



![Caracterización del desarrollo vegetativo y su relación con la fructificación y producción en naranjos [Citrus sinensis (L.) Osbeck]](/img/es/prev.gif)






Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCultivos Tropicales
versión impresa ISSN 0258-5936versión On-line ISSN 1819-4087
cultrop vol.36 no.2 La Habana abr.-jun. 2015
Efecto de la calidad de la luz en el cultivo del maíz: una herramienta para el manejo de plantas arvenses
The effect of light quality on maize: a tool for weed plants management
Ms.C. Yaisys Blanco,I Dra.C. Maha Afifi,II Dr.C. Clarence J. SwantonII
IInstituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32700.
IIUniversity of Guelph, 50 Stone Road E., Guelph, ON, N1G 2W1, Canada.
RESUMEN
Las señales de luz como el rojo y el rojo lejano (R/RL) son reflejadas desde el tallo y la superficie de las hojas de las arvenses vecinas, estas pueden desencadenar una respuesta por efecto de sombra en las plántulas de maíz, lo que resulta en cambios morfológicos. Dos experimentos de laboratorio se llevaron a cabo bajo condiciones controladas con el objetivo de determinar el efecto de las arvenses vecinas y la competencia por luz sobre plantas de maíz en etapas tempranas de su desarrollo. En el primer experimento, se utilizó Turface®; como suelo para plantar las semillas de maíz, y se estudiaron dos tratamientos (libre de arvenses y con arvenses). Raigrás (Lolium perenne L) se utilizó como especie de arvense. En todos los parámetros estudiados se observó una disminución en el experimento con arvenses después de las 48 h de la siembra excepto en la longitud del tallo, que antes de las 48 h fue mayor en el tratamientos libre de arvenses, ocurriendo el efecto contrario después de las 48 h. El segundo experimento, constó de tres tratamientos con diferentes densidades de siembra (baja, media y alta) con nueve, 28 y 81 plantas respectivamente. Los diferentes parámetros se comportaron de forma mayor en la densidad baja, excepto el largo del tallo, que fue mayor en la densidad alta, donde las plantas de maíz tuvieron una reacción de evitación por sombra de las arvenses, debido a la competencia por luz, ya que era el único factor limitante, incluso cuando las plantas eran mayores a la etapa V4. Este enfoque puede proporcionar una oportunidad para mejorar la competitividad del cultivo en condiciones de alta densidad.
Palabras clave: Zea mays, radiación, competencia interespecífica, Lolium perenne.
ABSTRACT
Light signals such as the red to far-red ratio (R/FR) reflected from stem and leaf surfaces of neighboring weeds can trigger a shade avoidance response in maize seedlings, resulting in morphological changes. Two different experiments were conducted under conditions of non-limiting resources with the objective to determine the effect of neighboring weeds competition for light in corn plants in early stages of development. In the first experiment; we used Turface® as soil for planting corn seeds, it consisted in two treatments (weedy –free and weedy). Perennial ryegrass (Lolium perenne L) was using as a model weed species, in all parameters studied, a decrease was observed in the experiment with weed after 48 h of planting except stem length 48 h before this was greater in the weed-free treatments opposite effect occurring after 48h. The second experiment consisted in three treatments with different densities(low, medium and high) with nine, 28 and 81 plant respectively, the effect occurred in the different parameters behaved low density higher, except in the stem large , which was greater in the high density. Where the corn plants had an avoidance reaction by weeds shade, due to light competition, because was the unique limitant factor, inclusively when the plants were high on V4 stage. This change in growth may help explain the importance of early-season weed control in corn.
Key words: Zea mays, radiation, interespecific competition, Lolium perenne.
INTRODUCCIÓN
La interferencia por arvenses es uno de los factores más importantes en la reducción de los rendimientos de los cultivos en todo el mundo (1). La competencia interespecífica entre los cultivos y las arvenses se produce principalmente por luz, agua, nutrimentos y espacio físico (2, 3).
La mayoría de las especies de plantas responden de manera diferente a la calidad de la radiación (color o longitud de onda) y a la cantidad de ella (densidad del flujo fotónico-DDF o irradiancia), así como las combinaciones de ambas, lo cual representa un factor fundamental en la interferencia entre cultivos y arvenses. El efecto de la calidad y la cantidad de radiación se combina con el componente ambiental de sombra que produce un espectro de radiación característico bajo el dosel de las plantas. Las hojas absorben fotones en el azul y rojo del espectro de la radiación fotosintéticamente activa (RFA), mientras que la absorción en el verde y especialmente en la región del rojo lejano es más débil y gran parte de estos fotones se refleja como radiación difusa (radiancia). Los precursores en demostrar la importancia de la proporción entre el rojo y el rojo lejano (R/RL), como componente fundamental de la sombra entre plantas vecinas, la captación temprana de esta señal por los entrenudos y su relación con la densidad del follaje, la cual modula la cantidad de radiación también demostraron que las plantas pueden detectar la presencia de plantas vecinas, mucho antes de que estén sombreadas (4, 5).
Las arvenses son una limitación importante en el cultivo del maíz Zea mays L.; se calcula una pérdida anual por rendimiento de hasta un 30 %, debido a los daños que realizan las mismas al cultivo (6). Por lo tanto, son un problema en la producción, principalmente en las primeras etapas de desarrollo del cultivo. Por otra parte, el período crítico para el control de arvenses en los cultivos comienza desde el crecimiento temprano del mismo; en el caso particular del cultivo en estudio, el período crítico es cuando aparece la tercera o cuarta hoja en la planta (7).
