










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCultivos Tropicales
versión impresa ISSN 0258-5936
cultrop vol.35 no.1 La Habana ene.-mar. 2014
Nod factor detection in B. elkanii ICA 8001. Culture medium influence
Detección de factores Nod In. B. elkanii ICA 8001. Influencia del medio de cultivo
Dra.C. María C. Nápoles García,I Dr.C. Juan C. Cabrera,II Dr.C. Ellen Luyten,III Dr.C. Bruno Dombrecht,III Dr.C. Jos Vanderleyden,III Dra.C. Josefa Hormaza,IV Dr.C. Armando Rodríguez AlfonsoV
IInvestigador Titular del departamento de Fisiología y Bioquímica Vegetal, Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, Mayabeque, Cuba.
IIUnité de Biotechnologie (UBT). Belgique.
IIIF. A. Janssens Laboratory of Genetics. Catholic University of Leuven. Belgium.
IVInstituto de Investigaciones Científicas del Azúcar, La Habana, Cuba.
VCentro de Productos Marinos, La Habana, Cuba.
ABSTRACT
Nodulation factors, synthesized by different species from Rhizobiaceae family, have been described as key signals in leguminous interaction. They constitute morphogens that elicit the nodular program in the plant, allowing the bacteria to enter the roots and showing a positive influence on bacteroids development and nitrogen fixation efficiency. This study is related to the detection, by means of two chromatographic techniques, of Nod factors production by Bradyrhizobium elkanii ICA 8001 strain, cultivated in different culture media, and the biological effect of inocula in the interaction with plant. Both methods demonstrated that the culture medium composition induce in a higher or lower quantity the Nod factors produced by the bacterium. Also, the inoculants from different media elicit a differentiated nodulation on soybean plant.
Key words: Bradyrhizobium, nodulation, chromatography, legumes.
RESUMEN
Los factores de nodulación, sintetizados por diferentes especies de la familia Rizobiaceae, han sido descritos como señales esenciales en la interacción con plantas leguminosas. Ellos constituyen morfógenos que inducen el desarrollo de nódulos en la planta, permitiendo la entrada de las bacterias a las raíces y han demostrado tener una influencia positiva en el posterior desarrollo de los bacteroides y la eficiencia de la fijación del nitrógeno. Este trabajo está relacionado con la detección, mediante dos técnicas cromatográficas, de la producción de factores Nod por la cepa Bradyrhizobium elkanii ICA 8001, cultivada en diferentes medios; así como el efecto biológico de los inóculos obtenidos en interacción con la planta. Ambos métodos cromatográficos demostraron que la composición del medio de cultivo induce, en mayor o menor cantidad, el número de estructuras de factores Nod producidos por esta bacteria. Además, que inoculantes obtenidos a partir de diferentes composiciones de medio, inducen una nodulación diferenciada sobre plantas de soya.
Palabras clave: Bradyrhizobium, nodulación, cromatografía, leguminosas.
INTRODUCTION
It has been demonstrated that Nod factors induce several responses in the plant in a period from seconds to days (1), including deformation and curling of radical hairs, membrane depolarization, cortex cell division, new flavonoids formation, nodulin genes induction, the activation of calcium spiking, early nodulin (ENOD) gene induction, and infection thread initiation (2, 3, 4). Nod factors are certainly a key trigger for legume symbiotic signaling and nodule organogenesis (5).
Lipochitinoligosaccharids, characterized in Bradyrhizobium elkanii, varies in the size of their basic structure, in the type of fatty acid, and in some modifications of the basic structure, such as acetyl and carbamoil group or N-methyl on the non reducing terminal and 2-O-metylfucose or fucose and glycerol presence on the end reducer. Ten different structures of these molecules in Bradyrhizobium elkanii USDA 61 from several fractions obtained by high pressure liquid chromatography were identified (6). In 1996 (7) completed the chemical characterization studies of Nod factors produced by this strain, purifying the peaks with smaller absorbance. Similar structures were identified, but with singular combinations of the substitutes. This variability allows the potential formation of at least 96 different structures, from which 23 have already been identified, demonstrating the metabolic diversity of this species.
