










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCultivos Tropicales
versión impresa ISSN 0258-5936
cultrop vol.35 no.2 La Habana abr.--jun. 2014
Clasificación taxonómica de la cepa de hongo micorrizógeno arbuscular INCAM-2 como Funneliformis mosseae, syn. Glomus mosseae
Taxonomic classification of the arbuscular mycorrhizal fungal strain INCAM-2 as Funneliformis mosseae, syn. Glomus mosseae
Dra.C. Yakelin Rodríguez,I Dr.C. Yolande Dalpé,II M.Sc. Sylvie SéguinIII
IInvestigadora Auxiliar del departamento Biofertilizantes y Nutrición de las Plantas, Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, Mayabeque, CP 32 700, Cuba.
IIInvestigadora y Profesora Adjunta de la Universidad de Montreal, Eastern Cereal and Oilseed Research Centre, Agriculture and Agri-Food Canada, 960 Carling Ave, Ottawa K1A 0C6, Canadá.
IIIEspecialista del laboratorio de la Universidad de Montreal, Eastern Cereal and Oilseed Research Centre, Agriculture and Agri-Food Canada, 960 Carling Ave, Ottawa K1A 0C6, Canadá.
RESUMEN
Dentro de la colección de cepas de hongos micorrizógenos arbusculares (HMA) del INCA se destaca por su importancia económica y ecológica la cepa INCAM-2, al constituir la base de biopreparados que se aplican en diversos sistemas agrícolas y se comercializan ampliamente a nivel mundial. El objetivo de este trabajo fue confirmar la clasificación taxonómica preliminar realizada de dicha cepa, mediante taxonomía polifásica. Para ello se analizaron con mayor profundidad las características morfológicas de las esporas y se utilizaron otros protocolos moleculares (ITS del ADN ribosomal y V-H+-ATPasa). Los resultados demostraron que la cepa INCAM-2 es Funneliformis mosseae, syn. Glomus mosseae. Las esporas se distinguen por su gran tamaño, su coloración de amarillo oscuro a ámbar, la pared consta de tres capas (C1 hialina mucilaginosa, C2 hialina y C3 laminada), donde C1 se tiñe de rojizo con la solución de Melzer, la unión de la hifa de sostenimiento a la espora es en forma de embudo, presenta un septo curvo y el poro está ocluido. El análisis filogenético de las secuencias obtenidas de ambos genes reveló que presentan un 98 % de homología con varios ecotipos de la misma especie registrados en las bases de datos del Centro Nacional de Información Biotecnológica de los Estados Unidos (NCBI).
Palabras clave: glomerales, caracterización molecular, identificación taxonómica, taxonomía polifásica.
ABSTRACT
The strain INCAM-2 outstands among arbuscular mycorrhizal fungal strains from the INCA´s collection due to its economic and ecological significance. It constitutes the base of bioproducts which are applied in different agricultural systems and are extensively commercialized at world level. This work aimed in confirms the taxonomic classification previously done of this strain through polyphasic taxonomy. For this, the spore morphologic characteristics with major depth were analyzed and other molecular protocols were used (rDNA ITS region and V-H+-ATPasa). The results showed that INCAM-2 strain is Funneliformis mosseae, syn. Glomus mosseae. The spores distinguish by their big size, color dark yellow to amber, the wall composed by three layers (C1 hyaline mucilaginous, C2 hyaline and C3 laminated). C1 stains reddish with Melzer´s reagent. Subtending hypha has a funnel shape and forms a recurved septum, the pore is occluded. The phylogenetic analysis of obtained sequences of both genes reveled a 98 % of homology with some ecotypes of the same species registered in the data bases of National Center of Biotechnologic Information of the United States of America (NCBI).
Key words: glomerales, molecular characterization, taxonomic identification, polyphasic taxonomy.
INTRODUCCIÓN
La mayoría de las especies de HMA descritas hasta el momento han sido caracterizadas sólo por la morfología de sus esporas debido a que muchas de ellas no han podido ser cultivadas en macetas en cultivos puros, en asociación con plantas hospederas, especialmente las que requieren nichos específicos (1, 2). Sin embargo, el criterio basado solamente en su morfología no refleja la gran plasticidad fisiológica y diversidad genética de las poblaciones micorrízicas (3).
