










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCultivos Tropicales
versión impresa ISSN 0258-5936
cultrop vol.34 no.2 La Habana abr.-jun. 2013
Experiencias obtenidas en el desarrollo participativo de híbridos lineales simples de maíz (Zea mays, L.) en condiciones de bajos insumos agrícolas
Field’s experiences on participatory development of simple maize’s hybrid, in agricultural low input conditions
Ernesto M. Ferro Valdés,I M.Sc. Eliecer Chirino González,II Maikel Márquez Serrano,II Erick Mirabal Báez,II Dr.C. Humberto Ríos Labrada,III Dr. Francisco Guevara Hernández,IV Francisco Alfaro HernándezV
IProfesor Auxiliar de la Universidad de Pinar del Río, Facultad de Agronomía de Montaña, Canalete, San Andrés, La Palma, Pinar del Río, Cuba,
CP 24310.
IIProfesores Asistentes de la Universidad de Pinar del Río, Facultad de Agronomía de Montaña, Canalete, San Andrés, La Palma, Pinar del Río, Cuba,
CP 24310.
IIIProfesor de la Universidad de la Habana, CEDEM, Centro de Estudios Demográficos, Ciudad de la Habana.
IVUniversidad Autónoma de Chiapas, Chiapas, México.
VCampesino experimentador, CCS «Pedro Lantigua», La Palma, Pinar del Río.
RESUMEN
Fue conducida una investigación con el objetivo de evaluar una metodología participativa para el desarrollo de híbridos heteróticos de maíz en condiciones de bajos insumos agroquímicos, en la finca de un productor del municipio La Palma, provincia de Pinar del Río. Se utilizaron como progenitores las diez variedades que más votos recibieron durante una feria de diversidad de maíz ejecutada con este fin. Se realizaron dos ciclos de autofecundación, bajo las condiciones fitotécnicas acostumbradas en la región y además se realizaron mediciones de ocho descriptores. Se analizó la carga recesiva en la generación F1 y se calculó depresión consanguínea para cada generación. Se procesaron los datos utilizando Excel y SPSS 11.5. Se encontró, que las líneas no soportaron la depresión consanguínea en F2 bajo el ambiente de manejo de limitados insumos agroquímicos y la generación F1 obtenida de progenitores locales, mostraron menor depresión que las obtenidas de progenitores de otras provincias del país.
Palabras clave: maíz, Zea mays, autofecundación, híbridos, mejora genética.
ABSTRACT
An investigation was conducted with the objective to evaluate a participatory methodology for maize’s hybrid development, in agricultural low inputs conditions at farmer’s property in La Palma, province of Pinar del Río. Were used as progenitors, ten varieties more votes received during the maize diversity’s fair. Two self fecundation cycles was performed as well as the evaluation of eight describers. The recessive charge was analyzed in F1 and consanguineous depression was evaluated for each generation. All data were processed using Excel and SPSS 11.5. It was found that lines didn’t tolerate the consanguineous depression in the second generation in low use of agricultural inputs, and F1 generation from local progenitors, manifest lower depression that those obtained from progenitors from other provinces of the country.
Key words: maize, Zea mays, self-fertilization, hybrids, genetic gain.
INTRODUCCIÓN
Las razas cubanas de maíz (Zea mays L.) constituyen un potencial de gran valor genético para Cuba y el Caribe, ya que poseen buena habilidad combinatoria con otras fuentes de germoplasma para la obtención de híbridos (1, 2, 3, 4, 5).
Antes de los híbridos, el mejoramiento de maíz se basaba en la selección masiva de variedades de polinización abierta. La mayor parte de esta selección era hecha por los mismos productores y la selección realizada por estaciones experimentales y por investigadores de compañías de semillas, era solo una pequeña parte del panorama. Una de las principales actividades del departamento de agricultura de los EE.UU. era la coordinación de una red de productores para seleccionar las mejores mazorcas polinizadas en sus lotes de maíz (6).
Tratando de mantener este modelo, los primeros investigadores de híbridos de maíz, dedicaron considerable tiempo intentando desarrollar formas en que los productores pudieran generar y producir sus propios híbridos (6). Sin embargo, tras descubrir el potencial valor de la heterosis, los programas de mejora de maíz han tendido a concentrarse en el desarrollo de híbridos, donde la mayor cantidad de trabajos de mejoramiento de maíz, se han llevado a cabo por compañías privadas orientadas a obtener ganancias, para quien los híbridos son una proposición comercial mucho más atractiva que las variedades de polinización libre (7), ya que estos pueden producir rendimientos de 20 a 30 % mayores que los de las variedades (8), y además, imposibilitaba el mantenimiento de la semilla por los mismos productores.