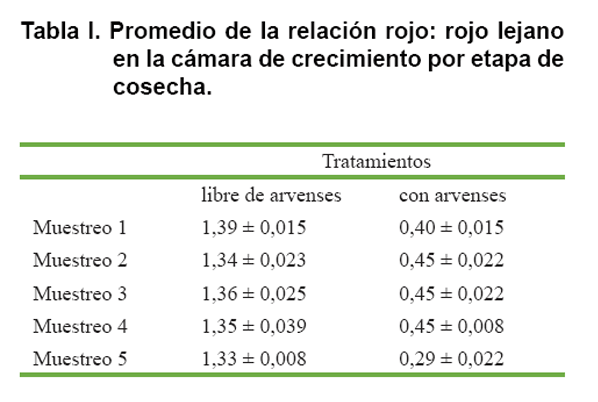
En este caso el recurso luz de la proporción entre el rojo y el rojo lejano (R/RL) juega un importante papel en la detección de las arvenses como plantas vecinas del maíz en una etapa muy temprana del crecimiento de las plantas (7, 10, 11). Los cultivos pueden ver estas plantas vecinas debido a la diferencia reflejada en la proporción entre el rojo y el rojo lejano (R/RL). Cuando la proporción entre el rojo y el rojo lejano (R/RL) en la luz reflejada es alta, las plantas detectan arvenses vecinas, mientras que en una baja proporción entre el rojo y el rojo lejano (R/RL) ocurre lo contrario (8, 9, 12, 13).
El tema no se ha abordado en toda su magnitud, pero algunos estudios han encontrado que hay una reducción en la biomasa de las raíces y en el volumen radical, tanto de trigo como de maíz, cuando se cultiva en condiciones de baja proporción entre el rojo y el rojo lejano (R/RL), comparado con las plantas cultivadas en altas proporciones (R/RL) (14, 15, 16). La reacción a las plantas vecinas es similar a la estrategia del efecto de sombra de las plantas; lo que reduce el número de granos final en las plantas de maíz (7, 10, 17). Dado que las plantas son capaces de detectar las plantas vecinas a muy temprana edad de su crecimiento es de vital importancia tener esto en cuenta para llevar a cabo un buen manejo de control de arvenses, para así evitar pérdidas de rendimiento durante el período vegetativo (18, 19, 20).
Debido a la importancia de la sombra y al efecto que ejercen las arvenses vecinas en el crecimiento del maíz, así como a que la mayoría de los trabajos no incluyen el componente de calidad, o sea, la longitud de onda de la radiación, el objetivo del presente trabajo fue determinar el efecto de las arvenses vecinas y la competencia por luz sobre plantas de maíz en etapas tempranas de su desarrollo.
MATERIALES Y MÉTODOS
Los experimentos fueron realizados en ambiente controlado en cámaras de crecimiento en el departamento de Plant Agriculture, en la Universidad de Guelph, Ontario, Canadá, utilizando el cultivar CG108 x CG102 (híbrido de maíz Zea mays L. de la Universidad de Guelph).
Los estados vegetativos que se utilizaron aparecen a continuación:
VE Emergencia
V1 Se ve el cuello de la primera hoja (la primera hoja siempre tiene la punta redondeada)
V2 Se ve el cuello de la segunda hoja
V3 Se ve el cuello de la tercera hoja
V4 Se ve el cuello de la cuarta hoja
Experimento I. Efecto de las arvenses en etapas de desarrollo temprano del cultivo.
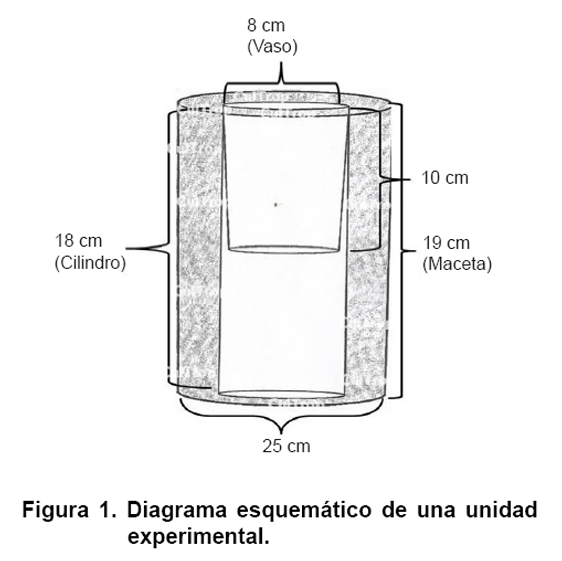
Las semillas de maíz fueron humedecidas con agua corriente durante 24 horas antes de la siembra. Una semilla por maceta fue sembrada en vasos plásticos de 355 mL (Dart Container Corporation, Mason, MI, USA) con 8 cm de diámetro y 10 cm de altura a 2 cm de profundidad. Los vasos plásticos fueron colocadas en cilindros (diámetro 8 cm, altura 18 cm, cilindro natural 1 L modificado a 18 cm, Consolidated Bottle Co., Toronto, ON, Canada). Posteriormente los cilindros fueron puestos en las macetas de 25 cm de diámetro (altura 19 cm, 6 L, Airlite Plastics Company, Omaha, NE, USA) (Figura 1).
En los tratamientos libre de arvenses y con arvenses las macetas y vasos plásticos fueron llenadas con Turface® 100 % arcilla calcinada con un tamaño de la partícula entre 2,5 y 3,5 mm (Turface MVP® Profile Products LLC, Buffalo Grove, IL, USA). En el caso del tratamiento con arvenses el área alrededor de los vasos plásticos fue sembrada con raigrás (Lolium perenne L. cv. Fiesta III) (dos a tres semanas antes de sembrarse las semillas de maíz).
Los experimentos fueron regados diariamente y fertilizados con una solución de nutrientes conteniendo N, P, K, Ca, Mg y con un suplemento de micronutrientes (21).