La aplicación de técnicas moleculares ha permitido un avance importante en los estudios concernientes a la identificación de estos hongos (4, 5).
En este sentido, una de las contribuciones más notables lo constituyó la separación de los mismos en un phylum independiente: Glomeromycota (6). Por esto la tendencia actual en estudios taxonómicos es la aplicación combinada de los métodos convencionales, en este caso morfológicos, con los moleculares, lo cual se conoce como taxonomía polifásica (7).
Por otro lado, la cepa de HMA INCAM-2, perteneciente a la colección del INCA, se considera una de las más promisorias dados los efectos beneficiosos que se observan en las plantas con su inoculación. Entre estos se destaca el aumento de la tolerancia al déficit hídrico (8), así como el incremento del rendimiento y un mejor estado nutritivo del tomate en condiciones de suelo Ferralítico Rojo Lixiviado (9) y el favorecimiento de la aclimatización de vitroplantas de papa (10).
En estudios previos se demostró que la cepa INCAM-2 es Glomus mosseae (11). Este resultado se basó en la caracterización morfológica de las esporas y en la aplicación de una técnica molecular sencilla (12). En la misma se realizaron reacciones en cadena de la polimerasa anidada (NESTED PCR), donde en la segunda PCR se utilizó un cebador taxón-específico discriminante para Glomus mosseae, conocido como 5,25 (12). Estos autores emplearon como marcador la subunidad ribosomal mayor (LSU).
Asimismo se han utilizado con frecuencia para estos fines los genes ribosómicos nucleares ITS y SSU (13, 14). No obstante, en la actualidad, continuan las investigaciones encaminadas al desarrollo de nuevos protocolos moleculares menos engorrosos y que permitan la identificación certera de un mayor número de especies de HMA. Entre estos resaltan los que se basan en genes que codifican para proteínas, como es el caso de la tubulina (15) y la V-H+-ATPasa vacuolar (16).
El objetivo de este trabajo fue confirmar la clasificación taxonómica de la cepa INCAM-2 como Glomus mosseae, mediante el análisis detallado de la morfología de las esporas y el empleo de otros dos protocolos moleculares.
MATERIALES Y METODOS
Aislamiento y selección de las esporas de HMA
Se analizaron esporas de la cepa de HMA perteneciente a la colección del INCA, INCAM-2 (Glomus mosseae (Nicol. & Gerd.) Gerd. & Trappe (17, 18)), que fueron extraídas de cultivos puros sobre Sorghum bicolor L. (19), mediante la técnica de Tamizado húmedo (300 y 38 µm) y centrifugación en gradiente de sacarosa (20), procediéndose al aislamiento manual de esporas jóvenes homogéneas según su apariencia al microscopio estéreo 70x (Olympus SZ-PT), con auxilio de micropipeta.
Caracterización morfológica
Se examinaron 100 esporas para analizar sus propiedades morfológicas: color, forma y tamaño de las esporas; número, estructura, grosor y respuesta a distintas tinciones de las capas de la pared, según terminología propuesta por Walker (21) y Stürmer y Morton (22); características de la hifa de sostenimiento: forma, número de capas de la pared y grosor, poro. Para ello se realizaron preparaciones permanentes con diferentes soluciones de montaje: alcohol polivinílico-ácido láctico-glicerol (PVLG) (23), PVLG con solución de Melzer (1:1, v/v) y PVLG con azul de algodón (0,15 % en ácido láctico) (1:1, v/v). La clasificación de colores se realizó según el código de la carta de colores de la colección internacional del cultivo de hongos micorrizógenos arbusculares y Vesículo-Arbusculares (24). Se utilizó un microscopio compuesto Nikon Eclipse 800, equipado con contraste de interferencia diferencial Nomarski y las fotografías se tomaron con una cámara digital Nikon CoolPix
4 500, lo cual posibilitó ahondar más en la morfología de las esporas.