No obstante, la mayor efectividad de la hibridación de líneas resulta afectada por el encarecimiento que representa la inversión adicional necesaria para la obtención y mejora de las líneas (9). El alto costo de producir la semilla híbrida presenta un desafío final para los programas de mejora de maíz, porque incluso cuando es posible desarrollar híbridos que significativamente se desempeñan mejor que las variedades de los campesinos, frecuentemente no es posible producir la semilla mejorada a un precio que los agricultores enfocados a la subsistencia, se sientan deseosos y puedan pagar por ella (7).
Otro elemento que complejiza estos programas de mejora, tiene que ver con los múltiples fines de uso del cultivo. Los mejoradores enfrentan el desafío adicional de tener que desarrollar muchos tipos diferentes de variedades para cubrir las preferencias varietales de los productores (10), las que por demás, deben poseer las capacidades para adaptarse a las variables condiciones medioambientales donde estas se cultivan.
A pesar de las ventajas productivas que presentan los maíces híbridos y que el uso de germoplasma mejorado ha aumentado en América Latina de conjunto, este solo se encuentra concentrado en áreas donde el mismo es un cultivo comercial, mientras que en las regiones caracterizadas por una agricultura orientada al autoconsumo, la mayoría de los agricultores continúan cultivando variedades locales (7).
En el caso puntual del municipio La Palma, las semillas de maíz proveniente del sector formal cubano, han sido poco comercializadas y no alcanzan a cubrir el 5 % del área cultivada anualmente. Este fenómeno en especial ha sido más notable con la semilla de maíz híbrido, la que históricamente ha sido menos comercializada en este sistema de producción y a su vez, es la que mayores precios de venta ha presentado (11).
Valorando la posibilidad de aprovechar el vigor híbrido desde formas más descentralizadas y justas socioeconómicas y ambientalmente, se considera evaluar el posible desarrollo de híbridos simples de maíz de mayor potencial productivo que los materiales locales, de buena adaptación y de mayor nivel de adopción en la región, utilizando una metodología de desarrollo que incluyera aspectos de mejora de plantas clásica y participativa, siendo la intención de este trabajo evaluar una metodología para el desarrollo participativo de híbridos lineales simples de maíz, en condiciones de bajos insumos agroquímicos y bajos costos económicos.
MATERIALES Y MÉTODOS
Con el objetivo de desarrollar híbridos simples de maíz, durante los años 2004 y 2005, se desarrolló una investigación participativa en la finca del campesino experimentador Francisco Alfaro Hernández, de la Cooperativa de Créditos y Servicios (CCS) «Pedro Lantigua» del municipio La Palma.
El suelo sobre el que se realizó la experiencia es clasificado como un Ferralítico Rojo Lixiviado (12), con una pendiente de un 2 % y 2.18 % de contenido de materia orgánica, con un Ph que oscila entre 6.5 y 6.7. Se utilizó en todos los experimentos, el marco de plantación 0,50 m entre plantas y 0,80 m entre surcos o hileras. Las atenciones culturales se efectuaron según las realizadas comúnmente por los productores en la región, dos riegos en la campaña y un deshierbe. La fertilización fue de 70 kg.ha-1, de una mezcla compuesta por el 20 % de NPK y 80 % de abonos orgánicos.
Se utilizó la palabra locales para referenciarse a las variedades procedentes del municipio La Palma. Igualmente, se utilizó el término ambiente meta, para referirse a las finca de los productores y ambientes convencionales de desarrollo, a los contextos donde se aplican los paquetes tecnológicos completos para el desarrollo del cultivo, o para la obtención de variedades mejoradas (Tabla I). Maíz criollo es un término campesino que comúnmente se emplea para denotar que es un material nativo de la comunidad, región, estado o país, y que se diferencia de un material foráneo, un maíz híbrido o una variedad mejorada (13).
La elección de los diez progenitores, fue llevada a cabo a partir de los resultados de la selección de variedades de maíz, en una feria de diversidad (14) realizada sobre este cultivo. En este evento, se expusieron a la libre evaluación de 83 productores palmeros, 55 materiales de seis provincias. Los 15 materiales que más votos recibieron en la feria de diversidad de maíz fueron objeto de discriminación, para los que se utilizaron los siguientes criterios:
que fuesen de los que más votos recibieron según los criterios de aceptación de los productores. Este elemento se consideró para asegurar en los híbridos, la mayor cantidad de genes portadores de las características aceptadas por los consumidores.
que cinco fueran locales y cinco no locales, ya que en el cruzamiento de pares de procedencia ecológica diferente podría lograrse repetidamente buena heterosis del rendimiento de los granos (15).
Se utilizaron ocho descriptores cuantitativos para la evaluación de todos los experimentos (Tabla II). Para ello se siguieron las indicaciones de IPGRI (16). Para la evaluación de las variedades, se utilizó una muestra de diez plantas por parcelas, utilizando el muestreo sistemático (17).

