










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCultivos Tropicales
versión impresa ISSN 0258-5936versión On-line ISSN 1819-4087
cultrop vol.37 no.4 La Habana oct.-dic. 2016
http://dx.doi.org/10.13140/RG.2.2.10964.19844
http://dx.doi.org/10.13140/RG.2.2.10964.19844
Reseña
El rol de las arvenses como componente en la biodiversidad de los agroecosistemas
The role of weeds as a component of biodiversity in agroecosystems
M.Cs. Yaisys Blanco-Valdes
Instituto Nacional Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32700.
Resumen
La agricultura considera a las arvenses como indeseables dentro de los agroecosistemas. Sin embargo, una gran mayoría de campesinos del trópico, las estiman por su valor y beneficio que aportan en un sistema agrícola. En el presente escrito se da a conocer la importancia de incorporar diferentes disciplinas científicas y sistemas de manejo de arvenses en una agricultura con menos impacto en el ambiente. Además, se hace mención de la utilidad de arvenses importantes como plantas: biocidas, medicinales, mejoradoras del suelo, fuente de nuevos cultivos y alimentación para humanos y animales.
Palabras clave: agroecología, manejo, competencia, arvenses, agricultura.
ABSTRACT
Agriculture consider weeds as undesirable in agroecosystems. However, a great amount of farmers of the tropics, estimate then by their value and benefit they bring in an agricultural system. In this paper, it is disclosed the importance of incorporating different scientific disciplines and weed management systems in agriculture with less impact on the environment. Besides it is refered to the usefulness of important weeds as plants: biocides, medicinal, soil improve, new crops and source of food for humans and animals.
Key words: agroecology, management, competition, weeds, agriculture.
INTRODUCCIÓN
La agroecología se perfila como la ciencia fundamental para orientar la conversión de sistemas convencionales de producción (monocultivos dependientes de insumos agroquímicos) a sistemas más diversificados y autosuficientes. Para esto, la misma utiliza principios ecológicos que favorecen procesos naturales e interacciones biológicas que optimizan sinergias de modo tal que la agrobiodiversidad sea capaz de subsidiar por si misma procesos claves tales como la acumulación de materia orgánica, fertilidad del suelo, mecanismos de regulación biótica de plagas y la productividad de los cultivos. Estos procesos son cruciales pues condicionan la sustentabilidad de los agroecosistemas. La mayoría de estos procesos se optimizan mediante interacciones que emergen de combinaciones específicas espaciales y temporales de los cultivos, los animales y los árboles, complementados por manejos orgánicos del suelo (1).
Esta nos proporciona guías para desarrollar agroecosistemas diversificados que aprovechen la integración de la biodiversidad animal y vegetal. La integración exitosa de las plantas y los animales puede fortalecer interacciones positivas y optimizar las funciones y los procesos del agroecosistema, tales como la regulación de los organismos dañinos, el reciclaje de nutrientes, la producción de biomasa y la formación de materia orgánica. De esta manera los agroecosistemas se vuelven mas resilientes (1, 2).
En la actualidad existe una gran cantidad de prácticas y las tecnologías para mejorar el funcionamiento de los agroecosistemas. Cuando los agroecosistemas son desarrollados para estar en sintonía con las condiciones ambientales y socioeconómicas existentes, el resultado final es una mayor sostenibilidad ecológica (3). Al adoptar prácticas de manejo ecológico el agricultor puede incrementar la estabilidad y la resiliencia del agroecosistema (4).
Hoy en día las arvenses han ganado espacio en los sistemas de cultivos, a través de una agricultura convencional, pues se considera que la presencia de diferentes especies de arvenses dentro de los cultivos, tiene un profundo impacto en la composición e interacciones de la entomofauna del cultivo, a tal punto que los predadores y los parasitoides son más efectivos en los hábitats complejos (5). Los insectos benéficos tienen mayores posibilidades de encontrar presas alternativas, abrigo, sitios para reproducción y refugios para dormancia (6, 7), además, se deben aprovechar otros beneficios que estas otorgan como: insecticidas, fungicidas repelentes, alimentao humano y animal, mejoramiento genético, medicinal y conservación del suelo (6, 8, 9).
La implementación de la combinación de los aspectos anteriores planteados, dan la pauta para establecer una estrategia en una agricultura sostenida, por efecto de un mejor impacto sobre el comportamiento de las arvenses, la reducción de la energía, la disminución de las perturbaciones continuas de los sistemas agrícolas e incremento en el número de interacciones biológicas, indispensables para mantener el equilibrio en los agroecosistemas. Por lo que a partir de lo expuesto anteriormente el objetivo de este trabajo es el de no perder el conocimiento empírico, sobre la competitividad y la utilidad del gran número de plantas que crecen en los campos agrícolas, los frutales, los huertos, los pastos, los forestales y otros sitios los cuales está aprovechando el ser humano.
BIODIVERSIDAD ASOCIADA
La biodiversidad asociada está definida como todas las especies de plantas, animales y microorganismos existentes que interactúan dentro de un ecosistema. En todos los agroecosistemas, los polinizadores, los enemigos naturales, las lombrices de tierra y los microorganismos del suelo son componentes clave de la biodiversidad y juegan papeles ecológicos importantes, al mediar procesos como la introgresión genética, el control natural, el reciclaje de nutrientes, la descomposición, entre otros (1, 10, 11).
Históricamente, la diversidad en la agricultura ha demostrado ser una vía para proteger a los agricultores de las plagas y enfermedades. Por el contrario, el camino de la especialización y el monocultivo provocan el aumento de la contaminación, por el uso de los agrotóxicos y fertilizantes , lo que conlleva a la degradación de recursos naturales. Como consecuencia se asiste a un proceso acelerado de “erosión genética” de las especies cultivadas, que ocurre por la sustitución de variedades, de gran diversidad y adaptación por cultivares denominados “modernos”, obtenidos a través de la manipulación y selección del material genético (12–14).
La biodiversidad en los agroecosistemas puede ser un indicador del grado de deterioro de los mismos. Se plantea que es la base para garantizar el suministro mundial de los alimentos, la supervivencia de los cultivos, los paisajes agrícolas y el seguro de la humanidad contra futuras amenazas a la agricultura y la alimentación. Particularmente, la diversidad agrícola o agro diversidad es un concepto, que reúne lo relativo a la diversidad biológica para la producción agrícola y comprende los recursos genéticos de las plantas, los animales, los organismos del suelo, los insectos y otros organismos en ecosistemas manejados o agroecosistemas; además, los elementos de ecosistemas naturales para la producción de alimentos (5, 6, 15).
En los últimos años, los científicos han comenzado a darle mayor importancia al papel que juega la biodiversidad en el funcionamiento de los sistemas agrícolas, considerando que es precisamente el principio fundamental de la agricultura sostenible (2, 5).
CLASIFICACIÓN
Se pueden reconocer dos tipos de componentes de la biodiversidad: el primero, la biodiversidad planificada, es la biodiversidad asociada con los cultivos y animales incluidos en el agroecosistema por el agricultor que variará de acuerdo con el manejo y los arreglos de cultivos (16). El segundo, la biodiversidad asociada, incluye la flora y la fauna del suelo, los herbívoros, los descomponedores y los depredadores, que colonizan el escenario productivo desde los ambientes circundantes y que permanecerán en el mismo dependiendo del tipo de manejo adoptado. Por lo que resulta clave identificar el tipo de biodiversidad que es deseable de mantener o incrementar, de manera que se puedan llevar a cabo las funciones o servicios ecológicos, para determinar cuáles son las mejores prácticas de manejo e incrementar la biodiversidad deseada (9, 17, 18).
Un agroecosistema debe tener alrededor de 150 especies agrícolas, para ser considerado de buena diversidad. Mientras más diversos son los sistemas productivos, más complejos y estables resultan; cuantos más componentes biológicos haya en los sistemas, mayores mecanismos de autorregulación habrá y entre mayores sean los mecanismos de autorregulación, mayor será el equilibrio de los sistemas (19–21).
AGROECOLOGÍA
Es la disciplina científica que enfoca el estudio de la agricultura desde una perspectiva ecológica y se define como un marco teórico, cuyo fin es analizar los procesos agrícolas de manera más amplia.
