










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCultivos Tropicales
versión impresa ISSN 0258-5936
cultrop vol.35 no.1 La Habana ene.-mar. 2014
Determinación de estabilidad genética en vitroplantas de clavel español (Dianthus caryophyllus L.), micropropagadas con Biobras-16
Determination of genetic stability in plants of Spanish carnation (Dianthus caryophyllus L.), micropropagated with Biobras-16
MSc. Yanelis Castilla Valdés,I Dra. María E. González Vega,II MSc. Regla M. Lara RodríguezIII
IAspirante a Investigador del departamento de Genética y Mejoramiento, Instituto Nacional de Ciencias Agrícolas (INCA), carretera a Tapaste, km 3½, gaveta postal 1, CP 32 700, San José de las Lajas, Mayabeque, Cuba.
IIInvestigador Titular del departamento de Genética y Mejoramiento, Instituto Nacional de Ciencias Agrícolas (INCA), carretera a Tapaste, km 3½, gaveta postal 1, CP 32 700, San José de las Lajas, Mayabeque, Cuba.
IIIEspecialista del departamento de Genética y Mejoramiento, Instituto Nacional de Ciencias Agrícolas (INCA), carretera a Tapaste, km 3½, gaveta postal 1, CP 32 700, San José de las Lajas, Mayabeque, Cuba.
RESUMEN
El clavel español (Dianthus caryophyllus L.) se encuentra entre las especies de flores y plantas ornamentales más solicitadas por la población cubana. Su propagación in vitro con el empleo del análogo de brasinoesteroide Biobras-16, producido en Cuba, permite obtener un elevado coeficiente de multiplicación. Sin embargo, algunas de las técnicas del cultivo in vitro y algunos de los biorreguladores empleados en estas, pueden introducir alteraciones en el material hereditario, por lo que resulta importante determinar si se mantuvo estable genéticamente. En el presente trabajo nos propusimos el objetivo de estudiar la estabilidad genética a través del empleo de marcadores bioquímicos, en vitroplantas de clavel español micropropagadas con diferentes concentraciones de Biobras-16 como sustituto de la citoquinina. El material vegetal fue obtenido a partir del cultivo de meristemos en medios con Biobras-16 como sustituto de la Kinetina, a las concentraciones de 0,1; 0,01 y 0,001 mg.L-1. Las plantas crecidas en cada uno de estos tratamientos a los tres meses fueron multiplicadas por esquejes durante dos subcultivos, sin el empleo de este análogo de brasinoesteroide. Para la preparación de las muestras del estudio isoenzimático, fueron tomadas hojas de plantas de los diferentes tratamientos del cultivo de meristemos y las propagaciones por esquejes, y fueron analizados los sistemas isoenzimáticos Peroxidasas, Fosfatasas Ácidas, Malato Deshidrogenasas y Esterasas. Las electroforesis se realizaron en geles de poliacrilamida (PAGE) en lámina vertical. Como resultado, aunque el sistema isoenzimático Fosfatasas Ácidas resultó polimórfico, de manera general se determinó que se mantuvo la estabilidad genética de las plántulas regeneradas.
Palabras clave: isoenzimas, Cuba, flores, plantas ornamentales.
ABSTRACT
The spanish carnation (Dianthus caryophyllus L.) is one of the flowers and ornamental plants most requested by the Cuban population. The in vitro propagation of this species with the employment of the analogous of brassinoesteroid Biobras-16, produced in Cuba, allows to obtain a high coefficient of multiplication. However, some of the in vitro culture techniques and some of the bioregulators employed, can introduce alterations in the genetic material, so it turns out necessary to supervise if it kept genetically stable. For these reasons, in the present work we proposed the aim to study the genetic stability with the employment of biochemical markers, of plants of carnation micropropagated with different concentrations of Biobras-16 in substitution of cytoquinin. The vegetal material was obtained from the meristems culture of spanish carnation vitroplants, in medium with Biobras-16 as a substitute of cytoquinin, at the concentrations of 0,1; 0,01 y 0,001 mg.L-1. The plants grown in each of these treatments, three months later were multiplied by cuts during two subcultures, without Biobras-16. For the sample preparation of the isoenzymatic study, it was taken leaves of plants of the different treatments of the meristems culture and the cutting propagations and it was studied the systems Peroxidases, Acid Phosphatases, Malate Dehydrogenases and Esterases. The electrophoresis were made on vertical slab polyacrylamide gels (PAGE). As result, although the Acid Phosphatases isoenzymatic system turned out to be polymorphic, in a general way it is considered that it was kept the genetic stability of the regenerated plants.