Caracterización molecular
Extracción de ADN. Las esporas se lavaron con agua destilada y se secaron al aire. La extracción de ADN se realizó con la utilización de micro-pistilo plástico, a partir de 26 esporas a las cuales se les añadieron 25 µL de la solución amortiguadora de extracción del juego de reactivos PreManUltra (Applied Biosystems). Las esporas maceradas se incubaron a 90°C por 10 min, seguido por incubación a temperatura ambiente por dos minutos y centrifugación a 15 000 rpm (International Equipment Company Micromax) por cinco minutos. El sobrenadante se colectó y diluyó (1:10) en Tris-HCl 10 mmol.L-1 (pH 8).
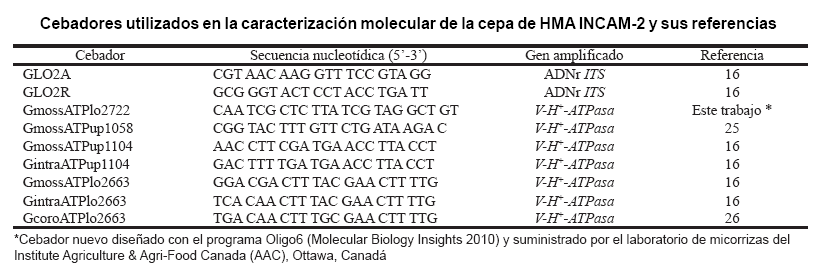
Todas las amplificaciones se realizaron en termociclador (Biometra TProfessional). Las polimerasas y los cebadores fueron proveídos por Invitrogen. La información referente a los cebadores utilizados aparece en la tabla.
Análisis de ITS del ADN ribosomal. Se realizó una amplificación cuya mezcla de reacción contenía 1x de solución amortiguadora Fermentas Taq (20 mmol.L-1 Tris (pH 8,4) y 50 mmol.L-1 KCl), 0,25 mmol.L-1 de cada dNTP, 1 mmol.L-1 de cada cebador (GLO2A y GLO2R), 0,4 unidades de Fermentas Taq polimerasa, 3 mmol.L-1 de MgCl2 y 1 µL de ADN genómico 1-5 ng.µL-1, en un volumen final de 10 µL (16). Se ejecutó el programa: 3 min a 94°C, 38 ciclos de 30 seg a 94°C, 60 seg a 50°C, 90 seg a 72°C, con un ciclo final de 7 min a 72°C (16). Los productos de la PCR se clonaron con el uso del juego de reactivos TOPO TA Cloning® (Invitrogen), empleando el vector pCR®4-TOPO® y la cepa de Escherichia coli (TOP10). La selección de las colonias se realizó con kanamicina primero y con ampicilina después. Luego se procedió a determinar y crecer las colonias con el inserto apropiado (650 pb), purificándose los plásmidos con el juego de reactivos GenEluteTM Plasmid Miniprep (Sigma).
Los dos protocolos se desarrollaron según instrucciones del fabricante para ADNr ITS.
Análisis de V-H+-ATPasa. Se utilizó un cebador nuevo diseñado y proveído por el laboratorio de micorrizas del Instituto AAC: GmossATPlo2722, el cual fue diseñado a partir de la secuencia del NCBI, AY193825. Se realizaron tres PCRs utilizando las siguientes combinaciones de cebadores: GmossATPup1058–GmossATPlo2722; GmossATPup1104, GintraATPup1104–GmossATPlo2722 y GmossATPup1058–GimcATPlo2663 (mezcla equimolar de GmossATPlo2663, GintraATPlo2663 y GcoroATPlo2663), tal como se aprecia en la tabla.
La mezcla de reacción de las PCRs contenía 1x de solución amortiguadora Titanium Taq (3,5 mmol.L-1 MgCl2), 0,1 mmol.L-1 de cada dNTP, 0,08 µmol.L-1 de cada cebador; 0,5x Titanium Taq polimerasa y 1 µL de ADN genómico 1-5 ng.µL-1, en un volumen final de 10 µL (16). Las amplificaciones se realizaron de la siguiente forma: 3 min a 94°C, a continuación 40 ciclos de 30 seg a 94°C, 45 seg a 58°C, 2 min a 72°C y el último ciclo fue seguido por una extensión final de 7 min a 72°C (16).