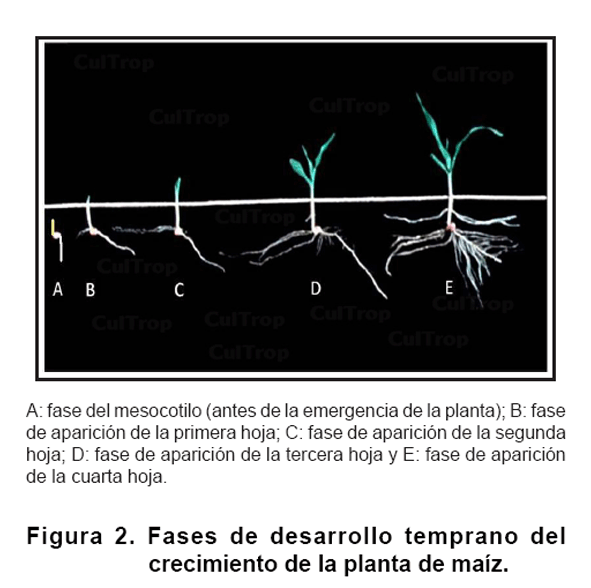
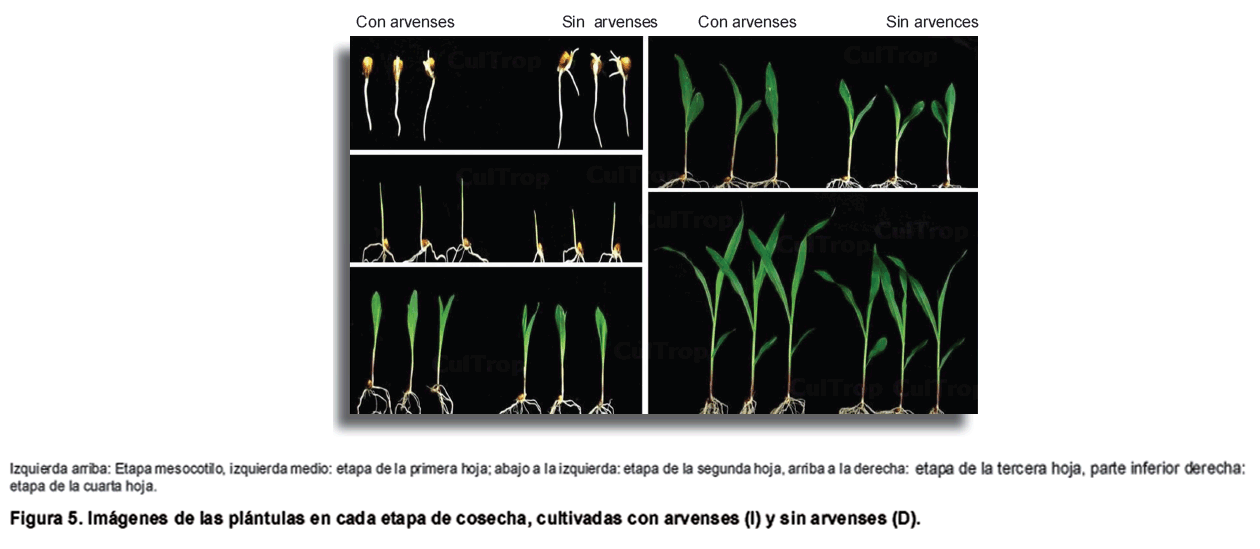
Las arvenses fueron cortadas cuando sobrepasaban las macetas para prevenir la sombra. Ocho plantas fueron extraídas al azar en cinco etapas diferentes de desarrollo: entre la etapa de mesocotilo (A) (B), (C), (D) y emisión de la cuarta hoja (E) (Figura 2). En total 40 plantas fueron extraídas por tratamiento.
La cámara de crecimiento en este experimento se dividió en dos con un divisor opaco de plástico (62 x 139 cm), para reducir la interferencia entre los dos tratamientos.
Las condiciones de la cámara de crecimiento fue la misma para las cinco repeticiones. La temperatura diurna fue de 28 oC y la nocturna de 20 oC. El fotoperiodo fue de 16 horas (desde las 6:00 am-10:00 pm).
La humedad relativa estuvo en el rango de 60-65 %. La irradiación fue proporcionada por el uso de tubos fríos fluorescentes blancos Sylvania intercalados con bombillos de tungsteno 40 Watts, para una densidad de flujo de fotones fotosintéticos (PPFD por sus siglas en inglés) de 250 µmol m-2 s-1.
Experimento II. Efecto de diferentes densidades de siembra en el desarrollo temprano del cultivo.
