, 1993, vol. 77, p. 253-257.
25. Ji, Y.; Scott, J. W.; Hanson, P.; Graham, E.; y Maxwell, D. P. Sources of resistance, inheritance, and location of genetic loci conferring resistance to members of the tomato-infecting begomoviruses. En: Tomato yellow leaf curl virus disease: management, molecular biology and breeding for resistance. Czosnek H (ed), Springer, 2007, p. 343–362.
26. Hanson, P. TYLCV resistance in tomato. En: AVRDC Report 1998. Asian Vegetable Research and Development Center, Tainan, Taiwan, 1999.
27. Piñón, M.; Gómez, O.; Cornide, M. T. RFLP análisis of
Cuban tomato breeding lines with resistance to Tomato yellow leaf curl virus. Acta Hortic., 2005, vol. 695, p. 273-276.
28. Scott, J.W. Breeding for Resistance for virus pathogens. En: Genetic Improvement of Solanaceous Crops: Tomato. Ed. Science Publishers, Enfield, NH, USA. Razdan, M.K., Mattoo, A.K. (eds.), 2007, p. 457-485.
29. Fuentes, A.; Ramos, P. L.; Fiallo, E.; Callard, D.; Sánchez, Y.; Peral, R.; Rodríguez, R. y Pujol, M. Intron–hairpin RNA derived from replication associated protein C1 gene confers immunity to Tomato Yellow Leaf Curl Virus infection in transgenic tomato plants. Transgenic Research, 2006, vol. 15, p. 291–304.
30. Lapidot, M.; Friedmann, M. Breeding for resistance to whitefly-transmitted geminiviruses. Ann. Appl. Biol., 2002, vol. 140, p. 109-127.
31. Vidavsky F. Exploitation of resistance genes found in wild tomato species to produce resistant cultivars; pile up of resistant genes. En: Czosnek H (ed) Tomato yellow leaf curl virus disease: management, molecular biology and breeding for resistance. Springer, 2007. p. 363–372.
32. Pérez de Castro, A.; Díez, M. J. y Nuez, F. Inheritance of Tomato yellow leaf curl virus resistance derived from Solanum pimpinellifolium UPV16991. Plant Disease, 2007, vol. 91, p. 7.
33. Lapidot, M.; Friedmann, M.; Lachman, O. y Yehezkel, A. Comparison of resistance level to tomato yellow leaf curl virus among commercial cultivars and breeding lines. Plant Disease, 1997, 81:1425-1428.
Recibido: 30 de marzo de 2011
Aceptado: 17 de Julio de 2012
Dra.C. Marta Álvarez Gil, Investigador Titular del departamento de Genética y Mejoramiento Vegetal, Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, Mayabeque. Email: malvarez@inca.edu.cu


{kind=link}
{kind=link}