Descripción del método
Se seleccionaron los diez materiales más gustados de los 55 materiales expuestos para la selección. De estos diez materiales, cinco fueron locales y cinco no locales, los que se almacenaron para posteriormente ser sembrados en la próxima campaña. Posteriormente se desarrolló un experimento con 150 plantas por variedad en un diseño completamente al azar, bajo siembras de 0,50 x 0,80 m. Se les midieron los ocho descriptores antes mencionados y se seleccionaron de ellas las 30 mejores plantas por variedad para ser autofecundadas, siguiendo el método estándar para obtener líneas consanguíneas (9).
Luego fueron llevadas a otro experimento los diez padres más las 20 líneas (dos líneas por padre) obtenidas en la primera autofecundación, a todos se les midió los ocho descriptores antes mencionados. Se sembraron 150 plantas por líneas bajo siembras de 0,50 x 0,80 m.
A las líneas se les midió también el porcentaje de carga recesiva; o sea, la cantidad de variaciones fenotípicas encontradas, asociadas a efectos de genes recesivos en la población de estudio cuantificado a partir del número de plantas por líneas que mostraban efectos recesivos como deficiencia de clorofila y achaparramiento, en relación al total de plantas por línea.
Se midió además el por ciento de depresión consanguínea de estas líneas siguiendo los criterios de Sigarroa (18), y por último en la época de primavera se sembraron nuevamente las plantas de la F2 que soportaron la depresión consanguínea junto a sus progenitores.
A los padres se les midió nuevamente los ocho descriptores antes mencionados y a la F2 se les midió la altura de la planta. El rendimiento calculado se realizó a un 12 % de humedad.
Análisis de los datos
Se computó la cantidad de expresiones recesivas observadas por las líneas obtenidas a partir de un mismo progenitor, determinando la ocurrencia de efectos recesivos en una muestra de 150 plantas por líneas. Se llevó a cabo análisis de comparación de proporciones entre las líneas de un mismo progenitor para determinar diferencias significativas entre estas. Se realizaron pruebas de t entre las líneas y entre estas y sus progenitores, comparando los resultados del rendimiento. Para ello se utilizó el programa SPSS 11.5.
Se calculó además, la frecuencia génica y genotípica utilizando la fórmula planteada por Hardy Weimberg, así como la depresión consanguínea (15, 18). De igual manera se realizó un análisis de comparación múltiple, para determinar las medias que eran significativamente diferentes por progenitor, acorde al número de expresiones recesivas cuantificadas en sus líneas. Se utilizó para ello Statgraphic 5.1.
RESULTADOS Y DISCUSIÓN
Carga recesiva
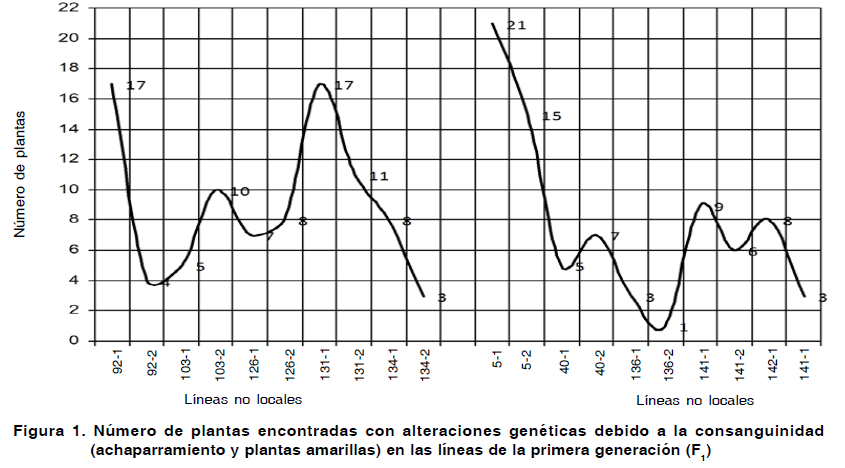
Debido a que en las primeras generaciones autofecundadas aparece un gran número de tipos de letales y subvitales (19), se cuantificaron las manifestaciones de estos en la F1 y se graficaron en la Figura 1.
Como se puede observar en la Figura 1, los efectos de los alelos recesivos fueron notables atendiendo al número de alteraciones fenotípicas encontradas por la consanguinidad en la F1. Cada valor graficado, representa la suma de plantas con achaparramiento y plantas amarillas encontradas en las 150 plantas de la población de cada una de las 20 líneas.
Evaluando de manera conjunta la F1, podremos observar que las líneas provenientes de materiales de otras localidades presentaron 90 variaciones fenotípicas visibles y 78 se desarrollaron en las líneas de origen local, o sea, 12 menos. No obstante, el análisis de comparación de proporciones no muestra existencia de diferencias significativas, acorde al número de alteraciones encontradas en correspondencia con el origen de las líneas.