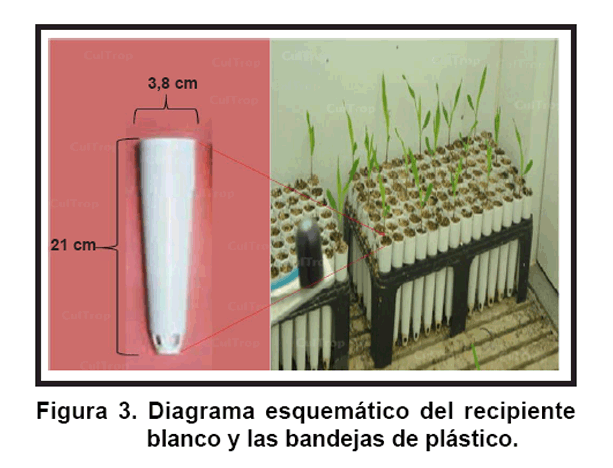
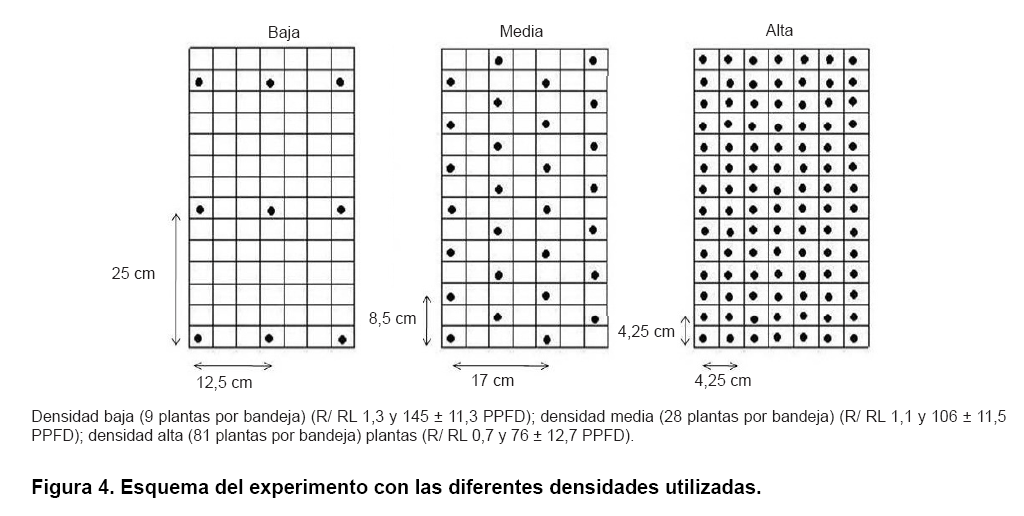
Con el fin de estudiar el efecto de diferentes densidades de siembra en el desarrollo temprano de plantas de maíz, se llevó a cabo un experimento preliminar para crear tres tratamientos con diferentes densidades de luz, bajo las mismas condiciones de cámara de crecimiento que se explican en el experimento anterior. Las semillas fueron plantadas a una profundidad de 2 cm en recipientes blancos (Ray Leach Cone-tainer Trays, Stuewe and Sons., Inc., Tangent, OR, USA) cubierto con una lámina de lana de roca (Grodan®; Grodania A/S, DK-2640 Hedelusene, Denmark) en la parte inferior, y después fueron llenados con Turface®. Estos recipientes se colocaron en bandejas de plástico (Ray Leach Cone-tainer Trays, Stuewe and Sons., Inc., Tangent, OR, USA) (Figura 3), con diferentes distancias para crear tres tratamientos diferentes. En el primer tratamiento que se consideró como un tratamiento de baja densidad (R/RL 1,3 y 145 ± 11,3 PPFD), se sembraron nueve plantas en una bandeja a 25 cm de distancia entre plantas. El tratamiento de densidad media consistió en 28 plantas a 8,5 y 17 cm entre las plantas (R/RL 1,1 y 106 ± 11,5 PPFD), mientras que el tratamiento de alta densidad constó de 81 plantas a 4,25 cm entre plantas (R/RL 0,7 y 76 ± 12,7 PPFD). La distancia entre las plantas en todos los tratamientos se cubrieron con recipientes de color blanco, los que se llenaron con Turface® (Figura 4).
Las bandejas de plástico, incluyendo los recipientes de color blanco se colocaron en una caja de plástico (del mismo tamaño que las bandejas) lleno de agua, para evitar el secado. Los recipientes se regaron dos veces al día usando una regadera y se cubrieron con una bandeja de plástico negro hasta la germinación de la semilla para mantener la humedad. Después de la emergencia de la planta, las plántulas se regaron una vez al día con un suplemento de micronutrientes (21). Este sistema aseguró que no hubiera ninguna competencia entre las plantas de maíz por agua o nutrimentos. Fueron extraídas 15 plántulas por tratamiento cuando las plantas estaban en la fase V4 (estadío vegetativo de la planta cuando aparece la cuarta hoja) para tener diferentes mediciones.
Mediciones morfológicas
En ambos experimentos se realizaron las siguientes mediciones: Longitud del tallo (cm), número de raíces de la corona (No.), diámetro del tallo (mm), biomasa de la raíz (g), biomasa del tallo (g), y la proporción tallo-raíz, estas variables se midieron para encontrar las diferencias en el desarrollo temprano de las plantas de maíz. Las características morfológicas se evaluaron una vez extraídas las plantas.
La longitud se midió con una regla graduada; el diámetro del tallo, utilizando un pie de rey (Mastercraft), pero esto sólo se midió a partir de la segunda extremidad de la hoja en adelante; la biomasa se obtuvo secando el material vegetal fresco en una estufa (80 oC) durante tres días. El material se pesó en una balanza analítica (Mettler Toledo).
Medición de la luz
La cantidad de luz (PPFD) y la calidad de la luz (R/RL) se midieron una vez al día durante el período experimental. El PPFD se midió a 10 cm por encima de las plántulas de maíz utilizando un punto cuántico radiómetro (LI-190SA; LI-COR Biosciences, Lincoln, NE, EEUU), con un sensor de coseno-corregido con cable de fibra óptica. Se midió la relación rojo/rojo lejano (R/RL) de la luz reflejada con regularidad durante el período de crecimiento con un sensor R/RL (SK R 110; Skye Instruments, Llandrindod Wells, Powys, Reino Unido). En el primer experimento se evaluó la reflectancia (capacidad de las superficies de reflejar la luz) a 10 cm por encima de la Turface® o las arvenses en cuatro posiciones elegidas al azar en cada tratamiento. La relación R/RL se midió después de la plantación de las semillas de maíz en los vasos plásticos y antes de cada cosecha. La relación R/RL, difirió entre tratamientos. La relación reflejada R/RL por tratamiento y por etapa se presenta en la Tabla I.
Diseño experimental y análisis estadístico
Todos los experimentos se diseñaron como bloques al azar. En estos experimentos, las repeticiones se definieron como entornos de la cámara de crecimiento en el tiempo y se combinaron para el análisis. El primer experimento fue repetido cinco veces. El segundo experimento se repitió tres veces. Todos los análisis estadísticos se realizaron en SAS 9.1 (SAS Institute, Cary, NC, EEUU), con una tasa de error de tipo 1 establecido en a=0,05. El test estadístico de Shapiro-Wilk se utilizó para probar la hipótesis de normalidad. No se requirió de transformaciones antes del análisis estadístico. Se analizaron las diferencias significativas entre los tratamientos utilizando la Prueba Tukey.