Los productos de las PCR, en ambos casos, fueron analizados mediante electroforesis en geles de agarosa (1,5 %) en TAE que contenían bromuro de etidio (0,05 µL.mL-1) y se usó solución amortiguadora de corrida TAE 1x (Tris 40 mmol.L-1 (pH 7,8), ácido acético y EDTA). Se añadió 1 µL del marcador de peso molecular 1Kb Plus DNA Ladder (Invitrogen) 1 µg.µL-1. Las corridas se realizaron a 60 voltios por 60 minutos en equipo de electroforesis horizontal (BIORAD). Los fragmentos de ADN fueron visualizados y fotografiados bajo luz ultravioleta.
Análisis de las secuencias. La secuenciación se efectuó con el juego de reactivos “Cycle Sequencing BigDye Terminator” 3.1, en un secuenciador automático ABI 3100-Avant (Applied Biosystems/Hitachi), en el laboratorio de Micología de AAC. Las secuencias obtenidas se editaron manualmente en SeqMan (27) y se compararon con las bases de datos del NCBI a través del BLAST. Se seleccionó el modelo de trabajo en cada caso. El valor del parámetro Gamma y el logaritmo de verosimilitud se estimaron por el método estadístico de máxima verosimilitud. Se utilizó el programa MEGA 5,0 (28) para alinear las secuencias seleccionadas y construir los árboles filogenéticos correspondientes mediante el método de distancia del vecino más cercano. Se empleó el método de bootstrap con 1000 repeticiones como prueba de filogenia. En los árboles filogenéticos se empleó la nomenclatura y clasificación de los HMA propuesta por Schüßler y Walker (29).
RESULTADOS Y DISCUSIÓN
Caracterización morfológica
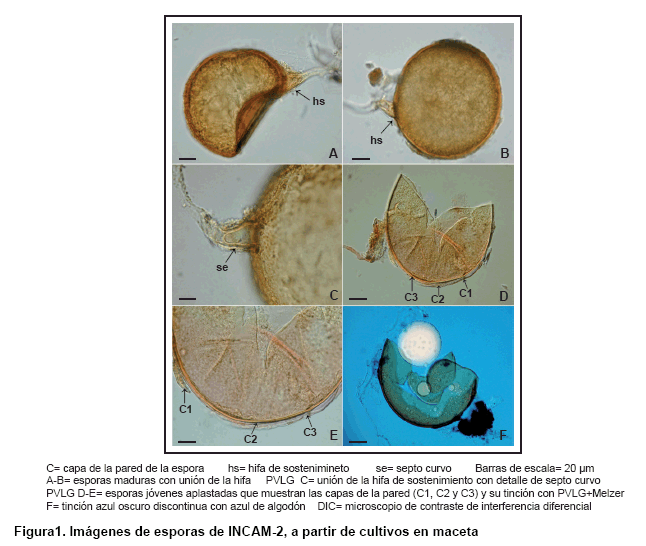
Las esporas de la cepa INCAM-2 son de color amarillo a ámbar (0/30/100/0)-(0/20/80/0), poseen forma globosa o subglobosa y tamaño entre 100-250 µm (Figura 1A-B). La pared de las esporas está compuesta por tres capas (Figura 1D-E), la C1 es hialina, mucilaginosa, aparece frecuentemente degradada en esporas maduras, de espesor entre 1,5-2,5 µm. C2 es hialina, rígida, con espesor de 0,8-1,6 µm. C3 es de color amarillo, laminada y presenta un espesor de 3,2-6,4 µm. La reacción con la solución de Melzer es positiva (C1-rojizo; C3-amarillo claro). C1 se tiñe de azul oscuro en forma discontinua o de parches con azul de algodón (Figura 1F). La hifa de sostenimiento posee el mismo color de las esporas, presenta forma de embudo, es en su mayoría curva, el ancho en la base de la espora es 21-30 µm (Figura 1A-C). La pared de la hifa de sostenimiento es del mismo color de la pared de la espora (hialino y amarillo), el espesor en la base de la espora es entre 6-7,5 µm, está compuesta por dos capas continuas con C1 y C3 de la espora (Figura 1A-C). El poro está cerrado al presentar un septo curvo localizado de 0-32 µm de la base de la espora (Figura 1C). Las esporas se encuentran en forma simple en el suelo (Figura 1A-B).