In this paper, we studied the Nod factors production by B. elkanii ICA 8001 strain in different culture media, using two chromatographic techniques and the biological effect of the inoculants on the interaction with soybean plants was validated.
MATERIAL AND METHODS
Microorganism, media and culture conditions
Bradyrhizobium elkanii ICA 8001 strain, from the Institute of Animal Sciences collection was used. The strain was cultivated in the different culture media in agitation conditions at 230 rpm and 30°C of temperature. The following culture media were used:Yeast Extract Manitol medium (YEM) (8), Propagation medium (9) and Modified Propagation mediumA.
In vitro nodulation assays
Experiments for nodulation studies were carried out in a totally randomized design, using 10 plants per treatment, under laboratory conditions. The data was processed by using the normality test (Bartlett´s test) and variance homogeneity (Kolmogorov-Smirnov´s test) and analysis of variance of simple classification was applied, using Duncan test of multiple ranges (p<0,05) to discriminate differences between media.
Soybean seeds, variety William 82, were used. They were disinfected in 70 % ethanol during 30 seconds and later in mercury dichloride 0,2 % (v/v) during 90 seconds. They were rinsed ten times with sterile distilled water and then allowed to rest for one hour in sterile distilled water to stimulate the germination. Ten seeds were placed in each Petri dish on water-agar; they were covered with aluminium foil and incubated during four days at 30°C. Ten dishes were used per experiment.
The germinated seeds, with approximately 20 mm of emergent roots, were placed in flasks of 250 mL containing 150 mL of Norris and date semisolid medium (10), one seed per flask. The plant roots were inoculated by discharging 200 µL of the inocula obtained with the strain B. japonicum ICA 8001 in the different culture media, in cellular concentration of 1,5 x 108 CFU.mL-1, corresponding to OD = 0,1 at 600 nm. A treatment of plants without inoculation was used as control in each assay. Plants were cultivated in a growth chamber with a photoperiod of 12 h light/12 h darkness, at a day/night temperature of 26/22ºC and relative humidity of 70 %.
Four weeks after the inoculation, the number of nodules per plant and the fresh and dry mass of the nodules were determined. Also the ability to fix atmospheric nitrogen was measured by means of the acetylene reduction assay, using a gas chromatograph (5890 A; Hewlett-Packard, equipped with a column of CHROMPACK "PLOT" (Porous Layer Open Tubular), 50 meters x 0,32 mm with Al2O3/KCl 5,0 µm to 60°C) as it is described in (11). The temperature of the oven was 130°C, the flow of N2 of 50 mL.min-1 and air and hydrogen was of 300 mL.min-1.
The plants were cut only leaving the radical system with the nodules. The flasks were sealed with rubber hermetic taps through which 10 mL of acetylene were injected. They were incubated during 15 minutes to measure the quantity of formed ethylene, then extracting 100 µL of gas and injecting it in the chromatograph. An internal standard of 100 µL of propane gas was used as a reference in each flask. The obtained values were expressed as ethylene micromoles produced by plant per hour (µmol C2H4. pl-1.h-1).
Detection of Nod factors
The Nod factors production were isolated, purified and identified following the methodology described by some authores (12). For the characterization by thin layer chromatography, Bradyrhizobium cultures were radioactively labelled adding 125 µL of 14C [2-14C] acetic acid as sodium salt and for high performance liquid chromatography determination; the cells were cultivated in 1 L of culture medium without isotopic label.
Determination of Nod factors production by thin layer chromatography (TLC): The Nod factor profile produced and excreted by B. japonicum ICA 8001 strain in YEM, propagation and modified propagation culture media was studied. Genistein 10 µM was also used as amplifier in all the extracts. The samples were applied on reverse-phase TLC plates (RP-18 F254 , Merck). H2O/acetonitrile (1:1, vol./vol.) was used as the mobile phase.