El enfoque agroecológico considera a los ecosistemas agrícolas como las unidades fundamentales de estudio y en estos sistemas, los ciclos minerales, las transformaciones de energía, los procesos biológicos y las relaciones socioeconómicas son investigadas y analizadas como un todo (2, 22).
La agroecología es una disciplina relativamente nueva, pero se nutre de la experiencia y el saber milenario de tradicionales sociedades agrícolas que la agricultura “moderna” buscó marginar y desarticular. Esta propone un enfoque de la agricultura, desde una perspectiva ecológica, que pretende construir un marco teórico, cuyo fin es analizar los procesos agrícolas de la manera más amplia. Propone una perspectiva sistémica, que posibilita estudiar las condicionantes de la sociedad global en las condiciones de producción agraria y considera los agroecosistemas como unidades, basa su accionar en un entendimiento profundo de su naturaleza y los principios que la gobiernan (2, 23).
La idea de la agroecología es ir más allá del uso de prácticas alternativas y desarrollar agroecosistemas con una dependencia mínima de los agroquímicos y los subsidios de la energía, haciendo énfasis en los sistemas agrícolas complejos, en los cuales las interacciones ecológicas y los sinergismos, entre sus componentes biológicos, proveen los mecanismos para que los sistemas subsidien la fertilidad de su propio suelo, la productividad y la protección de los cultivos (24).
La agroecología provee las bases ecológicas para la conservación de la biodiversidad en la agricultura, además del rol que ella puede jugar en el restablecimiento del balance ecológico de los agroecosistemas, de manera que alcance una producción sustentable.
La biodiversidad promueve una variedad de procesos de renovación y servicios ecológicos en los agroecosistemas (reciclaje de nutrientes, control biológico de plagas y conservación del agua y del suelo); cuando estos se pierden, los costos pueden ser significativos (25).
El auge de la revolución verde es uno de los factores de mayor peso en el deterioro de los agroecosistemas, no solo por el impacto ambiental que causan las grandes cantidades de productos químicos que se vierten al medio y provocan desequilibrios ecológicos, sino también por la erosión genética de las plantas de cultivos. El sistema de extensas áreas de monocultivo utilizado, trajo consigo el deterioro a largo plazo de la diversidad, aumento de la resistencia a plagas, así como el detrimento de la fertilidad de los suelos. Sin embargo, se ha demostrado que las innovaciones tecnológicas y el desarrollo agrícola no han hecho un gran impacto en áreas campesinas, donde los sistemas de manejo permanecen inalterables en algunos casos (26).
Es posible también identificar las causas indirectas en la pérdida de la biodiversidad, como son las políticas nacionales e internacionales, los derechos de propiedad y las influencias del mercado. Todas estas condiciones hacen referencia a la importancia de analizar las relaciones humanas con los recursos de la biodiversidad dentro de los modelos de desarrollo (27).
CONCEPTO Y GENERALIDADES
Se consideran como arvenses a todas las plantas superiores, que por crecer junto o sobre plantas cultivadas, perturban o impiden el desarrollo normal, encarecen el cultivo y merman sus rendimientos o la calidad (28).
Algunos autores, mencionan que las arvenses son especies vegetales que conviven con los cultivos económicos y su manejo es considerado como la actividad de selección y conservación de coberturas nobles, que evitan la competencia interespecífica durante su período crítico y simultáneamente contribuyen a la protección del recurso natural suelo (29, 30).
Por otra parte, otros argumentan que las arvenses son plantas silvestres que crecen en hábitats frecuentemente disturbados por la actividad humana. Una planta es arvense si, en cualquier área geográfica específica, sus poblaciones crecen sin que sean cultivadas con deliberación (31).
Las arvenses son importantes porque tienen efectos negativos sobre las actividades del ser humano y por los costos en los que se incurre en su manejo para mantener las poblaciones a un nivel que no reduzca el rendimiento del cultivo y no interfieran con las actividades de los humanos ni causen repulsión a la vista (4, 32).
Aspectos negativos:
- Costos por manejo.
- Dificultan y demoran las labores agrícolas.
- Son hospedantes de plagas.
- Reducen el rendimiento de los cultivos.
- Reducen la calidad del producto.
- Envenenan a los animales.
- Causan problemas de salud al hombre.
- Disminuyen el valor de la tierra.
Entre los aspectos positivos de las arvenses se nombran los siguientes (4):
- Contribuyen a la conservación del suelo.
- Son fuente de alimento como algunas gramíneas y leguminosas.
- Sirven como medicinas.
- Incrementan la cantidad de material genético.
- Incrementan la estabilidad del agroecosistema.
- Son fuente de materia prima para la elaboración de fertilizantes orgánicos.
Principales características de las arvenses
Los principales atributos morfológicos y reproductivos para que una especie sea exitosa como arvense, son los siguientes (33, 34):
- Producción de semilla abundante
- Germinación, dispersión y latencia de las semillas
Crecimiento vegetativo
Clasificación
Según su hábitat, pueden ser: agrestes, ruderales, arvenses de pasturas y arvenses acuáticas:
- Según el tipo de hoja: hoja ancha y hoja angosta.
- Según la consistencia del tallo: leñosas, semileñosas y herbáceas.
- Según el ciclo de vida: anuales o perennes.
- Según su nocividad: puede ser alta, mediana o levemente nocivas.
IMPORTANCIA ECONÓMICA DE LAS ARVENSES
Las arvenses compiten con los cultivos por los nutrientes del suelo, el agua y la luz; hospedan insectos y patógenos dañinos en las plantas de los cultivos y sus exudados de raíces y filtraciones de las hojas pueden ser tóxicos para las plantas cultivadas. Además interfieren con la cosecha del cultivo e incrementan los costos de tales operaciones; en la cosecha, las semillas de estas pueden contaminar la producción. Por lo tanto, la presencia de arvenses en las áreas de cultivo reduce la eficiencia de los insumos tales como el fertilizante y el agua de riego, fortalecen la densidad de otros organismos y plagas, finalmente, reducen severamente el rendimiento y la calidad del cultivo (35).
Se han multiplicado las investigaciones y estudios sobre interacciones entre arvenses como plantas hospedantes, como indicadores ecológicos, como fuente de biomasa y agentes alelopáticos en sistemas de cultivos tropicales. Sin embargo, la necesidad del manejo de arvenses se continúa investigando con el fin de encontrar la forma más eficiente de manejar o controlar y en algunos casos, erradicar especies o poblaciones de estas plantas en los ecosistemas agrícolas del trópico (36).
Investigaciones realizadas en diferentes cultivos del trópico se han estudiado únicamente la eficiencia de los métodos químicos y los mecánicos aplicados; esto no entusiasmó y fueron incluidos estudios sobre la fenología del cultivo y de las arvenses presentes (37). A partir de aquí fue el inicio para una base de datos con informaciones sobre la biología de las arvenses tropicales y sus interacciones con cultivos anuales, perennes, la flora y la fauna correspondiente.
En Cuba, Nicaragua y México fueron desarrollados estudios en cafetales sobre la composición de la flora adventicia, los efectos que causaron los diferentes métodos de control de las arvenses en las décadas pasadas y su impacto en la estabilidad ecológica de la fauna y la flora (34, 35).
Como resultado de estas investigaciones surgió, que se deben mejorar los conocimientos herbológicos sobre la flora adventicia por parte de los productores y los asesores, así mismo como se requiere la implementación del monitoreo en las fincas; éstos son el fundamento para un manejo sostenible de las arvenses basado en decisiones correctas en cuanto al momento del control y del método adecuado por su eficiencia económica e inocuidad ecológica.
LAS ARVENSES Y EL AGROECOSISTEMA
La Agricultura constituye la mayor fuerza selectiva en la evolución de las arvenses. Como consecuencia de haber desplazado la sucesión hacia estados tempranos en forma recurrente, las actividades agrícolas han mantenido las comunidades vegetales en estadíos inmaduros. La mayoría de los componentes de esas comunidades son lo que en la agricultura llamamos arvenses. Este término involucra a unas 250 especies vegetales que no constituyen una clase botánica en particular (38).