Al comparar las características morfológicas de las cepas analizadas con las que aparecen descritas en el catálogo INVAM (24) se confirmó que INCAM-2 es Funneliformis mosseae (Nicol. & Gerd.) Walker & Schüßler (29), syn. Glomus mosseae.
No obstante, en la observación al microscopio óptico la especie F. coronatum (Giovann.) Walker & Schüßler (29, 30) muestra elevada similitud a F. mosseae, con la salvedad de la coloración ámbar más oscura de sus esporas, la unión de la hifa de sostenimiento a la espora en forma de embudo más marcada en la mayoría de la población, y que su pared tiene sólo dos capas, aunque en ocasiones es difícil la distinción debido a que en esporas maduras de F. mosseae la C1 generalmente aparece degradada. Las esporas de F. caledonium (Nicol. & Gerd.) Walker & Schüßler (17, 29) también se asemejan, pero esta especie tiene una pared conformada por cuatro capas con estructuras y fenotipos distintivos que la diferencian de F. mosseae. Además, la unión de la hifa de sostenimiento a la espora es cilíndrica y en ocasiones ligeramente constreñida.
Es válido destacar que en el presente trabajo, a diferencia del realizado por otros autores (11), dentro de las variables morfológicas se analizó con mayor detalle la estructura de las capas de la pared de las esporas y de la hifa de sostenimiento, particularizando en el grosor de las mismas. En general, se estudiaron todas las características morfológicas de la hifa de sostenimiento de las esporas de la cepa INCAM-2, las que no habían sido evaluadas con anterioridad. Se determinó la respuesta a la tinción con azul de algodón. Además, para el color se indicó el código según la carta de colores de la colección internacional del cultivo de hongos micorrizógenos arbusculares y vesículo-arbusculares, lo cual facilita la comparación con esporas de especies similares.
Caracterización molecular
El análisis filogenético de las secuencias obtenidas de ambos genes (Figura 2) sustenta la caracterización morfológica de esta cepa y corrobora que, en efecto, INCAM-2 es Funneliformis mosseae, syn. Glomus mosseae, al presentar dichas secuencias un 98 % de homología con varios ecotipos de la misma especie registrados en las bases de datos del NCBI, para los dos genes analizados.
Además, la comparación de las secuencias de ITS mostró un 90 y un 88 % de similitud con las de F. caledonium y F. coronatum, respectivamente, y la de V-H+-ATPasa, un 93 % de homología con las de F. caledonium y G. multiforum Tadych & Blaszkowski.
La secuencia resultante de V-H+-ATPasa posee 1641 pb, mientras que las de ITS presentan 524 (tres clones), 521 y 517 pb. Las mismas fueron inscritas en el NCBI bajo los números de acceso JQ178252 para V-H+-ATPasa, y para los cinco clones analizados del ADNr ITS fue del JQ178247 a JQ178251 (Figura 2).
Lo expuesto evidencia la importancia de realizar la caracterización molecular para constatar la identidad de las cepas de HMA, con lo cual se integran los criterios morfológico y molecular (taxonomía polifásica). El análisis molecular realizado demostró la efectividad de la aplicación conjunta de dos marcadores que se complementan, en este caso ITS y V-H+-ATPasa, para identificar estos hongos. Esto coincide con trabajos recientes donde se combinaron los ITS y SSU (31, 32, 33, 34, 35), lo que resulta pertinente cuando uno de los marcadores no separa la especie descrita al nivel requerido. Asimismo, se han combinado los tres genes ribosómicos nucleares (SSU, ITS y LSU), estableciendo como un código de barra, para discernir entre especies muy cercanas filogenética y morfológicamente (13, 14).