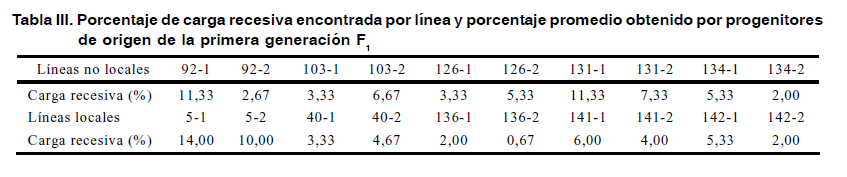
Sin embargo, cuando analizamos las afectaciones por líneas, encontramos que algunas se destacan por presentar una mayor carga de recesivos que otras. El número de manifestaciones recesivas osciló en el rango desde 1 (0,67 %) a 21 (14 %) por línea, destacándose cuatro de ellas por mostrar entre 15 a 21 plantas afectadas de un total de 150. En ese caso se encuentran las líneas 5-1, 5-2, 131-1, 92-1 (Figura 1 y Tabla III).
Las líneas que presentaron mayor depresión provienen de variedades campesinas, pero en contraposición a ello, las que menos carga recesiva manifestaron, entre 0,67 y el 2 % de los individuos de su población, también fueron de origen campesino. Las líneas de los progenitores provenientes del sistema formal de semilla (var. 103, Francisco mejorado y var. 126, VST-6), se comportan similar a las restantes diez líneas de origen campesino de valor medio (entre 2 y 10 %) conforme a la carga recesiva que manifiestan en la F1. Las dos líneas procedentes del material progenitor (var. 5), sumaron la mayor cantidad de plantas con afectaciones de cargas recesivas, 33 individuos con achaparramiento y tres amarillas, para un total de 36 plantas afectadas de 300. Esto lleva a suponer una mayor acumulación de recesivos en este progenitor que en los restantes.
Si se une las afectaciones ocurridas en las líneas de este progenitor 5 (3 amarillas y 33 plantas achaparradas), apreciamos la mayor carga recesiva de todos los progenitores utilizados en esta experiencia.
Estos resultados apuntan a niveles significativos de endogamia dentro de este material, lo cual pudiera guardar estrecha relación con la ubicación geográfica de la finca, dentro del contexto productivo local (al interior de un sistema de intercambio natural o a uno de sus bordes) y a la frecuencia con que los productores donantes de estas semillas, realicen intercambio con otros productores.
Se observó en la primera generación, que las plantas amarillas aparecieron en menor frecuencia que las plantas con achaparramiento. Se encontraron cuatro plantas amarillas en una población de 3000 y 164 con claras manifestaciones de achaparramiento en la misma, lo que da una distribución de una planta amarilla por cada 750 plantas y una con achaparramiento de cada 18.
La frecuencia génica de los recesivos para las plantas amarillas en la población F1 fue de 0,037, con la probabilidad de existencia de 211 plantas heterocigóticas para este carácter. Sin embargo, la frecuencia de este recesivo en las líneas procedentes de la var. 5 fue de 0,10, con una posibilidad de que 54 individuos en esa población sean heterocigóticos.
La prueba de comparación múltiple de muestras, arrojó la existencia de diferencias significativas entre el progenitor 5 para el número de afectaciones genéticas encontradas por efecto de la consanguinidad en la F1, con el resto de las variedades para un 95 % de significación, exceptuando con las variedades 131 y 92 (Figura 2).
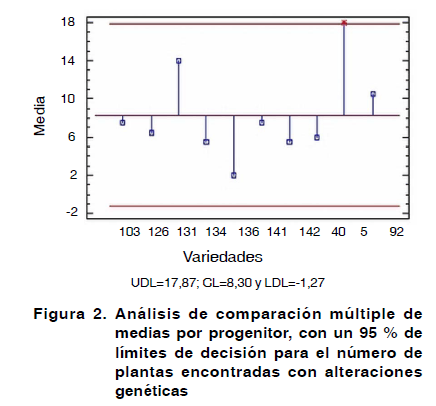
La prueba múltiple de rangos, muestra la formación de tres grupos por orden de significación. El primero compuesto por la variedad 5, 131 y 92 en disposición de importancia y el segundo compuesto por las variedades 103, 141, 126, 40, 134 y 142. En el tercero se encuentra la variedad 136, la que por demás, presentó claras diferencias con la variedad 131. Esta finalmente fue la que presentó diferencias con todas las demás variedades, ya que fue la que menos expresiones recesivas mostró.
Los resultados de la comparación de proporciones entre las líneas de un mismo padre, muestra la no existencia de diferencias significativas entre las manifestaciones recesivas de ambas líneas, con excepción del progenitor 92, quien muestra diferencias significativas para el 99 %. En general, este resultado apunta que en un 90 % de los casos, ocurre una distribución homogénea de los recesivos entre las líneas en formación provenientes de un mismo progenitor.