Muchas de ellas se han introducido desde áreas geográficas muy distantes o nativas y son particularmente favorecidas por las perturbaciones causadas por la actividad agrícola. Cualquiera que sea su origen, las arvenses son un componente integral de los agroecosistemas y como tales influencian la organización y el funcionamiento de los mismos desde los albores de la agricultura (39, 40).
De manera tradicional y principalmente debido a su impacto sobre el rendimiento, las arvenses se han considerado organismos indeseables. Tanto en la literatura como en la tradición agrícola es muy profundo el sentimiento de aversión que estos organismos vegetales despiertan en el ser humano. En general, están asociadas a maldad, descuido, daño, pérdida o inconvenientes de algún tipo (41).
Desde tiempos remotos y hasta épocas recientes, el problema de las arvenses en los cultivos fue enfocado desde el punto de vista de su exclusión del cultivo.
El esfuerzo por lograr ese objetivo ha sido descomunal y al tiempo que demuestra las habilidades del ser humano para desarrollar diferentes prácticas de eliminación o control, desnuda la ingenuidad con que ha sido enfocado el problema, salvo excepciones, las arvenses de los cultivos son tanto problema en la actualidad como un siglo atrás (38).
La gran diferencia está basada en la gama de herramientas tecnológicas hoy disponibles que permiten diseñar secuencias de cultivos con bajos niveles de infestación. Esto ha sido posible gracias a la investigación básica y aplicada relacionada con el estudio de los mecanismos fisiológicos de absorción, transporte, acción tóxica y al desarrollo de síntesis y producción de herbicidas. Se disponen hoy de más de 130 principios activos. El mercado mundial de agroquímicos moviliza una cifra superior a los 16 000 millones de dólares por año: los herbicidas significan un 60 % de esta cifra y han contribuido en forma significativa al logro de los altos niveles de producción de las últimas décadas (37, 42).
Hasta aquí, podemos concluir que los problemas de arvenses de la actualidad son de similar envergadura que los existentes en el pasado y que la diferencia estriba en el rango de tecnologías que disponemos para enfrentarlas. Estas herramientas tecnológicas no son utilizadas en la mayoría de los casos con la eficiencia que demandan los tiempos actuales (43, 44).
Una cuestión adicional se debe plantear: los grandes avances tecnológicos operados a partir de mediados del siglo XX han causado un cambio de importancia en las ideas centrales de la agricultura y han estado basadas en el uso intensivo de agroquímicos, de variedades e híbridos con altísimo potencial de rendimiento en condiciones de alta disponibilidad de recursos y en la reducción del requerimiento de trabajo humano a través del empleo generalizado de nuevas y potentes maquinarias y alto consumo de energía fósil. A este sistema se lo reconoce actualmente como agricultura convencional (45).
Ya en 1973 se alertó sobre las consecuencias de la concepción descrita y se plantearon posibles alternativas en el informe “Los límites del crecimiento”, el cual si bien contenía algunas imprecisiones, planteaba la duda acerca de la sustentabilidad del sistema convencional (39).
La primera aproximación se ha ido enriqueciendo con información y evidencias que demuestran que con la tecnología promovida por la agricultura convencional ha ocurrido al menos en algunas áreas del planeta una fuerte contaminación de aguas superficiales y subterráneas, se ha incrementado la erosión del recurso suelo, han aparecido formas de resistencia en plagas, y empiezan a registrarse residuos de plaguicidas en ciertos alimentos. Desde el punto de vista energético la agricultura convencional exhibe un balance de energía fuertemente negativa (32).
A partir de este diagnóstico, se han propuesto distintas alternativas tecnológicas que con diversas variaciones semánticas (sostenibilidad, sustentabilidad, agricultura orgánica), intentan el planteo más o menos amplio de nuevos sistemas de manejo que eliminen o reduzcan la manifestación de los procesos antes enunciados (2, 46).
Con este enfoque, se han implementado programas de trabajo en EE.UU. y en Europa bajo la sigla generalizada de (LIFE), LISA, instrumento financiero para el medio ambiente, entre otras. En la Argentina, el Proyecto de Agricultura Conservacionista (PAC), lanzado hace algunos años por el Instituto Nacional de Tecnología Agropecuaria (INTA), se inscribe en la misma tendencia. Como regla general, los programas mencionados intentan la generación de tecnologías que permita la obtención de niveles de producción similares o algo inferiores a los actuales, sostenibles a largo plazo y con una sustancial reducción en algunos casos del 50 % en el uso de los agroquímicos, los combustibles y las labranzas (47, 48).
Resulta claro que el nuevo enfoque no significa un retorno de los métodos anteriores a la Revolución Verde, sino básicamente en una combinación selectiva de las prácticas proporcionadas por la tecnología moderna (27).
Si se tiene en cuenta que los ecosistemas bajo cultivo ocupan en la actualidad casi los dos tercios de la superficie terrestre y que contribuyen con el 20 % de la productividad neta del planeta, resulta claro que el protagonismo de los agricultores y técnicos que toman y ejecutan decisiones en tal sistema es de una envergadura superior a la de cualquier otro grupo social del planeta (26).
Por lo tanto, este replanteo de procesos productivos requiere necesariamente de productores y técnicos con un muy buen nivel de conocimientos acerca del sistema en que actúan. En síntesis, la toma de decisiones y la implementación de medidas y prácticas que permitan la sustentabilidad de los sistemas agropecuarios es una cuestión de alta tecnología (14).
En ese contexto, el manejo de plagas animales y vegetales ocupa un lugar central. Como se ha dicho en el comienzo, las arvenses forman parte de un ecosistema altamente perturbado y en estado de etapa inicial de la sucesión secundaria. El enmalezamiento posee además características diferenciales de la sucesión secundaria convencional (49):
a) Existe un subsidio de energía al sistema (fertilizantes, combustibles, agroquímicos).
b) Hay recurrencia y cierta periodicidad en las perturbaciones (labranzas, herbivoría).
c) El sistema produce información (modificaciones del ambiente térmico o lumínico) que es captada y almacenada por el banco de propágulos del suelo.
El banco de propágulos, semillas, frutos, rizomas, estolones, tubérculos y cualquier otra forma de propagación constituye la pieza clave del proceso de regeneración de la vegetación. El estudio y la comprensión del funcionamiento de las poblaciones espontáneas que recurrentemente aparecen luego de la remoción del suelo o de la instalación de un cultivo constituyen el motivo de este curso: como ya se ha dicho una arvense no constituye una clase botánica particular sino que es una población vegetal espontánea, que exhibe características propias para un sistema, en un determinado lugar y tiempo (35).
El tratamiento del problema de arvenses en los cultivos bajo una óptica de corto plazo y en gran medida ignorando las características que exhiben las poblaciones espontáneas, ha demorado no solo la realización de estudios ecológicos aplicados al agroecosistema sino la implementación de estrategias soportadas sobre la base de teoría ecológica disponible (50).
Resulta, por lo tanto, urgente no sólo la convergencia sino también la profundización de los estudios y la difusión de los conocimientos obtenidos en los agroecosistemas a los fines de optimizar los esfuerzos realizados y compatibilizarlos con los desafíos de la agricultura sustentable (14, 48).
LAS ARVENSES: POBLACIONES EXITOSAS EN EL AGROECOSISTEMA
Las arvenses son una forma especial de vegetación altamente exitosa en ambientes agrícolas, ya que son poblaciones vegetales que crecen en ambientes perturbados por el hombre sin haber sido sembradas. Desde un punto de vista ecológico las arvenses pueden ubicarse dentro de las pioneras de la sucesión secundaria. En el agroecosistema el impacto más crítico de las arvenses es el efecto negativo sobre las plantas cultivadas ejercido a través de la competencia por recursos limitados y las alelopatías. Trastornos en la recolección y el acondicionamiento de los granos y la disminución de la calidad del forraje constituyen perjuicios adicionales en muchos sistemas (26).
Desde el punto de vista evolutivo, implica generalmente la perdurabilidad o continuación de la línea genética a través del tiempo. De esta forma el éxito evolutivo está reflejado por el número de individuos, la capacidad reproductiva, el área y el rango de hábitats ocupados por la especie en cuestión. La comprensión acerca del concepto de éxito en el agroecosistema es necesaria para explorar la naturaleza del enmalezamiento y sentar las bases sólidas de manejo en los sistemas de producción (4).