Key words: isoenzymes, Cuba, flowers, ornamental plants.
INTRODUCCIÓN
En la actualidad, a nivel mundial, la floricultura ha dejado de ser la simple afición de cultivar flores para convertirse en un negocio de grandes proporciones, y la exportación internacional de flores frescas ha ascendido a 7 390 millones de dólares (1). De esta cifra, los claveles representan aproximadamente el 6 %, pues se encuentran entre las flores más cotizadas debido a su belleza, su duración después de cortados, su posibilidad de florecer todo el año y su resistencia al embalaje y a la transportación. Los claveles que se cultivan en Cuba se destinan al mercado nacional, aunque su demanda no se encuentra satisfecha debido principalmente a la ausencia de métodos de propagación seguros y eficientes, ya que en las condiciones climáticas de la isla, el clavel español (Dianthus caryophyllus L.) generalmente no produce semillas y se multiplica a partir de esquejes, lo que facilita la diseminación de algunas plagas y enfermedadesA.
Los métodos de propagación in vitro permiten superar algunos de estos inconvenientes, como por ejemplo: la obtención de plantas genéticamente idénticas, la modificación de las condiciones ambientales, y la obtención de plantas libres de patógenosB. Además, resulta posible incrementar los coeficientes de multiplicación de las vitroplantas, con el empleo en los medios de cultivo de hormonas del crecimiento vegetal o de sustancias que tengan una actividad biológica semejante a estas, como es el caso de los análogos de brasinoesteroides (2). Entre ellos el Biobras-16, producido en Cuba, ha sido utilizado durante diferentes fases de la micropropagación de cultivos tan diversos como la piña (Ananas comosus) y el corojo (Acrocomia aculeata) (3, 4) con resultados satisfactorios.
Sin embargo, es conocido que además de los beneficios que se obtienen con la aplicación de la propagación in vitro y el uso de nuevos biorreguladores en los medios de cultivo, estos pueden introducir alteraciones en el material hereditario, como microlesiones y macrolesiones (5), por lo que un requerimiento importante para que el proceso de regeneración in vitro sea exitoso es que las plantas regeneradas presenten estabilidad genética, comparadas con las plantas propagadas convencionalmente (6). La electroforesis de isoenzimas, como un estudio directo del producto de los genes, permite la observación de cambios somáticos, ya que la pérdida o aparición de bandas isoenzimáticas puede indicar la presencia de mutaciones debidas a la recombinación mitótica, deleciones, duplicaciones, cambios de bases o eventos de transposición (7). Estos estudios, entre otros, permiten detectar algunas de estas alteraciones, además de que la metodología empleada para su análisis es rápida, sencilla y económica con respecto a las técnicas moleculares (8, 9).
El estudio de la detección de estabilidad genética mediante marcadores isoenzimáticos en vitroplantas de clavel español micropropagadas con el empleo de análogos de brasinoesteroides, constituye un tema de investigación novedoso a nivel nacional e internacional, ya que aunque el cultivo in vitro de esta especie ha sido ampliamente estudiado (10, 11), los antecedentes acerca de los estudios isoenzimáticos son escasos y se refieren principalmente a la detección de genotipos en cruzamientos intraespecíficos (12, 13). En Cuba se han realizado estudios acerca de la micropropagación del clavel español y el empleo con este fin de otro análogo de brasinoesteroide, el Biobras-6 (14, 15); en cambio, estos estudios no incluyeron la detección de estabilidad genética. También pueden citarse otros resultados con respecto a la micropropagación del clavel español con el empleo del Biobras-16 como sustituto de auxinas y el estudio isoenzimático de las vitroplantas obtenidas, así como el empleo de dicho biorregulador como sustituto de citoquininas (16, 17, 18), en cambio, hasta el momento no se ha realizado el estudio isoenzimático para la detección de estabilidad genética en las plantas, en estas últimas condiciones experimentales.