Entre los genes ribosómicos nucleares, el ITS resuelve gran parte de las especies conocidas del género Glomus (14). Esto justifica su selección en este estudio, demostrando ser eficaz, lo que concuerda con trabajos recientes (32, 33, 34,), donde se describen nuevas especies de HMA a partir de este marcador. Aunque no se debe obviar la desventaja de su gran variabilidad intra-específica e intra-espora para estos hongos en particular, lo cual implica necesariamente realizar el protocolo de la clonación que es largo y muy engorroso en términos de manipulación, dificultando el procedimiento de identificación. Los cebadores de ITS específicos de Glomus utilizados aquí: GLO2A-GLO2R (16), fueron ensayados anteriormente con éxito en la identificación de dos especies nuevas (23, 34), lo cual en conjunto con los resultados de este trabajo avalan su utilidad para este género.
Por su parte, el marcador V-H+-ATPasa ofrece la ventaja de presentar una sola secuencia para cada especie (26), indicando que no es necesario clonar, lo cual explica su selección en este trabajo y demuestra que su implementación en la identificación molecular de los HMA contribuye a disminuir los costos, reducir el tiempo y a simplificar el proceso. Este marcador ya había sido utilizado en la taxonomía de los HMA con resultados satisfactorios; sin embargo, se considera novedoso para este fin pues solo ha sido implementado en dos estudios recientes (16, 23), donde en uno de ellos demostró ser eficaz en la identificación de una nueva especie: Glomus cubense (23).
El cebador nuevo de V-H+-ATPasa informado en este estudio resultó útil, al menos, para la especie F. mosseae, aunque no se descarta su funcionalidad para otras especies de la familia Glomeraceae. Además, permitió amplificar un fragmento del gen de talla mayor (1641 pb) respecto al amplicón de 1468 pb que reveló la previa secuenciación de este gen (26), por lo que se recomienda su aplicación en la identificación de otras especies.
Resulta útil disponer de un marcador de fácil aplicación, como es el V-H+-ATPasa que permita discriminar entre especies cercanas de Glomerales, si se considera que este orden constituye el de mayor representación en los ecosistemas y, en la mayoría de los casos, las diferencias morfológicas entre algunas de las especies son muy sutiles (13, 14, 36). Esto valida la ventaja de su uso, en comparación con los marcadores ribosómicos, por lo que es conveniente continuar trabajando en el diseño de nuevos cebadores que funcionen en un mayor número de especies.
La estrategia desarrollada para la secuenciación de los fragmentos de V-H+ -ATPasa sientan las bases para el diseño de cebadores específicos tanto de la cepa estudiada como de otras, los cuales pudieran ser empleados en el monitoreo y en la identificación de las mismas en los agrosistemas donde se aplican los productos a base de estas cepas.
REFERENCIAS
1. Oehl, F.; Sieverding, E.; Ineichen, K.; Mäder, P.; Wiemken, A. y Boller, T. Distinct sporulation dynamics of arbuscular mycorrhizal fungal communities from different agroecosystems in long-term microcosms. Agr. Ecosyst. Environ., 2009, vol. 134, p. 257-268.
2. Palenzuela, J.; Barea, J. M.; Ferrol, N.; Azcón-Aguilar, C. y Oehl, F. Entrophospora nevadensis, a new arbuscular mycorrhizal fungus, from Sierra Nevada National Park (southeastern Spain). Mycologia, 2010, vol. 102, p. 624-632.
3. Redecker, D.; Morton, J. B. y Bruns, T. D. Ancestral lineages of arbuscular mycorrhizal fungi (Glomales). Mol. Phylogenet. Evol., 2000, vol. 14, p. 276-284.
4. Blaszkowski, J.; Kovács, G. M. y Balázs, T. Glomus perpusillum, a new arbuscular mycorrhizal fungus. Mycologia, 2009, vol. 101, no. 2, p. 247-255.
5. Blaszkowski, J.; Kovács, G. M.; Balázs, T.; Orlowska, E.; Sadravi, M. y Wubet, T. Glomus africanum and G. iranicum, two new species of arbuscular mycorrhizal fungi (Glomeromycota). Mycologia, 2010, vol. 102, no. 6, p. 1450-1462.