DEFINICIÓN DE ÉXITO
Las arvenses más exitosas en los ecosistemas agrícolas son a menudo las que se consideran más problemáticas. El éxito puede medirse en este contexto, según la rapidez de colonización, la dificultad de su eliminación y el efecto negativo sobre la productividad de las especies cultivadas.
En general, la gran mayoría de las características de una planta que contribuye a una alta capacidad de colonización y competitividad se corresponde con las exhibidas por las arvenses, y en un grado mucho mayor que el exhibido por las especies cultivadas (9).
ESTRATEGIAS ADAPTATIVAS DE LAS ARVENSES
Una aproximación comúnmente utilizada por los ecólogos con el fin de comprender las relaciones entre los organismos y su ambiente es la de comparar sus estrategias adaptativas. Las poblaciones se adaptan al ambiente mediante el balance de asignación y la partición de recursos y energía entre los procesos fisiológicos y los componentes de la biomasa a lo largo del ciclo de vida (51).
En otras palabras, debe existir una solución de compromiso entre los recursos que se asignan para competir exitosamente en el presente (biomasa asignada a estructuras vegetativas) y aquellos que se destinan al futuro, es decir a la descendencia (biomasa asignada a semillas). De este concepto surge la idea de “Valor Reproductivo” una cuestión central en una población exitosa: el conjunto de características de la historia de vida que maximizan la supervivencia (lx) y la reproducción (Bx) configuran la adecuación (=fitness), en un ambiente particular. Las poblaciones de arvenses exhiben una adecuación elevada o máxima (20).
LA SELECCIÓN r-K
Una teoría ampliamente aceptada y que concierne con las estrategias de especialización evolutiva es la de la selección r-K. Mientras que los organismos r seleccionados están adaptados para la colonización y la reproducción en ambientes impredecibles, los organismos K seleccionados están adaptados para la persistencia y la reproducción en ambientes estables. Las poblaciones r seleccionadas ocupan los estadíos iniciales de la sucesión, en cambio las poblaciones K seleccionadas ocupan mayormente los estadios mas tardíos de la sucesión Si bien muchas de las poblaciones de arvenses exhiben características propias de la estrategia r, otras poblaciones pueden ubicarse dentro de K y en muchos casos las poblaciones exhiben características comunes a ambas ubicándose en un continuum entre r y K (7).
LA SELECCIÓN C.S Y R
Se sugirió una hipótesis alternativa en relación con las estrategias adaptativas y que define más claramente el rol del ambiente en determinar el éxito de la planta, donde se propuso que los factores externos que limitan la cantidad de biomasa que puede generarse en un ambiente dado pueden ser clasificados en dos categorías: el primero es el estrés y el segundo es el disturbio (52).
Si bien estos dos factores varían entre los hábitats, los casos extremos pueden combinarse de tal manera que resultan en tres tipos de hábitats y tres combinaciones posibles de estrategias evolutivas. Las competitivas maximizan la captura de recursos en condiciones altamente productivas y no perturbadas. Son abundantes durante las etapas tempranas e intermedias de la sucesión. Las tolerantes a estrés son plantas longevas adaptadas para vivir en condiciones de productividad limitada y a menudo ocupan los estadíos tardíos de la sucesión (52, 53).
Las ruderales, son a menudo hierbas de vida muy corta con alta producción de semillas y que ocupan los estadíos muy tempranos de la sucesión. Con el fin de incluir las estrategias secundarias que ocurren en hábitats intermedios en relación con los extremos que se han descrito, se propuso un modelo triangular: las tres estrategias extremas - competitivas - tolerantes a estrés (las ruderales se ubican en los vértices del triángulo) (36). De esta forma y basado en el estudio de una serie de especies, muchas arvenses herbáceas incluyendo anuales, bienales y algunas perennes así como muchos cultivos poseen características comunes a competidoras y ruderales, pudiendo ser clasificadas como competitivas-ruderales. Estas plantas están caracterizadas por un rápido crecimiento, rápida captura de recursos y alta producción de semillas (30, 54).
HABILIDAD COMPETITIVA
Las secciones precedentes han demostrado que las arvenses están adaptadas al ambiente agrícola a través de una serie de estrategias que maximizan un rápido crecimiento y una reproducción prolífica en hábitats perturbados. Estas características también contribuyen a su competitividad. La habilidad competitiva de las arvenses es una función compleja donde se combinan características que resultan en un rápido agotamiento de los recursos necesarios para el cultivo (4).
Ciertas características están recurrentemente asociadas con la habilidad competitiva: entre ellas se incluye a una gran cantidad de reservas acumuladas en órganos de propagación vegetativa o almacenaje que conduce a una rápida expansión del follaje, un sistema aéreo y subterráneo vigoroso y de rápido crecimiento que permite un rápido aprovechamiento de los recursos del ambiente y una expansión tanto lateral como horizontal que resulta en una muy alta densidad de vástagos y raíces.
La fenología es otra característica de la habilidad competitiva (55).
CARACTERÍSTICAS DE LAS ARVENSES ASOCIADAS AL ÉXITO ECOLÓGICO
Clasificación y características de las arvenses cosmopolitas
Si consideramos una cifra superior a 250 000 especies vegetales dentro de las fanerógamas, menos de 250 se consideran importantes en amplios sectores del planeta. De estas unas 70 se consideran “ las peores del mundo”. Estas 70 especies están distribuidas en 30 familias, pero casi los dos tercios están ubicadas en ocho familias y alrededor del 50 % están ubicadas en solo dos familias: Poáceas y Asteráceas.
El bajo número de especies consideradas arvenses en relación al total, ejemplifica la fenomenal presión selectiva ocurrida desde tiempos remotos (55).
De las 30 familias botánicas que contienen a las peores arvenses del mundo, cinco de ellas Poáceas, Solanáceas, Convolvuláceas, Euforbiáceas y Fabáceas también suministran el 75 % del alimento mundial. Esta observación implica que los cultivos y las arvenses comparten características taxonómicas y orígenes evolutivos comunes. Se describen a continuación características eco fisiológicas que exhiben las plantas que los seres humanos designamos como arvenses (35).
Características de la fase regenerativa
Los eventos que suceden en la fase regenerativa de las plantas (dispersión de las semillas, latencia, germinación y establecimiento de las plántulas) son particularmente importantes para las arvenses en ambientes agrícolas (33).
Dispersión de las semillas en el espacio
La dispersión es la diseminación de los propágulos de una planta desde un lugar a otro. La significancia para un organismo de este proceso depende de su ambiente. Tanto para las arvenses como para otras especies que conforman la sucesión, la supervivencia continuada depende del escape y el establecimiento a nuevas áreas que puedan ser más benignas para la supervivencia que la ocupada por su progenitor. Además del número de semillas que cada individuo produce, otros factores importantes que determinan el número de semillas que caen por unidad de área son la dispersabilidad de las semillas y la actividad de los agentes de dispersión (33, 35).
La dispersabilidad es una función de las características de las semillas las cuales incluyen apéndices sobre la semilla o el fruto. Muchas arvenses comunes poseen formas muy especializadas de dispersión mecánica, mientras que en otras la semilla es propulsada por diversos mecanismos de tensión que ocurren en el fruto. Ejemplos de estos mecanismos se encuentran en Oxalis corniculata L., Erodium sp. y Avena fatua (34).
La forma más común de dispersión es la pasiva, la cual se incrementa por la actividad de los agentes de dispersión como el viento, el agua, los animales y el hombre. En algunos casos los agentes de dispersión naturales constituyen el mecanismo principal (caso del viento y las semillas provistas de pappus en Carduus sp., muy livianas como en el caso de la parásita Orobanche sp. o transportadas por la planta entera, como es el caso de Salsola kali) las actividades del hombre constituyen el aporte más significativo en este aspecto. La siembra de semilla sin certificación de pureza, la falta de limpieza en las herramientas e implementos de labranza y cosecha, el traslado de hacienda y el agua de riego constituyen entre otras las principales vías de dispersión e invasión de arvenses en nuevas áreas. Las normas, las regulaciones y las leyes de orden sanitario implementadas por los gobiernos de todo el mundo apuntan a disminuir este efecto (9).