Depresión consanguínea
La máxima expresión de la heterosis (vigor híbrido) se manifiesta en el híbrido simple (20) y se define como el evidente incremento del vigor, en relación con el promedio de los padres o del progenitor superior, de la descendencia obtenida del cruzamiento de dos genotipos diferentes (21). Por su parte, la depresión consanguínea es el proceso inverso y se ha definido como la reducción en la medida fenotípica del carácter estudiado (18) en cruzamientos emparentados.
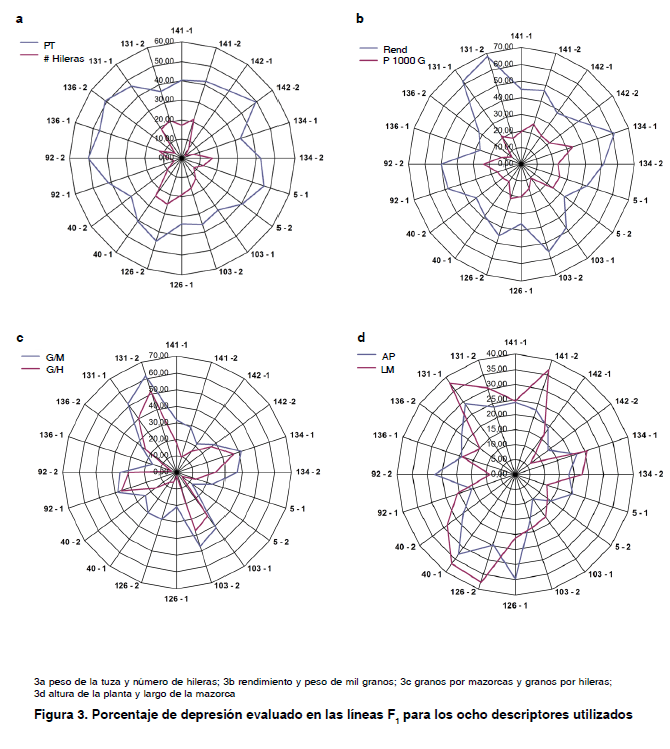
Los porcentajes de depresión consanguínea registrados en la F1, se grafica en la Figura 3. Se puede apreciar en las Figuras 3a y 3b que el número de hileras y el peso de mil granos fueron los menos afectados por la depresión consanguínea, ya que muestran los porcentajes más bajos. Para número de hileras los valores oscilaron entre 2,14 y 25 %, con un promedio general de 13,14 % de reducción y para el peso de mil granos el rango de valores se mantuvo entre 7,66 y 33,38 %, con un promedio de 19,21 %.
Por otra parte, rendimiento y peso de la tusa fueron los caracteres que mayores porcentajes de reducción manifestaron. Como promedio estos caracteres manifestaron en el orden antes mencionados 45,10 y 41,35 % de reducción fenotípica. Los restantes caracteres se comportaron en promedio de la siguiente forma: granos por mazorcas (32,01 %), largo de la mazorca (22,35 %), altura de la planta (21,54 %), granos por hileras (21,52 %).
Cabe señalar que granos por mazorcas y granos por hileras, muestran los valores de mayor variación entre los mayores y menores porcentajes de depresión, siendo las líneas de variedades no locales, con excepción de las líneas del progenitor 126, las que más depresión mostraron para ambos caracteres. Caso similar ocurrió con el rendimiento, todas las líneas de origen no locales mostraron mayor depresión que las líneas provenientes de progenitores locales.
Como se puede observar en la Figura 3, las líneas de la F1 para el rendimiento muestran porcentajes de depresión diferentes entre sí. El valor de mayor depresión lo muestra la línea 131-2 con un 67,64 % y la línea de menor valor de depresión para el rendimiento fue la 136-1 con 26.62 %.
La prueba de t realizada al rendimiento entre los padres y sus líneas arrojó diferencias para el 99 % de significación en todos los casos y al realizarla entre las líneas de una misma variedad, fue en todos los casos no significativa.
La depresión promedio en el rendimiento de las líneas provenientes de variedades no locales fue de 52,16 % y para las líneas de variedades locales de 38,04 %. Este resultado nos sugiere que las líneas de variedades locales amortiguan mejor la depresión consanguínea para este carácter.
Los porcentajes de depresión consanguínea evaluado por carácter, para cada grupo de origen, nos muestran en la Figura 4, como de forma general las líneas de progenitores locales se deprimen en menor porcentaje en todos los caracteres, que las originadas de progenitores no locales, con la excepción del carácter peso de la tusa, donde las locales cuantificaron un 43,09 % de reducción frente al 39,61 % que mostraron las no locales.