6. Schüßler, A.; Schwarzott, D. y Christopher, W. A new fungal phylum, the Glomeromycota: phylogeny and evolution. Mycol Res., 2001, vol. 105, no. 12, p. 1413-1421.
7. Tindall, B. J.; Rosselló-Móra, R.; Busse, H. J.; Ludwig, W. y Kampfer, P. Notes on the characterization of prokaryote strains for taxonomic purposes. International Journal of Systematic and Evolutionary Microbiology, 2010, vol. 60, p. 249-266.
8. Fundora, L. R.; Rodríguez, Y.; Mena, A.; González, P.; Rodríguez, P. y González-Peña, D. Estabilidad de la eficiencia de la cepa Glomus mosseae en la respuesta del tomate a condiciones de estrés hídrico fuera de su periodo óptimo. Cultivos Tropicales, 2008, vol. 29, no. 2, p. 47-53.
9. Mujica, Y. y Medina, N. Respuesta del tomate (Solanum lycopersicum L.) a la formulación líquida de cuatro cepas de Glomus en condiciones de campo. Cultivos Tropicales, 2008, vol. 29, no. 3, p. 23-25.
10. Rodríguez, Y.; Mena, A.; Marentes, F. L. y Fernández, K. Respuesta de enzimas antioxidantes y crecimiento de vitroplantas de papa micorrizadas in vitro. Cultivos Tropicales, 2008, vol. 29, no. 1, p. 29-35.
11. Rodríguez, Y.; Tuinen, D. van y Fernández, K. Reclasificación taxonómica de dos cepas de hongos micorrízicos arbusculares. Cultivos Tropicales, 2009, vol. 30, no. 1, p. 31-35.
12. Tuinen, D. van; Zhao, B. y Gianinazzi-Pearson, V. PCR in studies of AM fungi: from primers to application. En: Mycorrhizal manual. New York. Ed. por Varma, A. K. Springer, Berlin Heidelberg 1998, p. 387-399.
13. Stockinger, H.; Walker, Ch. y Schüßler, A. ‘Glomus intraradices DAOM197198’, a model fungus in arbuscular mycorrhiza research, is not Glomus intraradices. New Phytol., 2009, vol. 183, p. 1176-1187.
14. Stockinger, H.; Krüger, M. y Schüßler, A. DNA barcoding of arbuscular mycorrhizal fungi. New Phytol., 2010, vol. 187, p. 461-474.
15. Msiska, Z. y Morton, J. B. Phylogenetic analysis of the Glomeromycota by partial ß-tubulin gene sequences. Mycorrhiza, 2009, vol. 19, p. 247-254.
16. Sokolski, S.; Dalpé, Y.; Séguin, S.; Khasa, D.; Lévesque, C. A. y Piché, Y. Conspecificity of DAOM 197198, the model AM fungus, with Glomus irregulare: molecular evidence with three protein-encoding genes. Botany, 2010, vol. 88, p. 829-838.
17. Nicolson, T. H. y Gerdemann, J. W. Mycorrhizal Endogone Species. Mycologia, 1968, vol. 60, no. 2, p. 313-325.
18. Gerdemann, J. W. y Trappe, J. M. Endogonaceae in the Pacific Northwest. Mycologia Memoir., 1974, vol. 5, p. 1-76.
19. Instituto Nacional de Ciencias Agrícolas (INCA). Producto inoculante micorrizógeno. Inventor: Fernández Martín, F. M. A01N 63/00, A01N 63/04. Cuba, Oficina Cubana de la Propiedad Intelectual (OCPI). Patente de Invención: CU 22641. 2001. 2000-5-2.
20. Dalpé, Y. y Hamel, C. Arbuscular mycorrhizae. En: Manual of Soil Sampling and Methods of Analysis. 4rd Edition, Canadian Society of Soil Science Lewis Publishers of CRC Press, 2007, p. 355-377.
21. Walker, C. Taxonomic concepts in the Endogonaceae: Spore wall characteristics in species descriptions. Mycotaxon, 1983, vol. 18, p. 443-455.