Latencia y germinación de las semillas
La mayoría de las arvenses exitosas poseen prolongada viabilidad y pronunciada latencia, permitiendo su supervivencia en condiciones inadecuadas para el crecimiento de las plantas y la persistencia por largos periodos en el suelo. La latencia puede ser definida como un estado en el cual las semillas o yemas viables no germinan aun cuando existen condiciones de temperatura, humedad y concentración de oxígeno adecuadas (28).
Se denomina germinación al proceso por el cual el embrión reasume el crecimiento, lo cual resulta en su emergencia a través de las cubiertas de la semilla o fruto. La latencia está considerada como el factor primario que contribuye a la presencia continuada de semillas de arvenses en los suelos agrícolas. Como contraste, la latencia ha sido eliminada por el mejoramiento genético en la mayoría de las especies cultivadas de tal manera que ocurra una germinación y una emergencia sincrónica (4).
Que una semilla se duerma o germine depende de una serie de los factores relacionados con la característica de la semilla y factores ambientales. Estos dos componentes interactúan de tal manera que la latencia y la germinación son procesos fuertemente sincronizados con los cambios ambientales.
La sincronización ambiental provee a las semillas de una mayor seguridad de supervivencia, dado que la germinación ocurre en el momento en que las condiciones ambientales son también favorables para el crecimiento (31).
La latencia es una característica muy variable y aún entre las semillas de la misma planta. Este polimorfismo somático asegura la supervivencia continuada de las especies por reducción del riesgo de pérdida total en el caso que las condiciones luego de la germinación sean desfavorables para el establecimiento. El polimorfismo es muy común en las familias Asteráceas, Brasicáceas, Chenopodiaceas y Poáceas (4).
Si bien la latencia ha sido estudiada en detalle en numerosas especies, la fisiología de este proceso no está totalmente comprendida. De forma general, se puede decir que la latencia y la germinación son procesos muy complejos e interrelacionados, que están controlados por factores bioquímicos, fisiológicos y estructurales (41).
El rol del ambiente en la regulación de la latencia de la semilla y su desbloqueo es cuando la germinación se extiende hasta el periodo de maduración cuando la semilla esta en desarrollo en la planta progenitora. Las semillas que están en estado de dormancia cuando se desprenden de la planta se designan con latencia innata o primaria. La latencia innata puede estar causada por una inmadurez o exceso de humedad en el embrión o un requerimiento de postmaduración conferido por un régimen especial de temperatura, luz o estratificación (35).
Las semillas que no germinan debido a la restricción impuesta por un factor ambiental se designan como quiescentes o con latencia impuesta. Las no dormitadas y que entran en latencia luego de la dispersión a menudo en respuesta a condiciones ambientales que inhiben la germinación se consideran con latencia inducida o secundaria. Las que se encuentran presentes en el perfil del suelo presentan ciclos de latencia /no latencia que promueven la germinación durante el periodo más favorable para el desarrollo de la plántula. Esta compleja estrategia puede ser ejemplificada con el comportamiento de Ambrosia artemisiifolia, una especie propia de los estadíos sucesionales tempranos (28).
Cuando las semillas caen al suelo se encuentran dormidas y germinan luego de la estratificación invernal siempre y cuando el suelo sea perturbado y las semillas sean llevadas a la cercanía de la superficie del suelo. Las semillas que no germinan, pueden inducirse a una latencia secundaria. Esta latencia puede ser desactivada por otro ciclo de estratificación (4).
La germinación de las arvenses y otras especies de la sucesión temprana a menudo requieren luz para germinar pero son inhibidas por la luz filtrada por la vegetación (rojo lejano) y promovidas por temperaturas fluctuantes. Estas condiciones se cumplen cerca de la superficie del suelo desnudo donde los niveles de irradiancia son altos y las amplitudes térmicas elevadas. La germinación de semillas de arvenses también se ve muy disminuida con la profundidad, donde los niveles de oxígeno son bajos, hay ausencia de la luz y las temperaturas menores (56).
La mayoría de las semillas de arvenses germinan mejor a profundidades de 0,5 a 2 cm. La adaptación a germinación cerca de la superficie asegura la disponibilidad de recursos y reduce la probabilidad de competencia con las especies de germinación y el establecimiento más tardío. De esta manera, la germinación de semillas de arvenses es promovida por las labranzas, que traen semillas ubicadas en la profundidad del suelo hacia la superficie (28).
Una práctica común de manejo es la de traer semillas del estrato profundo al superficial mediante labranzas, esperar la ocurrencia de lluvias en zonas de secano o irrigar el campo en zonas de riego antes de la siembra removiendo las plántulas de arvenses con nuevas labranzas o herbicidas. Al revés, en sistemas con mínima labranza, la ausencia de perturbación del perfil del suelo asegura que las semillas permanezcan en los estratos profundos, donde no se generan condiciones aptas para la germinación (35).
Germinación y establecimiento de plántulas
Cuando la latencia concluye, la secuencia normal de eventos conduce a la emergencia de la radícula, el crecimiento subterráneo de la raíz primaria, la emergencia del vástago por encima del suelo y finalmente al establecimiento de la plántula. El establecimiento concluye cuando la plántula es capaz de realizar un crecimiento independiente de las reservas de los cotiledones. La emergencia en la mayoría de las dicotiledóneas es epígeo, es decir que los cotiledones se elevan por encima de la superficie del suelo. En la emergencia hipogea los cotiledones permanecen debajo de la superficie del suelo. Luego de la emergencia, los cotiledones adquieren rápidamente su condición de órganos fotosintetizantes y el área foliar se agranda rápidamente (41).
Se ha propuesto el concepto de “Sitio seguro” y se define como la zona en la cuales se encuentran condiciones apropiadas para la germinación y la supervivencia. El sitio seguro provee entonces no solo estímulos para desbloquear la latencia y favorecer la germinación sino también ausencia de peligros como los depredadores, los competidores, los patógenos y las toxinas (33).
A los propósitos del manejo de arvenses, resulta imperativo distinguir a las plántulas provenientes de semillas de los brotes o expresiones vegetativas derivados de órganos de multiplicación vegetativa como rizomas, estolones o tubérculos. La plántula representa el estadío más vulnerable en el ciclo de vida de la planta y generalmente es el más fácil de ser destruido ya sea mecánicamente o con herbicidas (4).
Características de la fase establecida
La fase establecida de las plantas involucra numerosas funciones relacionadas con la captura de los recursos, el crecimiento y la reproducción. Los eventos de esta fase son los mayores determinantes del éxito competitivo y de la supervivencia en el agroecosistema. En términos de reproducción, las arvenses de los agroecosistemas tienen sistemas de intercambio genético variables y generalistas, producen abundante cantidad de semillas por un largo periodo de tiempo y se reproducen vigorosamente a partir de fragmentos si son perennes. Estas características contribuyen en forma significativa al éxito en ambientes perturbados (34).
Orden de emergencia y capital inicial
El éxito de una planta está asociada con la rapidez con que se inician los procesos de germinación y la velocidad con que se genera biomasa adicional. Está demostrado que las semillas que germinan primero y que poseen mayor capital inicial (embriones mayores o tejidos de acumulación y reservas más abundantes) poseen una clara ventaja sobre los individuos de germinación más tardía o con menor capital inicial. Como se explica más adelante, la ocupación del espacio y la captura de recursos limitantes constituyen la clave de una población exitosa en la fase de establecimiento (32).
Crecimiento y captura de recursos
En general, los individuos que crecen más rápido que sus vecinos utilizan una fracción desproporcionada de los recursos disponibles, en detrimento de aquellos. La Tasa de Crecimiento Relativo (TCR) ha sido usada como medio para integrar las características fisiológicas en un valor común que permita comparar diferentes especies (57).
Diferentes autores han determinado la tasa de crecimiento potencial para 132 especies. Las plantas anuales tienen con frecuencia valores de RGR mayores en relación con plantas leñosas. En términos de estrategia adaptativa, tanto las competitivas como las ruderales poseen RGR elevadas. Esta característica está altamente correlacionada con una explotación oportunista de los hábitats perturbados y también constituye una tendencia general entre las especies que conforman los estadíos sucesionales tempranos (58).