Determination of Nod factors production by normal phase high performance liquid chromatography (HPLC): The production of these molecules in YEM, propagation and modified propagation media was determined, as well as in the last one with genistein addition as amplifier at 10 µM concentration. The Nod factors purified were dissolved in 100 % acetonitrile and then injected into a normal phase column Ultropac TSK OH-120 5um LKB with dimensions of 4,6 x 250 mm. The flow rate 1 mL.min-1; as solvent were used: acetonitrile (A): water (B); the detector: an UV spectrophotometer at 206 nm and a cell of 10 mm, the Gradient in a Knauer pump: 0/0 10/0 70/20 t/%B and with an Injection of 250 µL in 100 % acetonitrile. The running time in all cases was 70 minutes. The chromatographic profile of the Nod factors was evaluated according to the number, distribution and relative intensity of the obtained peaks.
RESULTS AND DISCUSSION
Influence of different culture media on the soybean nodulation
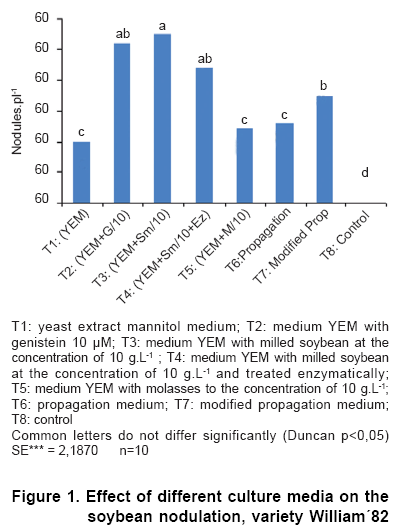
The evaluation of the influence of different culture media on the soybean nodulation was carried out on the base of previous results (Figures 1, 2, 3, 4). A new culture medium was designed from propagation medium, in which the milled soybean seed was incorporated up to the concentration of 10 g.L-1, the molasses concentration was cut down to 5 g.L-1 and the yeast extract was removed.
For the number of nodules (Figure 1), it was observed that the treatments presented significant differences. The best treatments were T2 (YEM + genistein), T3 (YEM + milled soybean) and T4 (YEM + soybean treated enzymatically), followed by T7 (modified propagation medium) that did not differ significantly from T2 and T4. The treatments T5: (YEM+molasses) and T6: (propagation) were inferior and statistically similar to the T1 (YEM) which only overcame the control without inoculation. The smallest number of nodules in the treatments T1 and T6 could be associated to the absence in them of the inducers behaved by the soybean.
The addition of milled soybean shows a similar behaviour to the genistein addition to the culture medium, reaching similar values of nodulation.
Figure 2 shows the influence of the treatments on the nodular fresh weight. The best results were reached in the treatments with genistein, milled soybean without enzymatic treatment and the modified propagation medium, which did not differ significantly. The enzymatic treatment to the soybean diminished its effect on this variable.
Regardless the milled soybean without enzymatic treated (T3), the other treatments neither differed from the T4 (YEM + milled soybean tried enzymatically) and T5 (YEM + molasse 10). The last one, were statistically similar, also, to the YEM and propagation media which only overcame to the absolute control.
Although the nodules developed by the inoculum in the modified propagation medium went inferior in number to those produced by the treatment with milled soybean (Figure 1), they showed a bigger size, what makes them statistically similar when analyzing the variable nodular fresh weight (Figure 2).
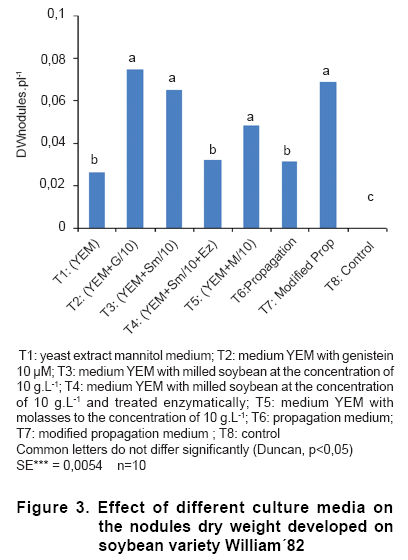
When the variable nodule dry weight was analyzed (Figure 3), the effects of the genistein addition were higher, along with the soybean without enzymatic treatment, the molasses and the modified propagation media. The treatments T4 (soybean treated enzymatically) and T6 (propagation medium) did not differ from the control YEM (T1) and they only surpass the control without inoculation. The enzymatic treatment affected negatively the nodular dry weight.