RESULTADOS Y DISCUSIÓN
Experimento I. Efecto de las arvenses en etapas de desarrollo temprano del cultivo.
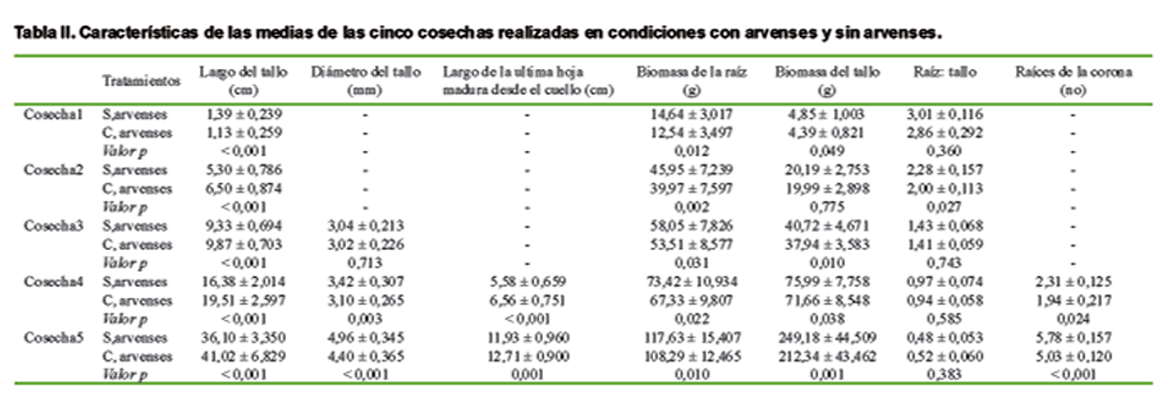
La longitud del tallo en la primera cosecha, después de las 48 horas, ya difería entre los tratamientos, a pesar de que no había brotado aún sobre la superficie del suelo. La longitud del tallo de las plántulas cultivadas en condiciones libres de arvenses fue significativamente mayor (1,39 ± 0,239 cm) que las plántulas cultivadas en condiciones con arvenses (1,13 ± 0,259 cm) (Tabla II y Figura 5), mientras que no hubo competencia de las raíces, debido a que las arvenses no hacían ningún contacto con las raíces de las plántulas.
A partir del segundo muestreo, el efecto contrario era visible en las plantas; la longitud del tallo en las plántulas que crecen bajo condiciones con arvenses fue significativamente mayor que la longitud del tallo de las plantas cultivadas en condiciones libres de arvenses. La mayor longitud del tallo para las plantas cultivadas en condiciones con arvenses se mantuvo visible durante el resto del experimento, lo que puede estar explicado por el efecto de interferencia que producía las arvenses sobre las plántulas.
El largo de la ultima hoja madura desde el cuello, fue visible a partir de la etapa V3 (estadío vegetativo cuando aparece la tercera hoja) en adelante, y tuvo resultados similares para la longitud del tallo; el cuello de la hoja de las plantas cultivadas en condiciones con arvenses fue significativamente mayor que el cuello de la hoja de las plantas cultivadas en condiciones libres de arvenses (Tabla II). Dado que tanto el largo del tallo y el cuello de la hoja son características de la altura de la planta, no se esperan diferencias, aunque en este caso especifico si se obtuvieron por la interferencia que había de las arvenses con las plántulas.
Resultados contrarios fueron visibles para la biomasa del tallo. La biomasa del tallo fue mayor en plántulas crecidas en condiciones libres de arvenses en todas las etapas de cosecha, excepto en la etapa V1 (estadío vegetativo cuando aparece la primera hoja), aunque el largo del tallo fue mayor cuando se sembró en condiciones con arvenses, desde la etapa V1 en adelante. Esta biomasa aérea mayor está en correspondencia con los resultados del diámetro del tallo de las plántulas. El diámetro del tallo, medida desde la etapa V2, fue mayor en las plantas cultivadas en condiciones libres de arvenses en la etapa V3 y V4. Cuando el diámetro del tallo se comparó frente a la longitud del tallo (Tabla II), se observó que el diámetro del tallo en condiciones con arvenses fue más pequeño que en condiciones libres de arvenses, incluso en la misma longitud del tallo. En la etapa V2, no se encontró ninguna diferencia en la biomasa de raíces. Además la biomasa radical tuvo los mismos resultados que la biomasa aérea. Con respecto a las plantas cultivadas en condiciones libres de arvenses obtuvieron una biomasa de raíces superior a las plántulas crecidas en condiciones con arvenses en todas las etapas de cosecha. La proporción raíz-tallo no fue diferente entre los tratamientos, excepto en la etapa V1, donde la proporción raíz: tallo fue significativamente mayor para las plantas cultivadas en condiciones libres de arvenses. El número de raíces corona, fue visible primero en la etapa V3, desarrollado en línea con la biomasa de las raíces. En condiciones libres de arvenses las plántulas habían producido un número significativamente mayor de raíces de la corona que en condiciones con arvenses. En la etapa V3, el promedio del número de raíces de la corona se comportó mayor en condiciones libres de arvenses en comparación con las condiciones con arvenses. Este efecto se hizo aún mayor en la etapa V4 en condiciones libres de arvenses comparada con las condiciones con arvenses.