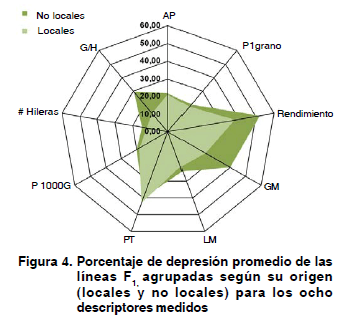
Se cumplió lo planteado por Cornide, cuando expresó, que tras aplicar un proceso de consanguinidad, se encontrará un mayor efecto depresivo en los caracteres ligados al valor adaptativo (18). No obstante, las diferencias encontradas en los resultados de la depresión entre ambos grupos de líneas, pudieran estar condicionadas fuertemente debido a que las líneas locales poseen en su dotación genética, una base hereditaria mejor ajustada a los rigores de los ambientes donde se desarrollan, como resultado del proceso de selección realizado por los campesinos, por muchos años.
Cambios sustanciales evolutivos pueden inicialmente ocurrir con la selección de pocos genes (22), ya que unos pocos mayores influyen a menudo en una gran cantidad de la variabilidad genética, incluso, aunque un número alto de genes pueda afectarse por la selección artificial durante la domesticación (23).
Este resultado se apoya, en los resultados encontrados por varios investigadores, sobre el comportamiento diferencial de los híbridos y sus progenitores en distintos ambientes (21, 24).
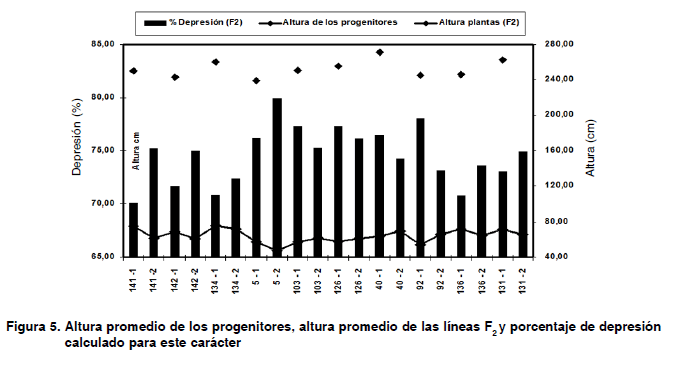
Otro planteamiento que se cumplió en esta experiencia, elude que el vigor y fecundidad de muchas líneas disminuyen hasta el punto que estas no pueden conservarse ni en condiciones óptimas de cultivo (19). En la segunda generación (F2) solo se pudo medir altura de las plantas, debido a que ninguna de las 3000 plantas de esta generación, logró sobrepasar el ciclo reproductivo exitosamente.
Los valores de depresión para la altura de las plantas se muestran en la Figura 5, se registraron en el rango de 70,01 a 79,92 % de reducción del valor fenotípico con respecto a sus padres.
Líneas de la F2
Como se planteó anteriormente, las líneas no sobrevivieron para proseguir a otro ciclo de autofecundación, detuvieron su crecimiento vegetativo tempranamente hasta su paulatina muerte fisiológica. Solo apenas unas decenas de plantas, con inflorescencias mal desarrolladas se cuantificaron.
Durante el período de desarrollo de la población F2, las lluvias acumuladas fue el único factor que se registró fuera del rango adecuado para el desarrollo del cultivo. Se acumularon únicamente 225,70 mm, lo que representó del 37,11 al 49 por ciento de los 460 a 600 mm que se ha planteado por diversos investigadores (25, 26, 27), como los óptimos para este cultivo. No obstante a ello, se realizaron dos riegos durante la campaña para cubrir el déficit de las lluvias. Los demás parámetros climáticos y medio ambientales como temperatura, humedad relativa, pH del suelo y porcentaje de materia orgánica, oscilaron en el rango adecuado.
Descartando el factor medio ambiental, como el elemento determinante del marcado deterioro de las líneas en F2, nos quedaría el factor asociado al grado de consanguinidad y su relación con el ambiente de desarrollo.
Las líneas, en el ambiente convencional de desarrollo, reciben suministro adicional de nutrientes a base de fertilizantes químicos, lo que en cierta medida ayuda a amortiguar la depresión consanguínea.
En este caso, la fertilización se realizó utilizando bajas dosis de químicos en mezcla con abono orgánico, en pos de provocar una más ajustada respuesta del genotipo a las condiciones reales de cultivo localmente.
Debido a la naturaleza heterogénea del cultivo, todas las líneas poseen desigual grado de consanguinidad. En la Tabla IV, intentamos resumir las manifestaciones fenotípicas esperadas, a partir de las relaciones que se establecen entre el grado de consanguinidad de las líneas y el ambiente de desarrollo. Se pudieran formar tres grandes grupos para explicar los fenómenos relacionados con la expresión diferenciada de los individuos durante el proceso de autofecundación en dos ambientes contrastantes (uno de alto uso de agroquímicos y otro de bajo uso de estos).