As for the activity of acetylene reduction (Figure 4), the inducers genistein, non treated soybean, molasses and the modified propagation medium, all showed superior results and statistically different to the rest of the treatments. Once again, the effect of the addition of soybean and/or molasses in the culture medium stands out, and is comparable to the action of genistein, and the capacity of the bacteria to fixing nitrogen, with regard to other culture media. Soybean enzymatic digestion before adding it to the culture medium influenced negatively the capacity of biological fixation, making it significantly different to the effect of the soybean without treatment and similar to YEM and propagation media.
Comparing the results with enzymatic digestion of soybean, it was verified that it didn't affect the number of nodules, while it significantly diminished the nodular weight and their efficiency. These results could be due to the hydrolysis was on those inducers or sensitive precursors to this type of enzymes, being degraded some components from soybean seed that participate in the establishment of the bacteroids, while other components of the seed that were not degraded, can also influence in the colonization and effectiveness of.
Comparing the influence of the treatments T5 (YEM with molasses) and T6 (propagation medium that contains molasses in same proportion) it was appreciated that on the number and nodular weight, the behaviour was very similar, but on the nitrogen fixation (ARA), YEM medium with molasses overcame the propagation. This indicates that not only the molasses have influence on, but also to keep in mind the role of the other components from the culture medium and the possible synergic or antagonistic effect that they can exert. The yeasts, for example, possess in their wall polymers of N-acetylglucosamine (13), those during the hydrolysis process for the extract obtainment, can be liberated as oligomers and so as they are contained in the culture medium, they could act as precursors of the nodulation factors synthesis in Bradyrhizobium. Then, increments in the production of certain structures of these biomolecules would lead to a bigger nodular efficiency. In the propagation medium, these compounds are in concentration five times higher than in YEM medium, what could produce inhibition for excess of this compound. An important influence was demonstrated by molasses on the activity of acetylene reduction, similar to that happened in previous experiment, where it didn't exercise an outstanding function as for the number neither nodules fresh weight, but in the dry weight of the same ones and in their fixation activity. This suggests that this compound influences less in the induction of the nodules formation, but more in the bacteroids establishment and in its function inside the nodule.
In general, the results suggest that the integration of the soybean seed milled and the molasses in the composition of modified propagation culture medium, led the bacteria to a physiologic state that was translated in significantly superior results of their biological activity on the soybean plant, in comparison with the traditional media.
Detection of the production of nodulation factors in the strain B. elkanii ICA 8001 by thin layer chromatography (TLC)
The profiles of the nodulation factors produced by this strain in presence of different compounds and culture media evaluated in the nodulation assay are shown in Figure 5. The strain exhibited a variety of profiles according to the inducer used for the activation of its nod genes.
YEM medium induced the production of 8 chromatographic spots. When genistein was used as inducer it showed 6 spots and those with smaller mobility according to TLC, stood out for its intensity.
The soybean sample without being enzymatically treated overcame in number and intensity the chromatographic spots of the soybean treated, the same way on some nodulation characters (Figures 2, 3 and 4). There were six spots in the raw soybean, all in the same position that those which appear in genistein, but this time the higher intensity corresponding to those of higher mobility.
The molasses, on the other hand, showed four compounds or group of compounds, most of them in great concentration and although only two in similar position to those of the soybean, coincidently, those of more intensity corresponded to the more mobility.
Similar to the results obtained in the nodulation assays, the soybean and the molasses constituted compounds that stood out in the Nod metabolites production with the synthesis of a great number of compounds, with the highest intensity.