Un componente importante del crecimiento de la planta es la partición de fotosintatos hacia la generación de área foliar adicional. En un estudio realizado por diferentes autores se demostró que la partición del área foliar o la tasa de expansión de área foliar nueva estaba altamente correlacionada con un rápido crecimiento. Otras investigaciones realizadas demostraron que los parámetros de crecimiento relacionados con el tamaño de la plántula y el área foliar fueron los mejores predictores de la competitividad en mezclas de las arvenses. Resulta claro que la rápida producción de área foliar, que se traduce en un rápido desarrollo del canopeo, es crítica para el éxito tanto de arvenses como de cultivos en el ambiente agrícola (57).
Así como, en la fase regenerativa, los eventos de la fase establecida están muy ligados a los factores ambientales. Los recursos, incluyendo la luz, el agua, los nutrientes minerales, el CO2 y el O2 son factores consumibles del ambiente que pueden limitar el crecimiento cuando están ausentes o en déficit. Como contraste, si bien las condiciones ambientales como la temperatura, el pH y la densidad aparente del suelo no son consumidos, tienen una crucial influencia sobre los procesos que determinan el crecimiento de la plántula.
La capacidad diferencial de las plantas de extraer recursos o de producir respuestas distintas a las condiciones ambientales son los principales determinantes del éxito competitivo entre especies (59).
Los procesos de particular importancia para el éxito de las arvenses en ambientes agrícolas incluyen el uso eficiente del CO2, la rápida y eficiente absorción del agua a través del sistema radical, la alta eficiencia en el uso del agua y la plasticidad morfológica y fisiológica en relación con cambios ambientales (57).
Ciclos fotosintéticos
Basadas en sus ciclos fotosintéticos, las plantas pueden dividirse en tres grupos principales. Estos incluyen C3 (Ciclo de Calvin-Benson),C4 (Ciclo Hatch-Slack) y CAM (60).
Si bien en cada uno de estos tres grupos se pueden encontrar a varias arvenses, la vía C4 es la más representada por las arvenses principales. Otros estudios han demostrado que solo el 0,4 % de la flora posee esta vía adicional de incorporación de CO2. De las 76 arvenses más importantes del mundo el 42 % emplean la vía C4 y el 78 % de las 18 arvenses más ubicuas y agresivas son C4. Un estudio similar ha demostrado que 16 especies cultivadas tienen la vía C3 (61).
Las plantas que poseen la vía C4 generalmente tienen tasas de fotosíntesis neta más elevadas que las plantas C3. La enzima que inicia la fijación de CO2 [PEP (fosfonenolpiruvato carboxilasa)] tiene una afinidad más alta por el CO2 que el de la RuBP (ribulosadifosfato) carboxilasa, enzima que inicia la fijación de CO2 en el Ciclo C3. En la vía C4 el CO2 es concentrado en las células del mesófilo y pueden mantenerse altas tasas de fotosíntesis, aún cuando los estomas están prácticamente cerrados o cuando la concentración de CO2 en el aire es baja. Además, dada la alta concentración de CO2 interna, la fotorespiración disminuye substancialmente, lo cual contribuye para el logro de altas tasas de fotosíntesis neta. Debe puntualizarse finalmente que en las plantas C4 funcionan en realidad ambos ciclos acoplados, actuando el ciclo Hatch-Slack como reservorio de CO2 fijado (60).
La alta eficiencia de fotosíntesis esta a menudo correlacionada con un rápido crecimiento, el que puede conferir una habilidad competitiva superior. Esto llevó a clasificar a las plantas según un criterio de eficiencia o no eficiencia basado en sus características fotosintéticas (C3 = ineficientes, C4 = eficientes.
Investigaciones posteriores han restado generalidad a esta idea, dado que especies C3 son altamente competitivas. Es que otros factores concurren para determinar la habilidad competitiva de una planta. Por ejemplo, para la mayoría de las plantas C4 la temperatura óptima para la fotosíntesis y el crecimiento es más alto que para las plantas C3. En un hábitat de condiciones térmicas más templadas la ventaja de las especies C4 no es tal, no solo porque la tasa fotosintética total es menor sino también porque la vía acoplada C3 - C4 tiene un mayor requerimiento energético para funcionar.
En ambientes agrícolas caracterizados por estrés hídrico y elevadas temperaturas muy probablemente la mayoría de las arvenses anuales y perennes de verano van a pertenecer al grupo C4, mientras que las anuales de invierno serán a menudo especies C3 (60).
Absorción de agua
Además del suministro estacional del agua, las plantas pueden regular la disponibilidad de agua en cualquier ambiente mediante una serie de mecanismos y procesos. Estos son: el desarrollo, la estructura y la distribución de las raíces, la tolerancial a bajo potencial de agua en los tejidos, el control de pérdidas de agua por transpiración y la eficiencia en el uso del agua (WUE) (62).
En uno de los primeros trabajos relacionados con la estructura de las raíces, se demostró que 20 días luego de la emergencia la mayoría de las arvenses tienen sistemas radicales mayores y superficies de absorción más elevadas que los cereales con los cuales compiten. El desarrollo del sistema radical es el principal factor que contribuye a una explotación de la humedad del suelo y los nutrientes asociados, confiriendo así una competitividad superior (63).
El control fisiológico de la disponibilidad de agua por la planta puede ser expresado como Eficiencia en el uso del Agua (WUE) que se expresa por la cantidad de CO2 fijado o la cantidad de materia seca producida por unidad de agua perdida por transpiración. Ya en 1913 se informó que las plantas difieren significativamente en sus requerimientos de agua. Posteriormente se determinó que los valores más altos de WUE estaban en especies C4 mientras que las especies C3 tenían valores relativamente más bajos (41, 60).
Los valores más altos de WUE en las plantas C4 en relación a las C3 es debido al mecanismo de conservación de la vía C4. De esta manera, las plantas C4 son más productivas que las C3 cuando el agua es limitante (64).
La ventaja competitiva de una planta con alto valor de WUE debido a la vía C4 no es siempre tan evidente. Se ha demostrado que en algunas situaciones, una planta que mantiene estomas abiertos y altas tasas de transpiración bajo condiciones de estrés puede controlar la disponibilidad de agua en mayor medida que una planta con valor más alto de WUE y estomas más sensibles al estrés. Esto se encontró en el caso de mezclas de Chenopodium album L. (C3) con Amaranthus retroflexus L. (C4): valores altos de WUE de A. reflexus no confieren ventaja adaptativa sobre Ch. album (62).
De esta manera, el rápido desarrollo radical asociado con un uso excesivo del agua contribuye en forma significativa tanto al enmalezamiento como a la competitividad. La ventaja conferida por la vía fotosintética C4 y los valores asociados de WUE elevados que se han encontrado en muchas arvenses serían más importantes en ambientes más cálidos, secos, con elevada irradiancia y donde ocurren con gran frecuencia situaciones de estrés hídrico (57).
Una cuestión adicional se demuestra en el caso de Xanthium sp., una arvense muy frecuente en el cultivo de la soya y se ha comprobado que tanto en los estadíos vegetativos como reproductivos del cultivo, el nivel de potencial agua del xilema es siempre más negativo en Xanthium sp. Paralelamente, el proceso de mejoramiento genético del cultivo, que ha venido jerarquizando la producción de grano parece haber descuidado este aspecto; un análisis de los cultivares expuestos recientemente al mercado norteamericano permite comprobar que el potencial de agua del xilema y el rango de flexibilidad ante situaciones de estrés es menor que en cultivares lanzados dos o tres décadas atrás (60).
Plasticidad y respuestas ambientales
Los ambientes agrícolas en los cuales se encuentran las arvenses son a menudo sistemas muy perturbados en los cuales se presenta el suelo altamente expuesto, sin cobertura con extremos de temperatura en su superficie y fluctuaciones importantes tanto en la humedad como en los niveles de nutrientes. Estos cambios generalmente ocurren de manera impredecible (64).