Por todo ello, el presente trabajo tuvo como objetivo estudiar la estabilidad genética, a través del empleo de marcadores bioquímicos, en vitroplantas de clavel español micropropagadas con diferentes concentraciones de Biobras-16 como sustituto de la citoquinina.
MATERIALES Y MÉTODOS
Material vegetal
Primeramente se describe el experimento a partir del cual fueron obtenidas las vitroplantas de clavel español (Dianthus caryophyllus L.), de las que se tomaron las muestras foliares para el estudio isoenzimático.
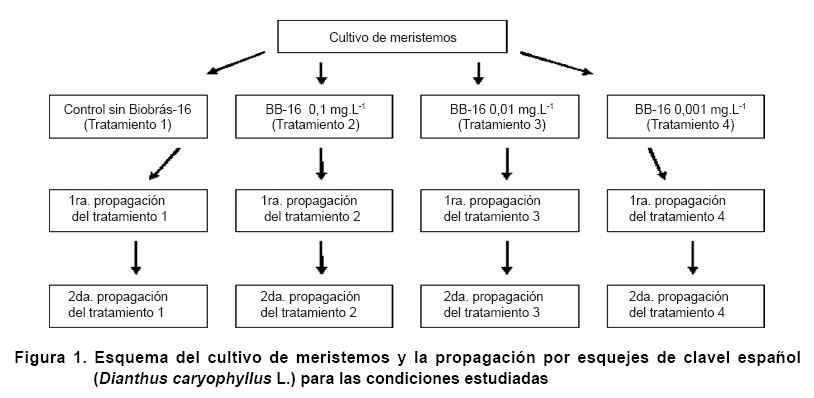
Plantas de clavel español previamente obtenidas in vitro a partir de la germinación de semillas, fueron tomadas como donantes de meristemos de 0,1 cm de longitud, los que fueron sembrados en medio de cultivo MS (19) con ácido indol-3 acético (AIA) 1,5 mg·L-1, Ácido Giberélico (AG3) 1 mg.L-1 y Kinetina (KIN) 0,8 mg.L-1 (tratamiento 1 o control) y en tres variantes de este medio, sustituyendo la KIN por diferentes concentraciones de Biobras-16: 0,1 mg.L-1 (tratamiento 2); 0,01 mg.L-1 (tratamiento 3) y 0,001 mg.L-1 (tratamiento 4). A los tres meses del establecimiento del cultivo de meristemos, a las plantas obtenidas de cada una de las variantes descritas anteriormente les fueron tomados todos los esquejes y fueron sembrados en medio de cultivo control, pero manteniendo los tratamientos de que provenían, y a su vez, a los tres meses estas plantas fueron multiplicadas por esquejes, considerándose esta como la segunda propagación (Figura 1) (18).
Estudio isoenzimático
Para comprobar la estabilidad genética de las plantas obtenidas a partir del cultivo de meristemos en los diferentes tratamientos y las respectivas propagaciones realizadas, se desarrolló un estudio isoenzimático para los sistemas peroxidasas (PER), fosfatasas ácidas (APS), malato deshidrogenasas (MDH) y esterasas (EST).
Para la preparación de las muestras, fueron tomadas las plántulas de tres meses de crecimiento, obtenidas a partir del cultivo de meristemos en los tratamientos 1, 2, 3, 4 y sus correspondientes propagaciones (Tabla I). A partir de estas plántulas fueron seleccionadas las hojas totalmente extendidas, provenientes de los tres nudos más cercanos al ápice y se prepararon seis muestras por cada tratamiento para los diferentes análisis.

Para la obtención de los extractos crudos, se maceró un gramo de hojas por cada muestra, para lo cual se utilizó un mortero frío, con cinco gotas de solución de sacarosa al 20 % y luego se centrifugó a 20000 r.p.m. Posteriormente se tomó el sobrenadante y fue envasado en viales y conservado a -15°C hasta su análisis.