The modified propagation medium overcame the Propagation medium with the production of eight chromatographic entities, some of them in high concentration. This medium causes the production of most of the Nod factors shown by the other inducers by separate, what is logical because its chemical composition, in which all of them are included.
Analyzing YEM and propagation media, it was observed that the first one overcame the latter one as for the production of these compounds; however, they did not show significant differences in the nodulation results.
The biggest production of Nod factors (evidenced in the TLC) in YEM medium was not translated in a bigger efficiency of the nodulation, when these results were compared with the nodulation in vitro assays, what suggests that not all the represented molecules had identical biological activity. It is known that not all the Nod factors produced and excreted, possess a significant biological activity, other rhizobial systems such as exopolysaccharide excretion (14), ethylene biosynthesis regulation (15) and protein secretion systems (16) are often required for the establishment of symbiosis with legumes, probably because they are involved in bacterial release into the host cytoplasm and bacteroid development.
The literature indicates the use of genistein as amplifier of the induction for Bradyrhizobium, constituting the best informed inducer for this genus. However, the results of this study demonstrated that their application to a concentration of 10 µM as amplifier of the synthesis of these biomolecules had a negative effect on 70 % of the assayed samples, what is the result of a production inhibition of these compounds. Only in those cases that did not show an appreciable production of Nod factors (soybean enzymatically digested and Propagation medium) the amplifier effect of genistein was present. The rest of the samples that revealed a clear synthesis of these biomolecules without the help of the genistein exhibited a repression in its production when this isoflavonoid was present. This is due; apparently, to an inhibition phenomenon for inducer excess (similar to what happens in the inhibition for substrate excess) exercised by the addition of the inducers contained in the culture medium plus the applied genistein. They have not been references of this phenomenon in the consulted literature, in which these inducers are always used in basal media, free of any other induction.
In a general way, the results of the Nod factors production for this strain were related directly with the nodulation response.
Detection of the production of nodulation factors in the strain B. japonicum ICA 8001 by means of high pressure liquid chromatography (HPLC) in normal phase
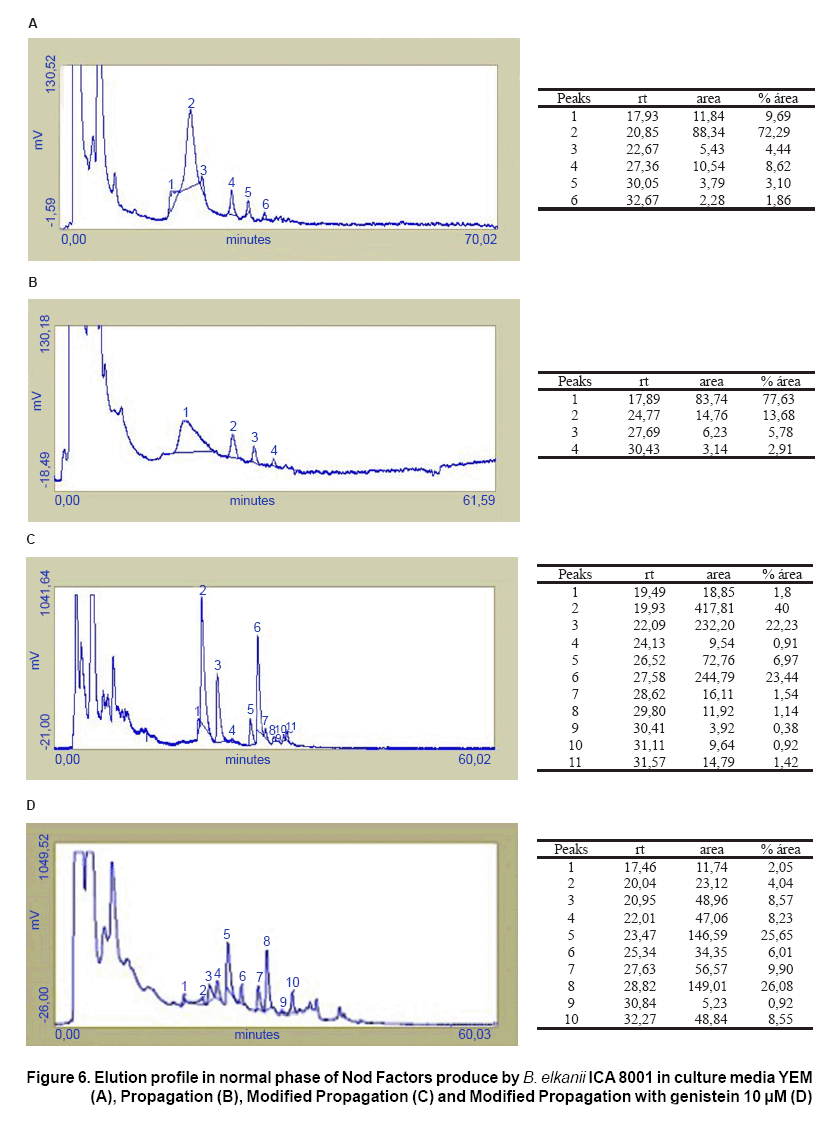
The analysis of the profiles obtained by means of liquid chromatography revealed marked differences between the number of chromatographic peaks and its relative intensity for the different culture media (Figure 6).