Las arvenses mejor adaptadas a estas condiciones son tolerantes a estas condiciones variables de tal manera que pueden crecer y reproducirse en forma exitosa. A esta tolerancia a la variación ambiental se le designa plasticidad (45).
La plasticidad es la capacidad de un ajuste fenotípico, traducido en variaciones morfo-fisiológicas - condiciones cambiantes. Baker consideró que las arvenses poseen un genotipo de todo uso o multipropósito, lo que en otras palabras implica la existencia de un amplio rango de caracteres exhibidos por un limitado número de genotipos (22).
Existen ideas distintas sobre este punto. Más bien se puede considerar a las arvenses como poblaciones que contienen genotipos que responden a un alto grado de especialización y con un grado de ajuste muy fino a la oferta de señales que realiza el ecosistema bajo cultivo (55).
Lo que sí es incuestionable es que las especies cultivadas han estado sujetas a un intensivo proceso de mejoramiento y a través de el, han perdido flexibilidad morfológica y fisiológica para responder a variaciones ambientales (4).
Un ejemplo sobresaliente de la plasticidad de las plantas es la variación del número de sus partes o módulos. Otras variaciones plásticas incluyen el tiempo que transcurre entre diversas etapas del ciclo de crecimiento (emergencia - floración o floración - maduración), el tipo de semillas que producen (nivel de latencia), el tamaño relativo y la distribución de las raíces y las hojas. Las variaciones en el número, el tamaño y la distribución de las partes aéreas y subterráneas de la planta reflejan respuestas plásticas de la asignación y la partición de fotosintatos. Todos estos procesos confieren a una arvense la capacidad de rápida respuesta ante cambios bruscos en la distribución de recursos del hábitat (32).
Las arvenses también exhiben una respuesta plástica a la densidad. Una densidad creciente impone un estrés entre las plantas como consecuencia de la disminución de los recursos ambientales, una cuestión que se magnifica en la medida que las plantas crecen. A medida que las plantas crecen, la mortalidad denso dependiente y el consiguiente aumento de tamaño de los sobrevivientes - crecimiento plástico - conduce a una situación en donde sobreviven pocos individuos pero muy grandes. En términos generales se puede afirmar que este es un proceso de amplia ocurrencia y esta designado como Ley del Auto raleo o si nos referimos al modelo lineal: Ley de -3/2 (65).
Como consecuencia de los procesos mencionados, hay situaciones donde existen muchos individuos pero pequeños y otras donde existen pocos pero más grandes. La expresión de la biomasa vegetal por unidad de área conduce a otra generalización: en un amplio rango de densidades, la biomasa por unidad de área es constante (Ley del Rendimiento Final Constante) (33).
Floración y producción de semillas
El sistema de mejoramiento de las arvenses constituye uno de los componentes cruciales del éxito.
La mayoría de las arvenses exhiben un alto nivel de endocría ya sea a través de la autogamia (autopolinización) o mediante la agamospermia. No obstante, en todas ellas existe un nivel de alogamia, lo cual permite el intercambio de material genético con otras poblaciones. La alogamia es producida generalmente por el viento o visitantes generalistas de la flor más que por polinizadores específicos (36).
Este sistema combinado conduce a la producción de duplicados estables del genotipo padre, pero surgidos de recombinación y variabilidad genética amplia. Las ventajas de este sistema combinado son evidentes: el testeo de una amplia gama de genotipos a los efectos de que uno de ellos logre el éxito y el rápido avance posterior del genotipo más adecuado. Este esquema resulta aun más eficiente en las especies que poseen propagación vegetativa (S. halepense, por ejemplo) (66).
Las arvenses están caracterizadas por una producción de semilla abundante y continuada bajo condiciones ambientales diferentes .También la densidad juega un papel crucial en la producción de semillas por cada individuo: la fecundidad es un atributo fuertemente densodependiente.
Dos arvenses estudiadas en detalle en la Argentina, como Amaranthus quitensis y Datura ferox muestran claramente esta respuesta. En este último caso, esta característica poblacional, asociada al diseño de las cosechadoras, que provocan una redispersión de semillas de la arvense en el campo, permiten explicar la presencia de Datura ferox (chamico) en lotes de soya aun cuando la eficiencia de los herbicidas utilizados para su control es muy elevada (66).
CARACTERÍSTICAS ESPECIALES DE LAS ARVENSES PERENNES
La mayoría de los atributos de las arvenses previamente descritas se aplican a las arvenses perennes, como es el caso de especies muy prolíficas como C. rotundus. La reproducción vegetativa vigorosa y la regeneración a partir de fragmentos constituyen otra de las características mencionadas por Baker en su lista de atributos de la arvense ideal (33).
La significancia de la propagación vegetativa en la lista original es importante; el 61 % de las 18 peores arvenses exhibe alguna forma de propagación vegetativa. Como se ha explicado antes, la propagación vegetativa tiene los mismos efectos que la autopolinización o retrocruza, el resultado de ambos procesos es una rápida multiplicación de individuos con genotipos apropiados para el ambiente en cuestión (55).
El balance entre la propagación vegetativa y la reproducción por semillas en cualquier especie es altamente dependiente de las condiciones ambientales, incluyendo la densidad. En el caso de las especies que poseen ambos medios de multiplicación sexual y asexual, como en S. halepense, la combinación selectiva de los dos procesos exhibe una complementariedad muy eficiente: la reproducción sexual es significativa en la etapa de colonización o de reinstalación de la plaga en el campo y la propagación vegetativa multiplica muy rápidamente los genotipos más exitosos (31).
Las plantas que se reproducen vegetativamente poseen estructuras especializadas que pueden sobrevivir en temperaturas bajas, algún nivel de sequía u otra condición desfavorable. Cuando estas estructuras están localizadas por debajo de la superficie del suelo son muy persistentes y robustas. La función y significancia de las estructuras reproductivas vegetativas es doble: por un lado producen yemas o meristemas que pueden generar vástagos y raíces y por el otro sirven como tejidos de almacenaje y de reserva de carbohidratos (4).
A menudo, más de un tipo se encuentra en la misma planta. Estas estructuras incluyen a estolones, rizomas, tubérculos, turiones, bulbos, cormos, coronas y raíces gemíferas. C. rotundus, por ejemplo, posee rizomas, bulbos y tubérculos. De estos, el último contribuye significativamente a la propagación de la arvenses (28).
La habilidad competitiva de una arvense perenne esta significativamente aumentada por la cantidad de carbohidratos de reserva almacenados en las estructuras subterráneas. La rápida expansión del canopeo, factor principal que contribuye a la habilidad competitiva de muchas arvenses perennes, ocurre debido a la movilización de las reservas de energía y de materiales estructurales acumulados en los órganos de reserva originados en la estación de crecimiento anterior. Estas reservas preexistentes permiten una rápida emergencia y crecimiento de los jóvenes vástagos que poseen así una ventaja competitiva sobre aquellos que provienen de semillas, en general más pequeñas y con menor capital (33).
La cantidad de carbohidratos almacenados en los órganos subterráneos varían en relación con la época del año. En las épocas más desfavorables se produce una disminución debido al consumo por respiración de mantenimiento y en las etapas iniciales de la brotación como consecuencia del abastecimiento a los jóvenes vástagos en crecimiento. En las épocas favorables, el sistema aéreo produce fotosintatos que son trasladados a nuevas estructuras subterráneas (67).
El estudio de las variaciones en el nivel de biomasa aérea y subterránea y de carbohidratos resulta de sumo interés no solo para comprender los procesos de interacción arvense-cultivo sino también para aumentar la eficiencia en las prácticas de control. La búsqueda de predictores de fácil medición para estas variaciones constituyen líneas de trabajo de suma importancia práctica (35).
La propagación vegetativa puede estar acompañada por una fragmentación de órganos tanto aéreos como subterráneos (estolones y rizomas de Cynodon dactylon L.). La división de las estructuras vegetativas puede ocurrir espontáneamente debido a la muerte de los tejidos de conexión, la muerte de la planta madre o a una labranza, sin ninguna duda este último evento es el más eficiente para dispersar los propágulos de la arvense. Luego de la separación, las plantas que resultan de cada propágulo vegetativo son clones de la planta madre lográndose así una propagación horizontal del genotipo (52).