Las corridas electroforéticas se realizaron en gel de poliacrilamida (PAGE) al 8,5 %, a una intensidad de corriente constante de 25 mA, en una cámara de electroforesis en lámina vertical Mighty Small II de Pharmacia Biotech y utilizando un buffer de corrida de Tris-glicina 0.025 M y pH=8.3 (20). Se realizaron seis repeticiones de cada análisis isoenzimático.
El tiempo de corrida se estableció según el desplazamiento de la banda de Kohlrausch hasta aproximadamente 8,5 cm del inicio.
Se aplicaron tinciones específicas con el objetivo de visualizar las bandas de cada sistema isoenzimático estudiado (Tabla II).
Después de las tinciones, los geles fueron lavados con agua destilada y mantenidos en una solución de ácido acético glacial al 7 %, hasta el momento de confeccionar los electroforetogramas.
La posición relativa de cada banda (Rf) fue establecida sobre la base de la distancia media de migración obtenida, dividida entre la distancia de migración del frente de corrida.
RESULTADOS Y DISCUSIÓN
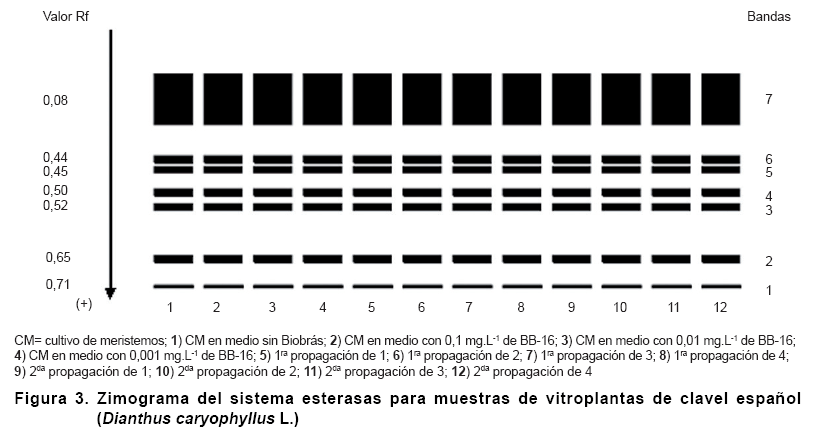
Durante el estudio para el sistema peroxidasas se presentaron un total de siete bandas, las cuales fueron comunes en posición e intensidad para todas las muestras analizadas (Figura 2).
Las Peroxidasas son un grupo de isoenzimas con participación activa en la lignificación de la pared celular y la regulación de los niveles de auxina, además, pueden estar relacionadas con el desarrollo de otros estados fisiológicos en las plantas, como la defensa contra infecciones, la curación de heridas y la biosíntesis del etileno (7). Estas isoenzimas son consideradas marcadores bioquímicos importantes de la morfogénesis y han sido empleadas para estudiar el efecto de oligosacarinas y brasinoesteroides en otros cultivos de interésC.
En estudios precedentes, se ha informado sobre la presencia de siete bandas de Peroxidasas en el cultivo de meristemos y las propagaciones por esquejes de clavel español, con el empleo del Biobras-16 como sustituto de auxinas (17).
En este caso, aunque algunas de las bandas polimórficas estuvieron asociadas a la presencia de este análogo de brasinoesteroide en el medio de cultivo, de manera general fue determinado que el empleo del Biobras-16 en el cultivo de meristemos, y el número de propagaciones realizadas, no afectaron la estabilidad genética del material.
En la Figura 3 se presenta el zimograma del sistema esterasas; donde se obtuvo la presencia de siete bandas y se presentó un marcado monomorfismo ya que en todos los tratamientos evaluados hubo igual posición, número e intensidad de bandas.
Semejantes resultados obtuvieron otros autores al realizar el estudio isoenzimático del sistema esterasas durante la micropropagación del cultivo de la yuca (Manihot esculenta Crantz), utilizando diferentes variantes de medios de cultivo con el empleo de una mezcla de oligogalacturónidos como sustituto total y parcial de auxinas, donde fueron observadas seis bandas comunes a todos los tratamientosD.