Se considera que en un experimento con líneas en formación, en diferentes ambientes de desarrollo (convencional y bajos insumos), se debería esperar porcentajes de resultados de sobrevivencia, similares a los que se muestran en la Tabla IV. Esto ocurrirá, siempre y cuando, los tres grupos de carga recesiva planteados en la tabla, se distribuyan de forma equitativa en el total de los individuos de la población.
Los grupos 1C, 2C y 3C, corresponden a la respuesta a esperar por las líneas en ambientes de desarrollo donde reciben todas las atenciones culturales recomendadas para el cultivo, con fuertes cargas de energía externa.
El grupo 1C es aquel en el que las líneas poseen baja carga recesiva y le son suministrados, además, fertilizantes químicos que aseguran su supervivencia. Estas líneas normalmente sobreviven hasta su uso en las pruebas combinatorias.
Se tiene también al grupo 2C, que presenta grados medios en su carga recesiva; son individuos que muestran reducción en sus fenotipos, pero con adecuadas atenciones culturales y en especial, con provisiones de fertilizantes, logran sobrevivir.
Y por último el grupo 3C, que presenta una gran carga recesiva, con importantes afectaciones morfofisiológicas letales, que inclusive con apoyo de los fertilizantes químicos, no logra pasar al nuevo ciclo.
En los grupos 1B, 2B y 3B, son ambientes de cultivo donde las líneas dan una respuesta de supervivencia acorde a atenciones de baja carga de fertilizantes químicos. En el grupo 1B las líneas poseen baja carga de recesivos, lo que le posibilita, aun en ausencia nutricional química adicional, sobrevivir gracias a su constitución genotípica favorable. El grupo 3B muere por los desvíos fisiológicos producidos por la alta carga recesiva que posee y el grupo 2B tiende a morir por su carga recesiva de valor medio, que en este ambiente de ausencia de apoyos externos asegura su muerte.
De manera general, se puede esperar un 66,66 % de supervivencia en las líneas, de los ambientes favorecidos con insumos externos y un 33,33 % en el ambiente de bajos insumos externos, para poblaciones donde se cumpla el factor de distribución equitativa del grado de carga recesiva en los grupos de la población total. La diferencia entre el resultado a esperar en ambos ambientes, lo marca esencialmente el grupo de líneas de cargas recesivas medias, las que por su constitución genotípica tras la autofecundación, pudiesen sobrepasar el estado depresivo con la ayuda de insumos externos.
CONCLUSIONES
El desarrollo de híbridos en condiciones de bajos insumos, reduce la posibilidad de éxito con respecto al ambiente de mejora convencional, debido a la baja sobrevivencia de las líneas por los efectos combinados de la carga recesiva y la carencia de agroquímicos.
En un 90 % de los casos, las líneas de un mismo progenitor tienden a comportarse lo más homogéneas posibles en relación con la cantidad de manifestaciones recesivas que muestran.
A pesar de mostrar mayor carga recesiva, las variedades locales se deprimen menos que los materiales no locales.
REFERENCIAS
1. Fernández, L.; Crossa, J.; Fundora, Z. M. y Gálvez, G. Caracterización de razas cubanas de maíz (Zea mays L.) mediante marcadores agromorfológicos en la colección nacional del cultivo. Cultivos Tropicales, 2009, vol. 30, no. 4, p. 62-70.
2. Betran, F. J.; Mayfield, K. y Menz, M. Breeding Maize Exotic Germplasm». Plant Breeding, 2006, p. 352-367.
3. Hallauer, A. R. Plant Breeding: The Arnell R. Hallauer International Symposium 2003, México D.F. 2006.
4. Sevilla, R. Colecta y clasificación para programar la conservación in situ de la diversidad de maíz en la Amazonía peruana, en fundamentos genéticos y socioeconómicos para analizar la agrobiodiversidad en la región Ucayali. Puclallpa, Perú. Biodiversity International, 2006, p. 33-50.
5. Taba, S.; Chávez, V.; Krakowsky, F. Rincon y Dreisigacker, S. Tropical Late Yellow Maize Germplasm Development Using Improved Lines and Gene Bank Core Accessions». CIMMYT. 2006.
6. DeBoer L. J. Camino Sinuoso en la Adopción de Agricultura de Precisión. [Consultado: 25 octubre de 2007]. Disponible en: <http://www.agriculturadeprecision.org/analecon/CaminoSinuoso.htm> .
7. Morris, M. L. y López Pereira, M. A. Impactos del mejoramiento de maíz en América Latina, 1966-1997. México, D.F. : CIMMYT. 2000. p. 55.