In all the profiles, less resolved peaks were presented in the smallest retention times (rt), which correspond to molecules more hydrophobic.
The smallest number of peaks was in the Propagation medium (Figure 6B), followed by YEM medium (Figure 6A). There is a small difference between these media concerning the distribution of peaks and the area under the curve, with four peaks in the propagation medium and six in YEM, in which three coincide to each other.
The biggest quantity in peaks and with more area under the curve was appreciated in the Modified Propagation medium (Figure 6C). In each one of the three majority peaks, the area is bigger that the total area of all the peaks in each one of the other samples. This indicates a higher production of Nod factors in this medium. When genistein was added to this medium composition, the depression of the chromatographic areas was observed, it mean that the production of these Nod factors was repressed, besides that a change in the distribution of the peaks happened (Figure 6D). Under these conditions, some peaks of the majority ones disappeared and one of them (rt.= 27,63 min) diminished to a third, while other (rt.= 28,8 min), increased ten times. Given the similarity in the retention time for both samples, they should correspond to similar structures of Nod factors present in them, although in front of the genistein they were presented in smaller concentration.
Different results were found by the other authors (17), where profile of these molecules with bigger number and intensity of peaks, characterized the samples induced with genistein and naringenine, respectively. The samples they studied, however, came from a basal medium that did not contain inducers, so the repression effect did not happen.
Although it is not possible to establish an exact relationship among the spots number obtained by TLC and the number of peaks found by means of HPLC, because the first method is much more specific, labelling radio-active and selectively the Nod factors, while HPLC is more potent to the separation of the products, the obtained results by the latter one, confirmed in general, those found with the thin layer chromatography.
By means of both techniques could be possible to verify the influence of the culture medium on the nodulation factors profiles. The tendency to the inhibition or repression of its synthesis in presence of an inducer excess in culture medium was also observed.
REFERENCES
1. Cooper, J. E. Early interactions between legumes and rhizobia: disclosing complexity in a molecular dialogue. Journal of Applied Microbiology, 2007, vol. 103, no. 5, p. 1355-1365.
2. Jones, K. M.; Kobayashi, K.; Davies, B. W.; Taga, M. E. and Walker, G. C. How rhizobial symbionts invade plants: the Sinorhizobium-Medicago model. Nature Reviews, 2007, vol. 5, p. 619-633.
3. Oldroyd, G. E. and Downie, J. A. Coordinating nodule morphogenesis with rhizobial infection in legumes. Annu. Rev. Plant Biol., 2008, vol. 59, p. 519-546.