Por otra parte, en un estudio similar se encontró cierto grado de polimorfismo en las esterasas al comparar plántulas de clavel chino (Dianthus chinensis L.) obtenidas a partir de la germinación in vitro y las propagaciones con el empleo de diferentes concentraciones de Biobras-16; sin embargo, de manera general no se presentó variabilidad genética significativa en los tratamientos analizadosE.
Las esterasas son isoenzimas que juegan un importante papel en los procesos fotosintéticos de las plantas y su estabilidad en la expresión enzimática les confiere gran importancia en estudios genéticos, por lo que son ampliamente utilizadas para estudiar los diferentes estadios de desarrollo (7).
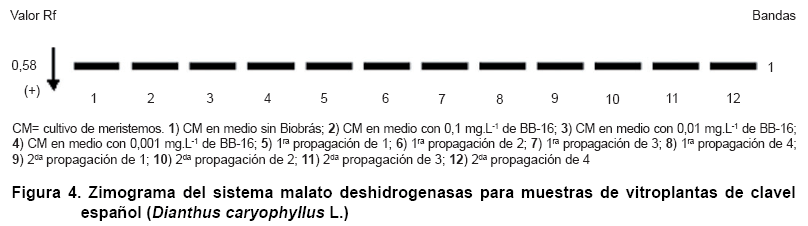
De igual manera, se observó un marcado monomorfismo en el sistema Malato deshidrogenasas, pero en este caso con la presencia de una banda común en todas las muestras estudiadas (Figura 4).
Otros autores al estudiar este sistema isoenzimático en el cultivo del tomate (Solanum lycopersicum L.), también encontraron similitud electroforética. Al monitorear la estabilidad genética de plantas de esta especie obtenidas in vitro, determinaron que el material evaluado presentó similaridad electroforética para diferentes sistemas evaluados, incluyendo el de la MDH (23).
Por otra parte, en plantas de yuca obtenidas a partir del cultivo in vitro con el empleo de un análogo de oligogalacturónido, también se encontró un patrón común de MDH para todos los tratamientos estudiadosD.
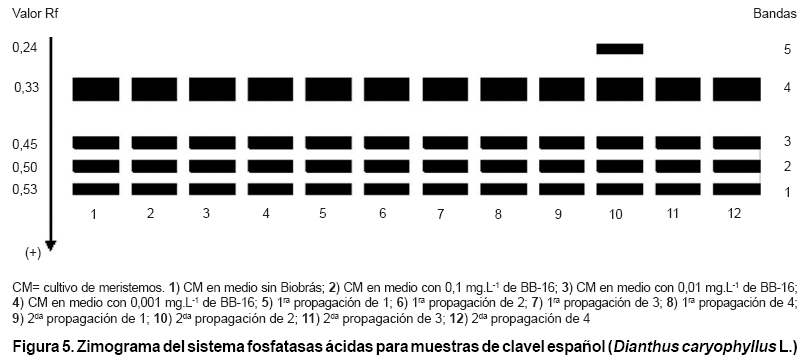
En el análisis del sistema Fosfatasas Ácidas (Figura 5) se observó un total de cinco bandas, cuatro de ellas comunes en posición e intensidad en todos los tratamientos. Sin embargo, en la muestra número 10, correspondiente a la segunda propagación de las plantas provenientes del cultivo de meristemos en el medio con 0,1 mg.L-1 de Biobras-16, se apreció una banda que no apareció en el resto de las muestras analizadas.
Este comportamiento pudo estar asociado con cierta tendencia observada en plantas procedentes de este tratamiento que se caracterizó por ser el de mayor concentración de Biobras-16, provocando un incremento en las variables morfológicas evaluadas, lo que puede haber ocasionado un aumento en la demanda de las sales y agua presentes en el medio, causando a su vez agotamiento de estas sustancias en el medio de cultivo. Precisamente pudiera atribuirse a este agotamiento en el medio de cultivo el surgimiento de un estado de estrés en las plantas, que quizás indujo la expresión de esta nueva banda de APS, ya que según lo señalado por otros investigadores, las fosfatasas ácidas son enzimas que se encuentran ampliamente distribuidas en las plantas y se han informado incrementos de las mismas frente a diferentes tipos de estrés abióticos, tales como el estrés salino, osmótico e hídrico (7).