22. Stürmer, S. L. y Morton, J. B. Developmental patterns defining morphological characters in spores of four species in Glomus. Mycologia, 1997, vol. 89, p. 72-81.
23. Omar, M. B.; Bolland, L. y Heather, W. A. A permanent mounting medium for fungi. Bulletin of the British Mycological Society, 1979, vol. 13, p. 31.
24. INVAM. Carta de colores de la Colección Internacional del Cultivo de Hongos Micorrizógenos Arbusculares y Vesículo-Arbusculares. [Consultado: 25 de febrero del 2012]. Disponible en: <http://www.invam.caf.wvu.edu>.
25. Rodríguez, Y.; Dalpé, Y.; Séguin, S.; Fernández, K.; Fernández, F. y Rivera, R. A. Glomus cubense sp. nov., an arbuscular mycorrhizal fungus from Cuba. Mycotaxon, 2011, vol. 118, p. 337-347.
26. Corradi, N.; Kuhn, G. y Sanders, I. R. Monophyly of ß-tubulin and H+-ATPase gene variants in Glomus intraradices: consequences for molecular evolutionary studies of AM fungal genes. Fungal Genet Biol., 2004, vol. 41, p. 262-273.
27. DNASTAR. Software utilizado para la edición de secuencias de ADN. Consultado: enero-marzo del 2012]. Disponible en: <http://www.dnastar.com>.
28. Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M. y Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Molecular Biology and Evolution, 2011.
29. Shüßler, A. y Walker, C. The Glomeromycota: a species list with new families and new genera. Gloucester: The Royal Botanic Garden Edinburgh, The Royal Botanic Garden Kew, Botanische Staatssammlung Munich and Oregon State University, 2010, 58 p.
30. Giovannetti, M.; Avio, L. y Salutini, L. Morphological, cytochemical, and ontogenetic characteristics of a new species of vesicular-arbuscular mycorrhizal fungus. Can. J. Bot., 1991, vol. 69, p. 161-167.
31. Blaszkowski, J.; Czerniawska, B.; Wubet, T.; Schäfer, T.; Buscot, F. y Renker, C. Glomus irregulare, a new arbuscular mycorrhizal fungus in the Glomeromycota. Mycotaxon, 2008, vol. 106, p. 247-267.
32. Blaszkowski, J.; Ryszka, P.; Oehl, F.; Koegel, S.; Wiemken, A.; Kovács, G. M. y Redecker, D. Glomus achrum and G. bistratum, two new species of arbuscular mycorrhizal fungi (Glomeromycota) found in maritime sand dunes. Botany, 2009, vol. 87, no. 3, p. 260-271.
33. Blaszkowski, J.; Wubet, T.; Harikumar, V. S.; Ryszka, P. y Buscot, F. Glomus indicum, a new arbuscular mycorrhizal fungus. Botany, 2010, vol. 88, p. 132-143.
34. Cano, C.; Bago, A. y Dalpé, Y. Glomus custos sp. nov., isolated from a naturally heavy metal-polluted environment in southern Spain. Mycotaxon, 2009, vol. 109, p. 499-512.
35. Palenzuela, J.; Barea, J. M.; Ferrol, N. y Oehl, F. Ambispora granatensis, a new arbuscular mycorrhizal fungus, associated with Asparagus officinalis in Andalucía (Spain). Mycologia, 2011, vol. 103, no. 2, p. 333-340.
36. Öpik, M.; Moora, M.; Zobel, M.; Saks, Ü.; Wheatley, R.; Wright, F. y Daniel, T. High diversity of arbuscular mycorrhizal fungi in a boreal herb-rich coniferous forest. New Phytol., 2008, vol. 179, p. 867-876.
Recibido: 10 de abril de 2012
Aceptado: 6 de mayo de 2013
Dra.C. Yakelin Rodríguez, Investigadora Auxiliar del departamento Biofertilizantes y Nutrición de las Plantas, Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, Mayabeque, CP 32 700, Cuba. Email: yakelin@inca.edu.cu

{kind=link}
{kind=link}
{kind=link}