De manera general, el crecimiento del maíz cultivado bajo condiciones de enmalezamiento, resultó con un reducido crecimiento de las plantas a las 48 horas después de la siembra, en comparación con las plantas cultivadas en condiciones libres de arvenses. Después de las 96 horas (etapa VE, emergencia-germinación) el efecto contrario era visible ya que se produjo una reacción para tratar de evitar la sombra en las plantas por la acción de las arvenses. Esto dio lugar a plantas más altas y más delgadas cuando se cultivaron en condiciones con arvenses. Por otra parte, el crecimiento de las raíces también se vió afectado, resultando más bajos la biomasa de la raíz y el número de raíces en la corona. Este resultado fue similar al encontrado por otros autores (16,17), quienes observaron que en las mismas condiciones las plantas presentaron menos número de raíces en la corona, bajo volumen total de las raíces y una menor biomasa de raíces cuando se cultiva en condiciones con arvenses ocurriendo lo contrario que cuando se cultiva en condiciones libres de arvenses.
Con el diámetro del tallo más pequeño resultaron las plantas que se sembraron bajo condiciones de enmalezamiento debido a la acción que hicieron las arvenses y como resultado hubo una reacción de evitación por sombra, resultados que coinciden con diversos autores (7, 9, 13), aunque estos estudios se llevaron a cabo bajo condiciones de invernadero y de campo, respectivamente, y las plantas fueron extraídas a partir de la etapa V4 en adelante, y en el caso de esta investigación solo se realizó hasta la etapa V4, lo que corrobora que en etapas tempranas de desarrollo de las plántulas también las arvenses ejercen una gran influencia en su crecimiento y desarrollo.
Con respecto a la proporción raíz-tallo hubo una disminución cuando las plantas fueron sembradas con arvenses lo que confirma estudios realizados al respecto (8, 9), donde también se obtuvieron resultados en la disminución raíz-tallo en plantas sembradas bajo el efecto de las arvenses, aunque los resultados se registraron después de la etapa V4. Excepto por la investigación realizada donde todas las observaciones se llevaron a cabo con plantas en la etapa V4 o más (16), otros estudios no han sido realizados con respecto al efecto de la proporción R/RL en plantas antes de la etapa V4; aunque en este estudio se encontró que la reacción de evasión por sombra en las plantas de maíz que se produce en condiciones con arvenses ya es visible desde la etapa V1, incluso sin ninguna competencia de las raíces.
Lo que no se explica todavía en la parte de arriba es la diferencia en el crecimiento en la fase de mesocotilo del maíz. El mesocotilo fue mayor cuando se sembró bajo condiciones libres de arvenses que cuando se cultivó con arvenses, mientras que esto cambió sólo 48 horas más tarde produciendo el efecto contrario. Esto significa que la plántula es capaz de visualizar la diferencia en la proporción R/RL en las primeras 48 horas de crecimiento de la planta. La pregunta sigue siendo por qué los resultados fueron opuestos después que la planta emergió, ya que ambas etapas no tenían competencia de las raíces y la diferencia entre la proporción R/RL fue la única diferencia que se produjo en el experimento, por lo que se deben realizar investigaciones relacionadas con este tema para conocer del por qué la planta tuvo esta respuesta.
Experimento II. Efecto de diferentes densidades de siembra en el desarrollo temprano del cultivo.
La longitud del tallo de las plántulas crecidas en diferentes densidades fue significativamente diferente en la etapa V4. Las plántulas cultivadas con una alta densidad fueron más altas (42,70 cm) que las plántulas crecidas en bajas densidades (39,68 cm) o media densidad (40,37 cm) (Tabla III). La longitud del tallo también difirió significativamente entre las densidades hasta el cuello de la segunda hoja. Las plantas que fueron sembradas con altas densidades tenían longitudes significativamente más altas (12,90 cm) que las plantas que se sembraron en densidades medias y bajas, pero también había diferencia entre plantas cultivadas bajo estas densidades. Las plántulas sembradas en bajas densidades tuvieron el cuello de la hoja significativamente menor (10,19 cm) que las plantas sembradas en densidades medias (10,91 cm). Lo opuesto se encontró para el diámetro del tallo, donde las plántulas cultivadas en bajas densidades tenían el diámetro de tallo mayor (5,04 mm), y las plántulas cultivadas en altas densidades tuvieron el diámetro del tallo más bajo con 4,61 mm.
Las plantas sembradas con una densidad media tuvieron un diámetro promedio del tallo de 4,81 mm, valores intermedios entre las densidades baja y alta. Estos resultados se muestran en concordancia con los de la biomasa de las raíces y la biomasa aérea, que fueron mayores para las plántulas sembradas a baja densidad y baja para las plántulas sembradas con alta densidad. Las plantas sembradas en densidad media no difirieron significativamente en la cantidad de biomasa de las raíces con las plantas sembradas en densidades bajas, mientras que la biomasa del tallo no fue diferente entre las plantas sembradas en las densidades bajas o altas.
Cuando las plantas de maíz se cultivaron con una alta densidad, se obtuvo como resultado una reacción de evitación por sombra de las arvenses, debido a la competencia por la luz, ya que era el único factor limitante, incluso cuando las plantas eran mayores a la etapa V4. En condiciones de bajas densidades de siembra, no se produjo el mismo efecto que ocurrió con la densidad anterior, debido al hecho de que no había competencia por la luz y la proporción R/RL era más alta que cuando se sembró bajo altas densidades de siembra.
Los efectos que la densidad de siembra tiene sobre el rendimiento final del maíz son conocidos desde tiempos atrás, pero el hecho de que los efectos de la densidad de plantación ya son visibles en la etapa V4 no se conocen todavía. No obstante, se ha confirmado que esta reacción de evitación por sombra es visible antes del cierre del dosel (22). Nuestra investigación está en concordancia con otros estudios donde se encontró que una alta densidad de siembra dio lugar a una reacción de evitación por sombra (13,23). Esta evitación por sombra se traduce en menores rendimientos finales en el maíz, por lo que es importante que los efectos de la evitación por sombra se reduzcan al mínimo, ya que esto dará lugar a rendimientos más altos (24). Esto contrasta con estudios previos realizados sobre densidades de siembra en el maíz, que indicaron que una alta densidad de plantación proporcionará un mayor rendimiento de las variedades con alto potencial productivo (20, 25, 26).