Por otra parte, se considera que la tendencia al incremento en las variables morfológicas evaluadas en las plantas sometidas a dicho tratamiento, se relaciona con el hecho de que los brasinoesteroides ejercen marcados efectos sobre el crecimiento de plantas estresadas, por lo que han sido considerados como hormonas del estrés (2). Es de destacar que aunque el Biobras-16 no fue utilizado en el medio de cultivo para la propagación por esquejes del clavel español, posiblemente su efecto se manifestó en este subcultivo ya que los análogos de brasinoesteroides presentan una actividad a largo plazo (long lasting activity) con respecto a los brasinoesteroides naturales, debido a diferencias estructurales entre ellos, que pueden conllevar a que transcurra determinado período de tiempo antes de que aquellos sean transformados a formas biológicamente activasF. Resultados semejantes se obtuvieron en la micropropagación del clavel español utilizando el Biobras-16 como sustituto de auxinas o como único suplemento hormonal, donde se observó que el efecto promotor del crecimiento de este análogo de brasinoesteroide, se manifestó durante el segundo subcultivo (16).
Por tanto, se considera que la estabilidad genética de las plantas micropropagadas no se encuentra influida por la banda de APS que apareció en la muestra número 10, sino que al parecer es el resultado de una variación epigenética detectada por el estudio isoenzimático, pues las isoenzimas son marcadores de expresión génica, que están influidas por las condiciones ambientales o de cultivo (7). En subcultivos posteriores se detectó que las variables morfológicas de las plantas provenientes de este tratamiento, mantuvieron sus valores promedio.
En estudios realizados en el clavel chino, también ha sido descrita la aparición de bandas polimórficas de APS durante la evaluación de la variabilidad y la estabilidad genética de plántulas micropropagadas con el empleo de diferentes concentraciones de Biobras-16. En este caso se observó la desaparición de bandas en algunos de los tratamientos en los que se utilizó este análogo de brasinoesteroideE. Otros autores han encontrado polimorfismo para el sistema APS al evaluar la actividad del Pectimorf en la embriogénesis somática de mandarina Cleopatra (Citrus reshni Hort. ex Tan) y plantean que estas isoenzimas juegan un papel importante en el ciclo biogeoquímico del fósforo orgánico y en la nutrición de las plantas (24).
De manera general, el análisis de los sistemas isoenzimáticos estudiados, permitió corroborar que no se produjo variabilidad genética en las plantas de los diferentes tratamientos del cultivo de meristemos y sus correspondientes propagaciones, en comparación con el control, por lo que se considera que tanto el empleo del Biobras-16 como los ciclos multiplicativos evaluados, no fueron causa de variaciones en las plantas de clavel español micropropagadas. Otros autores, al estudiar los sistemas peroxidasa, esterasas, anhidrasa carbónica y polifenol oxidasas para evaluar la estabilidad genética en accesiones de papa (Solanum spp.) regeneradas a partir de la conservación in vitro por 10 años con el empleo de manitol, no detectaron cambios en los patrones de bandas de las accesiones conservadas in vitro, en comparación con las mantenidas en el campo (25). Sin embargo, al realizar el análisis molecular por AFLP sí fueron detectados cambios que se produjeron a nivel de ácido desoxirribonucleico (ADN), aunque los índices de similitud entre los patrones de banda de cada accesión se mantuvieron elevados, por lo que resulta importante también aplicar técnicas más novedosas y avanzadas que complementen los resultados del estudio bioquímico.