4. Ito, S.; Ohtake, N.; Sueyoshi, K. y Ohyama, T. The autoregulation of nodulation mechanism is related to leaf development. Plant Cell Physiol., 2008, vol. 49, p. 121-125.
5. Kouchi, H.; Imaizumi-Anraku, H.; Hayashi, M.; Hakoyama, T.; Nakagawa, T.; Umehara, Y.; Suganuma, N. and Kawaguchi, M. How many peas in a pod? Legume genes responsible for mutualistic symbioses underground. Plant Cell Physiol., 2010, vol. 51, p. 1381-1397.
6. Carlson, R. W.; Sanjuán, J.; Bhat, U. R.; Glushka, J.; Spaink, H. P.; Wijfjes, H. W.; van Brussel, A. N.; Stokkermans, T. J. W.; Peters, N. K. and Stacey, G. The structures and biological activities of the lipo-oligosaccharide nodulation signals produced by Type I and Type II strains of Bradyrhizobium japonicum. J. Biol. Chem., 1993, vol. 268, p. 18372-18381.
7. Stokkermans, T. J. W.; Ron, O.; Kumar, V. S.; Rusell, C. W.; and Peters, N. K. Biological activities and structures of Bradyrhizobium elkanii low abundance lipo-chitin-oligosaccharides. Mol Plant-Microbe Interact., 1996, vol. 9, p. 298-304.
8. Vincent, J. M. A manual for the practical study of root-nodule bacteria/J.M. Vincent. En: International Biological Programme Handbook. No. 15. Blackwele scientific publications, Oxford, England. 1970.
9. López, M. Manual de Tecnologías de Producción del biofertilizante Bio-Rhizo. La Habana: Empresa Cubana de Productos Veterinarios Cuba-Vet. Instituto de Ciencia Animal, Instituto de Investigaciones Pastos y Forrajes. 1990.
10. Norris, D. O. and Date, R. A. Legume bacteriology Tropical Pasteur Research. Principles and Methods. C.A.B. Bill., 1976, vol. 51, p.134-174.
11. Dombrecht, B. The complex regulation and role of the Rhizobium etli Rpon regulon in the symbiotic interaction with the common bean plant (Phaseolus vulgaris L.). Dissertationes de Agricultura, December 2001, 151 p.
12. Laeremans, T. The role of the Nod factor sulphation genes in the Rhizobium-Phaseolus vulgaris (L.) interaction. Dissertationes de Agricultura, April 1998. 142 p.
13. Konopka, J. N-Acetylglucosamine Functions in Cell Signaling. Scientifica., 2012, 15 p. doi.org/10.6064/2012/489208.
14. Downie, J. A. The roles of extracellular proteins, polysaccharides and signals in the interactions of rhizobia with legume roots. FEMS Microbiol. Rev., 2010, vol. 34, p. 150-170.
15. Gresshoff, P. M.; Lohar, D.; Chan, P.; Biswas, B.; Jiang, Q. and Reid, D. Genetic analysis of ethylene regulation of legume nodulation. Plant Signal, 2009, vol. 4, p. 818-823.
16. Deakin, W. J. and Broughton, W. J. Symbiotic use of pathogenic strategies: rhizobial protein secretion systems. Nat. Rev. Microbiol., 2009, vol. 7312-320.
17. Cárdenas, L.; Domínguez, J.; Quinto, C.; López-Lara, I.; Lugtenberg, B.; Spaink, H.; Rademaker, G.; Haverkamp, J. and Thomas-Oates, J. Isolation, chemical structures and biological activity of lipo-chitin oligosaccharide nodulation signals from Rhizobium etli. Plant Molecular Biology, 1995, vol. 29, p. 453-464.
Recibido: 2 de septiembre de 2011
Aceptado: 4 de marzo de 2013
Dra.C. María C. Nápoles García, Investigador Titular del departamento de Fisiología y Bioquímica, Instituto Nacional de Ciencias Agrícolas, gaveta postal 1, San José de las Lajas, Mayabeque, Cuba. Email: tere@inca.edu.cu


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}