Ahora bien, con respecto a la proporción raíz-tallo apenas difirió de la biomasa, ya que las plantas sembradas en altas densidades tuvieron una proporción raíz-tallo inferior a las plantas sembradas en densidades de baja y media. Lo mismo se encontró para el número de raíces de la corona que fue 6,25 y 5,84 en las plantas sembradas en densidades baja y media, respectivamente, significativamente similares, y fue más bajo en plántulas crecidas bajo altas densidades (5,16 raíces).
En las imágenes de la Figura 6 es claramente visible que el volumen de la raíz es más pequeño cuando las plántulas se cultivan a altas densidades. Además de la diferencia en el volumen de la raíz, son también visibles las diferencias en la longitud y el diámetro del tallo.
La reacción de evitación por sombra en las plantas, es probablemente dirigida por hormonas como las auxinas (25, 26), las giberelinas (27), el etileno (27, 28, 29) y los brasinoesteriodes (26, 28).
Una baja proporción R/RL en los niveles de auxinas en las plantas superiores, podría ser una de las sustancias responsables del alargamiento, ya que regula las diferentes etapas del crecimiento de las plantas; por ejemplo, el metabolismo de la pared celular y la elongación celular (26, 28). Además de la elongación del tallo, las auxinas son también responsables de producir raíces más pequeñas, ya que las auxinas son transportadas desde las hojas jóvenes hacia abajo a través del tallo llegando a las raíces (25, 26).
Cuando las plantas se cultivan con bajas densidades de siembra, la proporción R/RL de distribución de las auxinas cambiarán, lo que resulta en una menor cantidad de auxinas que llegará a las hojas, y en una reducción del crecimiento de la raíz (26).
Por otra parte, la germinación de las semillas y el crecimiento de las plantas están reguladas por las giberelinas, las cuales van a estar afectadas por los fitocromos, los receptores de luz; por ejemplo la calidad de la luz (28). Estos autores sugirieron que los controles del fitocromo del alargamiento del tallo en las plantas son a través de la regulación de una giberelina. En un estudio realizado se encontró que la giberelina juega un papel importante en la respuesta a la baja proporción R/RL y la luz azul, ya que los mutantes deficientes de las giberelinas en tabaco no respondieron a diferentes proporciones R/RL mientras que los tipos silvestres sí lo hicieron (27).
Las giberelinas afectan la regulación de otras hormonas vegetales como, el etileno, el cual requiere de giberelinas para producir una reacción de evitación por sombra (26). Bajas proporciones R/RL conducen a un aumento de etileno en la planta, lo que resulta en fenotipos similares a los de las plantas que tienen reacciones de evitación por sombra (26, 28). También la deficiencia de etileno en mutantes de tabaco hizo que no fueran capaces de competir por luz, mientras que las plantas de tabaco de tipo silvestre fueron capaces de competir. El alargamiento en las plantas es el resultado del bajo nivel de etileno que las mismas producen. Esta hormona está regulada probablemente junto a las señales de luz como las proporciones R/RL, lo que resulta en la reacción de evitación por sombra (28). Como se indicó anteriormente las hormonas vegetales son la causa de las diferencias en el crecimiento de las plántulas en nuestro experimento, ya sea en condiciones libres de arvenses o con arvenses, así como en condiciones de diferentes densidades de siembra. Las hormonas son responsables del crecimiento, por ejemplo, el alargamiento del tallo y de tal modo del diámetro del mismo, pero también para la raíz y el número de raíces de la corona.
De acuerdo a estos resultados se puede concluir que la morfología de la planta se ve afectada por las plantas vecinas, ya sean arvenses o plantas de su misma especies; además, que las plantas reaccionan en estado temprano de su crecimiento, ya que la primera reacción fue visible a las 48 horas después de la siembra.
Después de las 96 horas las primeras características de reacción por evitar la sombra común eran visibles, en las plántulas sembradas en condiciones bajo arvenses, los tallos eran más altos y tenían un diámetro del tallo más pequeño que las plantas que fueron sembradas en condiciones libres de arvenses. Dado que las plantas son capaces de visualizar las arvenses vecinas en su desarrollo temprano, es importante tener en cuenta el control de las arvenses en etapas tempranas del crecimiento del cultivo del maíz, por la reacción de evitar la sombra debido a las arvenses lo cual ocasionará rendimientos más bajos.
BIBLIOGRAFÍA
1. Oerke, E. C. Crop losses to pests. The Journal of Agricultural Science, 2006, vol. 144, no. 1, pp. 31-43. ISSN: 0021-8596.
2. Scoot, T. /et al./. Weeds and the red to far-red ratio of reflected light: characterizing the influence of herbicide selection, dose, and weed species. Weed Science, 2011, vol. 59, pp. 424-430. ISSN: 0043-1745.
3. Moriles, J. C. /et al./. Microarray and growth analyses identify differences and similarities of early corn response to weeds, shade, and nitrogen stresses. Weed Science, 2012, vol. 60, pp. 158-166. ISSN: 0043-1745.
4. Ballaré, C. L.; Scopel, A. L. y Sánchez, R. A. Photocontrol of stem elongation in plant neighbourhoods: effects of photon fluence rate under natural conditions of radiation. Plant Cell and Environment, 1991, vol. 14, pp. 57-65. ISSN: 0140-7791.
5. Page, E. R.; Tollenaar, M.; Lee, E. A.; Lukens, L. y Swanton, C. J. Does the shade avoidance response contribute to the critical period for weed control in maize (Zea mays). Weed Research, 2009, vol. 49, pp. 563-571. ISSN: 0043-1737.