Por otra parte, al comprobar la estabilidad genética de las plantas de clavel español obtenidas en este estudio, bajo las condiciones señaladas y mediante los análisis isoenzimáticos evaluados, es posible proponer su propagación a mayor escala, ya que un requisito indispensable para que un sistema de cultivo in vitro sea aceptado para la propagación masiva de plantas, lo constituye la garantía de su estabilidad genética. Además, estos resultados resultan de particular importancia para la validación del Biobras-16 como un biorregulador que, a las concentraciones estudiadas, no provoca cambios genéticos en las plantas del cultivo in vitro. Al constituir un análogo de brasinoesteroide que se produce en Cuba, su empleo en lugar de fitohormonas de importación resulta muy ventajoso, a la vez que su obtención resulta menos tóxica y más económica que la de otros análogos como el Biobras-6F.
CONCLUSIONES
En la mayoría de los sistemas enzimáticos estudiados no se detectó la presencia de cambios genéticos en las vitroplantas obtenidas a partir del cultivo de meristemos y las propagaciones por esquejes.
REFERENCIAS
1. Traub, A. Las flores de corte en una nueva disyuntiva: ¿por cuál camino transitar? Oficina de Estudios y Políticas Agrarias, Chile. 2010. [Consultado: 16/febrero/2011]. Disponible en: http://www.odepa.gov.cl.
2. Mazorra, L. M. y Núñez, M. Estado actual sobre el conocimiento de la biosíntesis y los mecanismos moleculares de acción de los brasinoesteroides en las plantas. Cultivos Tropicales, 2008, vol. 29, no. 1, p. 91-105.
3. Catunda, P. H. A.; Marinho, C. S.; Gomes, M. M. de A. y de Carvalho, A. J. C. Brassinosteroid and substrates in acclimatization of 'Imperial' pineapple. Acta Scientiarium-Agronomy, 2008, vol. 30, no. 3, p. 345-352.
4. Borcioni, E. y Bonato, R. R. Aplicação de análogo de brassinosteroide (Biobras 16®) sobre a germinação e crescimento in vitro de embriões zigóticos e aclimatização de plântulas de bocaiuva. Ciencia Rural. 2012, vol. 42, no. 2, p. 270-275.
5. Hughes, K. Ornamental species. En: B.V. Conger. Cloning agricultural plants via in vitro techniques. Boca Ratón, Florida: CRC Press, Inc., 1981, 273 p.
6. Marín, E. A.; Vargas, T. E. y Oropeza, M. Variabilidad genética y anatomía foliar comparada de plantas de Dioscorea alata mantenidas en cultivo in vitro. Interciencia, 2012, vol. 37, no. 6, p. 476- 483.
7. González, C. Detección del polimorfismo genético mediante marcadores bioquímicos en plantas. En: Cornide, Ma. Teresa. Marcadores moleculares, nuevos horizontes en la genética y la selección de las plantas. La Habana: Editorial Féliz Varela. 2002, 366 p.
8. Valdés, M.; González, C.; Lara, R. M.; Román, M. I.; Hernández, Y.; Hernández, R. M.; Cabrera, M. y Torrecilla, G. Diversidad genética de especies silvestres del género Nicotiana I: caracterización mediante marcadores bioquímicos. Rev. Protección Veg., 2010, vol. 25, no. 2, p. 88-97.
9. Giraldo, M. C.; Ligarreto, G. A.; Cayón, G. y Melo, C. Análisis de la variabilidad genética de la colección colombiana de musáceas usando marcadores isoenzimáticos. Acta Agronómica, 2011, vol. 60, no. 2, p. 108-119.
10. Ali, A.; Afrasiab, H.; Naz, S.; Rauf, M. e Iqbal, J. An efficient protocol for in vitro propagation of carnation (Dianthus caryophyllus L.). Pakistan Journal of Botany, 2008, vol. 40, no. 1, p. 111-121.
11. Casas, J. L.; Olmos, E. y Piqueras, A. In vitro propagation of carnation (Dianthus caryophyllus L.). En: Mohan Jain, S. y Ochatt, Sergio J. Protocols for in vitro propagation of ornamental plants. 2010, 586 p.
12. Messeguer, R. y Arús, P. Electrophoretic identification of carnation cultivars. HortScience, 1985, vol. 20, no. 3, p. 372-373.
13. Messeguer, R. y Arús, P. Genetics of isozyme polymorfisms in carnation. Journal of Heredity, 1995, vol. 87, no. 2, p. 112-118.