8. Pérez Ponce, J. N. y Rodríguez Fuentes, C. Producción de semillas y propágulos. Editorial Pueblo y Educación. Ciudad de la Habana. 1989. p. 269.
9. Rodríguez Fuentes, C. /et al./. Mejora de plantas. Editorial Félix Varela. Ciudad de la Habana. 2006. p. 293.
10. Córdova S. H.; Barreto J. H. y Crossa J. Impacto del desarrollo de híbridos de maíz en Centro América. Rev. Agronomía Mesoamericana, 1994, vol. 5, p. 78-87.
11. Ferro, E. M. /et al./. Aporte del sistema formal en semillas mejoradas de granos y cereales a la seguridad alimentaria del municipio La Palma, provincia Pinar del Río. Cultivos Tropicales, 2009, vol. 30, no. 2, p. 59-65.
12. Hernández, A. /et al./. Claves para evaluar la nueva versión de clasificación genética de los suelos de Cuba. 1999. 73 p.
13. Acosta, R. El cultivo del maíz, su origen y clasificación. El maíz en Cuba. Cultivos Tropicales, 2009, vol. 30, no. 2, p. 113-120.
14. De La Fe, C. /et al./. Las Ferias de Agrodiversidad. Guía Metodológica para su organización y desarrollo en Cuba. Instituto Nacional de Ciencias Agrícolas. La Habana : Ediciones_INCA. 2003. 24 p. ISBN 959-7023-23-7.
15. Pérez, P. J. /et al./. Genética y mejoramiento de plantas tropicales. Tomo II. Ediciones EMPES. La Habana. 1991. p. 75-100.
16. IBPGR. Descriptors for Maize. International Maize and Wheat Improvement Center, México City/International Board. Plant. Genetic Resource, 2001. p. 261.
17. González, O. L. Experimentación forestal. Imprenta «Andre Voisin» Ministerio de Educación Superior. La Habana. 1986. p. 313.
18. Cornide, María Teresa /et al./. Genética vegetal y Fitomejoramiento, Editorial Científico-Técnica. La Habana. 1985. p. 639.
19. Allard, R. W. Principios de la mejora genética de las plantas. Capítulo 18: Aspectos generales de la heterosis y de la depresión producida por la consanguinidad. Ciudad de la Habana : Editorial Pueblo y Educación. 1977. p. 228.
20. Bejerano, Arnoldo. Híbridos simples: una alternativa promisoria para el cultivo de maíz (Zea mays L.) en Venezuela. [Consultado: 17 mayo de 2006]. Disponible en: <http://www.ceniap.gov.ve/bdigital/congresos/vjornada/tablacontenido.htm#Bejarano> .
21. Chassaigne, A. y Borges, O. Evaluación de la heterosis entre poblaciones de maíz (Zea mays L.) con distinta dureza de grano. Investigación Agrícola [En línea]. [Consultado: 20 noviembre de 2009]. Disponible en: <http://www.redpav-fpolar.info.ve/danac viewarticle.php?id=14> .
22. Hancock, James F. Contributions of Domesticated Plant Studies to our Understanding of Plant Evolution. Annals of Botany, 2005, vol. 96, p. 953-963.
23. Wright S. I.; Bi, I. V.; Schoeder, S. G.; Yamasaki, M.; Doebley, J. F.; McMullen, M. D. y Gaut, B. S. The effects of artificial selection on the maize genome. Science, 2005, vol. 308, p. 1310-1314.
24. Alejos, G.; Monasterio, P. y Rea R. Análisis de la interacción genotipo ambiente para el rendimiento de maíz en la región maicera del estado Yaracuy, Venezuela. Agronomía Tropical, 2006, vol. 56, no. 3, p. 369-384.
25. Pedro, H. M. /et al./. El cultivo de maíz y las condiciones climáticas. [Consultado: 30 octubre de 2009]. Disponible en: <http://www.engormix.com/el_cultivo_maiz_condiciones_s_articulos_699_AGR.htm> .
26. Leonard, David. Una introducción a los cultivos individuales. [Consultado: mayo 9 de 2008]. Disponible en: <http://media.payson.tulane.edu:8083/html/spanish/pc/m0035s/m0035s08.htm> .
27. Socorro Quesada, M. A y Martín Fagundo, D. S. Granos. Impreso en los Talleres Gráficos de la Dirección de Publicaciones y Materiales Educativos del Instituto Politécnico Nacional. México DF. México. 1998. p. 318.
Recibido: 8 de noviembre de 2011
Aceptado: 5 de octubre de 2012
Ernesto M. Ferro Valdés, Profesor Auxiliar de la Universidad de Pinar del Río, Facultad de Agronomía de Montaña, Canalete, San Andrés, La Palma, Pinar del Río, Cuba, CP 24310. Email: emferro@af.upr.edu.cu, eliecer@af.upr.edu.cu

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}