6. Rajcan, I.; Chandler, K. J. y Swanton, C. J. Red-far-red ratio of reflected light: a hypothesis of why early-season weed control is important in corn. Weed Science, 2004, vol. 52, pp. 774-778. ISSN: 0043-1745.
7. Liu, J. G.; Mahoney, K. J.; Sikkema, P. H. y Swanton, C. J. The importance of light quality in crop-weed competition. Weed Research, 2009, vol. 49, pp. 217-224. ISSN: 0043-1737.
8. Silva, P. S. L.; Souza, A. D.; Paula, V. F. S.; Oliveira, F. H. T. y Silva, K. M. B. Influence of corn sowing density and gliricidia intercropping on weed control. Planta Daninha, 2010, vol. 28, pp. 271-279. ISSN: 0100-8358.
9. Markham, M. Y. y Stoltenberg, D. E. Red: far-red light effects on corn growth and productivity in field environments. Weed Science, 2009, vol. 57, pp. 208-215. ISSN: 0043-1745.
10. Page, E. R.; Cerrudo, D.; Westra, P.; Loux, M.; Smith, K.; Foresman, C.; Wright, H. y Swanton, C. J. Why early season weed control is important in maize. Weed Science, 2012, vol. 60, pp. 423-430. ISSN: 0043-1745.
11. Sparkes, D. L.; Berry, P. y King, M. Effects of shade on root characters associated with lodging in wheat (Triticum aestivum). Annals of Applied Biology, 2008, vol. 152, pp. 389 -395. ISSN: 0003-4746.
12. Afifi, M. y Swanton, C. J. Maize seed and stem roots differ in response to neighbouring weeds. Weed Research, 2011, vol. 51, pp. 442-450. ISSN: 0043-1737.
13. Afifi, M. y Swanton, C. J. Early Physiological Mechanisms of Weed Competition. Weed Science, 2012, vol. 60, no. 4, pp. 542-551. ISSN: 0043-1745.
14. Green-Tracewicz, E.; Page, E. R. y Swanton, C. J. Shade avoidance in soybean reduces branching and increases plant-to-plant variability in biomass and yield per plant. Weed Science, 2011, vol. 59, pp. 43-49. ISSN: 0043-1745.
15. Merotto Jr. A.; Fischer, A. J. y Vidal, R. A. Perspectives for using light quality knowledge as an advanced ecophysiological weed management tool. Planta Daninha, 2009, vol. 27, pp. 407-419. ISSN: 0100-8358.
16. Cerrudo, D.; Page, E. R.; Tollenaar, M.; Stewart, G. y Swanton, C. J. Mechanisms of yield loss in maize caused by weed competition. Weed Science, 2012, vol. 60, pp. 225-232. ISSN: 0043-1745.
17. Tollenaar, M. Response of dry matter accumulation in maize to temperature: I. Dry matter partitioning. Crop Sci., 1989, vol. 29, pp. 1239-1246. ISSN: 0011-183X.
18. Ballaré, C. L. Illuminated behaviour: phytochrome as a key regulator of light forging and plant anti-herbivore defence. Plant Cell Environ., 2009, vol. 32, pp. 713-725. ISSN: 1365-3040.
19. Page, E. R.; Liu, W.; Cerrudo, D.; Lee, E. A. y Swanton, C. J. Shade avoidance influences stress tolerance in maize. Weed Science, 2011, vol. 59, pp. 326-334. ISSN: 0043-1745.
20. Markham, M. Y. y Stoltenberg, D. E. Corn morphology, mass, and grain yield as affected by early-season red:far-red light environments. Crop Science, 2010, vol. 50, pp. 273-280. ISSN: 0011-183X.
21. Wang, J. G.; Chen, C. H.; Chien, C. T. y Hsieh, H. L. Far-red insensitive 219 modulates constitutive phytomorphogenic1 activity via physical interaction to regulate hypocotyl elongation in Arabidopsis. Plant Physiol., 2011, vol. 156, pp. 631-646. ISSN: 0032-0889.
22. Keuskamp, D. H. S. /et al./. Auxin transport through PIN-FORMED 3 (PIN3) controls shade avoidance and fitness during competition. PNAS, 2010, vol. 107, pp. 22740-22744. ISSN: 0027-8424.
23. Morelli, G. y Ruberti, I. Shade avoidance responses. Driving auxin along lateral routes. Plant Physiology, 2000, vol. 122, pp. 621-626. ISSN: 0032-0889.
24. Vandenbussche, F.; Pierik, R.; Millenaar, F. F.; Voesenek, L. A. C. J. y Van Der Straeten, D. Reaching out of the shade. Current Opinion in Plant Biology, 2005, vol. 8, pp. 462-468. ISSN: 1369-5266.
25. Pierik, R.; Cuppens, M. L. C.; Voesenek, L. A. C. J. y Visser, E. J. W. Interactions between ethylene and gibberellins in phytochrome-mediated shade avoidance responses in tobacco. Plant Physiology, 2004, vol. 134, pp. 2928-2936. ISSN: 0032-0889.
26. Tao, Y. /et al./. Rapid synthesis of auxin via a new thryptophan-dependent pathway is required for shade avoidance in plants. Cell, 2008, vol. 133, pp. 164-176. ISSN: 0092-8674.
27. Bhalerao, R. P. /et al./. Shoot-derived auxin is essential for early lateral root emergence in Arabidopsis seedlings. The Plant Journal, 2002, vol. 29, pp. 325-332. ISSN: 1365-313X.
28. Kamiya, Y. y García-Martínez, J. L. Regulation of gibberellin biosynthesis by light. Current Opinion in Plant Biology, 1999, vol. 2, pp. 398-403. ISSN: 1369-5266.
Recibido: 2 de julio de 2013
Aceptado: 6 de marzo de 2014
Ms.C. Yaisys Blanco, Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32700. Email: yblanco@inca.edu.cu


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}