14. Montes, S.; Ramírez, L.; Hernández, M. M.; Santana, N.; Martínez, M. y Lara, D. Micropropagación de variedades de clavel (Dianthus caryophyllus L. y Dianthus plumarius l.) mediante el cultivo in vitro de meristemos. Cultivos Tropicales, 1997, vol. 18, no. 1, p. 75-81.
15. Montes, S.; Mesa, O.; Hernández, M. M.; Santana, N.; Núñez, M. y Varela, M. Uso del análogo de brasinoesteroide BB-6 en la micropropagación del clavel. Cultivos Tropicales, 1997, vol. 18, no. 2, p. 51-55.
16. Castilla, Y.; Pinares, A.; Xiqués, X.; Pérez, D. y Torres, M. Efecto del Biobras-16 en la propagación in vitro del clavel español (Dianthus caryophyllus L.). En: Taller de Biotecnología Vegetal (2004, 9 al 12 de noviembre: La Habana). Memorias del XIV Congreso Científico del INCA. La Habana, 2004. ISBN 959-7023-27-X.
17. Castilla, Y.; Pinares, A. y Xiqués, X. Estudio citogenético e isoenzimático de plántulas de clavel español (Dianthus caryophyllus L.) obtenidas por el cultivo in vitro de meristemos. En: Botánica Económica y Biotecnología Vegetal (Comisión 2: 2005, 20 al 22 de enero: Camagüey). Memorias del IX Encuentro de Botánica Johannes Bisse in Memoriam. Camagüey, 2005. ISBN 959-18-0005-3.
18. Castilla, Y. y González, M. E. Efecto de un análogo de brasinoesteroide en la propagación por esquejes de clavel español (Dianthus caryophyllus L.) a partir de plantas obtenidas por cultivo de meristemos. Temas de Ciencia y Tecnología, 2011, vol. 15, no. 44, p. 31-36.
19. Murashige, T. y Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiology Plantarum, 1962, vol. 15, p. 473-497.
20. Laemmli, U. K. Cleavage of structural proteins during assembly of the head bacteriophage T4. Nature, 1970, vol. 227, p. 680-685.
21. Wendel, J. F. y Weeden, N. F. Visualization and interpretation of plant isozymes. En: Isozymes in Plant Biology. New York. Dioscorides Press. 1989, p. 5-34.
22. Vallejos, E. Enzyme activity staining. En: Tanskley, S. y Orton, T. Isozimes in plant genetics and breeding, 1983, 516 p.
23. Lara, R. M.; Florido, M.; Plana, D.; Moré, O.; González, M. E.; Álvarez, M. y Hernández, M. M. Isoenzymatic analysis for detecting in vitro variability and/or stability of economically important crops. Cultivos Tropicales, 2003, vol. 24, no. 3, p. 39-47.
24. Hernández, R. M.; Lara, R. M.; Diosdado, E.; Cabrera, J. C.; González, C.; Valdés, M. y Xiqués, X. Evaluación de la efectividad del Pectimorf en la embriogénesis somática de mandarina Cleopatra (Citrus reshni Hort. ex Tan). Cultivos Tropicales. 2007, vol. 28, no. 4, p. 25-31.
25. Castillo, J. G.; Estévez, A.; Salomón, J. L.; Vargas, D.; Hernández, M. M.; Pérez, A.; Borrás-Hidalgo, O.; López, Y. y Arencibia, A. D. Determinación de la estabilidad genética en cuatro especies del banco de germoplasma de papa en Cuba conservadas in vitro. Cultivos Tropicales, 2010, vol. 31, no. 3, p. 51-57.
Recibido: 16 de noviembre de 2012
Aceptado: 1 de marzo de 2013
MSc. Yanelis Castilla Valdés, Aspirante a Investigador del departamento de Genética y Mejoramiento, Instituto Nacional de Ciencias Agrícolas (INCA), carretera a Tapaste, km 3½, gaveta postal 1, CP 32 700, San José de las Lajas, Mayabeque, Cuba. Email: yanelis@inca.edu.cu


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}