En algunos casos, los propágulos vegetativos exhiben latencia o se manifiestan fenómenos de correlación inhibitoria (dominancia apical), pero en la gran mayoría la ausencia de crecimiento de las yemas o meristemas está asociada a restricciones ambientales (sequía o temperatura). En el caso de la temperatura, existe un umbral de brotación variable para especies y aun ecotipos (68).
Cualquiera sea el mecanismo específico, las estructuras regenerativas de las plantas que almacenan reservas y que tienen la capacidad de propagarse vegetativamente son componentes significativos en los problemas de arvenses de los agroecosistemas.
MANEJO DE LAS ARVENSES
A pesar de que las arvenses reducen el rendimiento de un cultivo, su presencia contribuye a la estabilidad de los agroecosistemas. Por tal motivo se considera interesante aplicar métodos de análisis ecológicos a los sistemas agrícolas, con el fin de entender patrones como la distribución y la abundancia de las especies de arvenses, como lo han hecho (69–71). Los estudios ecológicos abren una nueva línea de investigación en el desarrollo de sistemas de manejo tendientes a buscar alternativas de control, que permitan obtener una buena producción agrícola sin menoscabo de los servicios ecológicos. El mejor uso y entendimiento de la diversidad de especies de arvense a nivel regional puede ser la clave para mantener la producción de los arrozales tradicionales de Tailandia (72).
Para desarrollar un apropiado manejo de arvenses en una agricultura sostenible, es necesario incluir investigaciones básicas de biología de las arvenses. Estos estudios, establecen la base fundamental para conocer mejor sus poblaciones, el crecimiento y el desarrollo, las interacciones con los aspectos bióticos y abióticos del agroecosistema, la fenología, la interferencia con los cultivos, la alelopatía y la competencia potencial de las adventicias (banco de semillas) (3).
Lo anterior ayudará a conformar adecuadamente las estrategias de manejo de las arvenses, especialmente para tomar la decisión propicia para llevar a cabo un determinado método de reducción de las arvenses y evitar efectos negativos sobre la calidad de la cosecha y el rendimiento. Por otro lado, la prevención es un factor que se considera poco en el manejo de las arvenses, este debe incluir el uso de semillas de cultivos no contaminados con semillas de arvenses, la limpieza del equipo de labranza y las herramientas, el manejo adecuado del agua para riego y de los animales para evitar la dispersión de las semillas en el campo y la prevención legal (72).
El manejo cultural de las arvenses, frecuentemente se menciona como un componente importante en el control de las arvenses; sin embargo, no se ejecuta plenamente. En una agricultura alternativa, las prácticas culturales deben ser combinadas para garantizar el éxito. Estas deben incluir: los sistemas de labranza, el uso de semillas de buena calidad o vigor, la época adecuada de siembra, la alta densidad, el momento oportuno de los métodos agronómicos, las coberturas muertas y vivas, la asociación, secuencia y rotación de cultivos (5).
Lamentablemente, hoy día, muchas de estas prácticas no se observan en los campos de los pequeños y medianos productores, consecuencia de la implementación de una agricultura moderna, que impulsó el uso irracional de químicos. No obstante; es necesario recuperarlas y conservarlas, si realmente se quiere desarrollar una agricultura con menos impactos en el ambiente (2).
Quizás las apreciaciones realizadas, sirvan como reflexión cuando se dice que una perspectiva histórica, sobre los métodos que realizan los campesinos son valiosas para aprender como hoy día podemos conservar mejor la energía, mantener los recursos y reducir el excesivo uso de químicos en la agricultura (6).
Algunos autores mencionan, que algunas de estas prácticas no solo ayudan a los agentes de biocontrol a actuar más eficientemente, sino también a conservar el suelo y hacer que el agroecosistema sea menos dependiente de fertilizantes, herbicidas y otros insumos agrícolas (49).
En la transición hacia una agricultura alternativa con pequeños agricultores, el uso de herbicidas puede jugar un papel importante en el manejo de las arvenses. Sin embargo, no debe ser el centro de un programa de arvenses, sino formar parte de un sistema que contemple otros métodos y orientarse al uso racional de las dosis, el tipo de herbicida adecuado a la situación y las rotaciones (72). Además, esta tecnología ofrece una posibilidad para la integración de programas de manejos más estables, para lo cual se debe considerar el carácter ecológico y ambiental en el empleo de herbicidas (73).
Indudablemente, si se desean implementar sistemas de manejo con un menor impacto en el ambiente, se requiere la incorporación de acciones conjuntas de la prevención, el manejo cultural, la limpieza mecánica, los fitogenéticos, los biológicos, los químicos, los aspectos políticos, los socioeconómicos y la capacitación de agricultores.
LAS BUENAZAS
Es la terminología que se viene utilizando en la agricultura orgánica para rescatar el nombre y la importancia de todas las plantas amigas y compañeras que crecen en medio de los cultivos, que comercialmente no representan ningún interés para la agricultura convencional y las cuales son mal denominadas como: “malezas, hierbas y plantas dañinas.
Las “buenazas” son las plantas que crecen naturalmente entre los espacios de los cultivos comerciales; éstas, como los demás vegetales contribuyen con la cobertura y la protección del suelo, con el reciclaje más eficiente de los nutrientes, con el aumento de la materia orgánica y el mejoramiento de la estructura del suelo (4).
En la agricultura orgánica es deseable que se mantengan naturalmente diferentes niveles de “buenazas” entre los cultivos, como una de las formas eficientes de recuperar y propiciar el seguimiento de la biodiversidad vegetal y crear condiciones para una mayor diversificación de la fauna entomológica, factor importante en la estabilidad del sistema (38). En este tipo de agricultura existen prácticas apropiadas al manejo de las “buenazas”, destacando las prácticas mecánicas, las rozadas y las podas manuales en los momentos más adecuados. También se utilizan plantas con efectos alelopáticos para reducir el incremento de las “buenazas”, principalmente a través de las fertilizaciones verdes y las coberturas muertas, entre otras (48).
UTILIDAD DE LAS ARVENSES
No solamente los aspectos mencionados anteriormente deben incluirse en el manejo de las arvenses en una agricultura sostenible, sino que es trascendental considerar la importancia y los beneficios que estos proporcionan en los agroecosistemas (Figura 1).
La óptica actual sobre las arvenses es consecuencia de la sucesión de las plantas en la agricultura moderna, producto de la perturbación del ambiente que favorece la especialidad de las arvenses competitivas. Aunque las adventicias interfieren con el plan de producción agrícola, algunas especies constituyen importantes componentes biológicos de los agroecosistemas, por lo que se les puede considerar elementos útiles en sistemas de uso de tierra (4).
Las arvenses interactúan ecológicamente con los otros subsistemas del agroecosistema y tienen mucha importancia contra la erosión y para la conservación del suelo, la formación de materia orgánica, la fijación del nitrógeno en el suelo, la preservación de los insectos beneficiosos y de la vida silvestre (74, 75).
Ciertas arvenses deben considerarse como componentes importantes de los agroecosistemas, debido a que pueden afectar en forma positiva la biología y la dinámica de insectos beneficiosos. Las arvenses ofrecen muchos requisitos de importancia a los enemigos naturales como presas huéspedes alternativas de polen o néctar, además de micro hábitats que no se encuentran presentes en los monocultivos libres de arvenses (9).
En Mesoamérica existe una gran diversidad de especies con valor antropocéntrico, de las cuales los agricultores poseen un amplio conocimiento biológico de las plantas que forman parte del medio ecológico (76). Este conocimiento conduce a la selección de numerosas especies para la alimentación humana, los animales domésticos y en el control de otras plagas de las especies cultivadas (5). Asimismo, las arvenses tienen gran interés en el ámbito científico, para el aumento de la diversidad genética y en trabajos de entomología y fitopatología como fuentes de sustancias repelentes o biocidas (77).
El pequeño y mediano agricultor de Centroamérica, se caracteriza por el uso correcto de los recursos limitados de su sistema agrícola, esto incluye también el manejo de las arvenses. Contrario al enfoque anterior, la agricultura intensiva y excesiva en el aprovechamiento de los insumos tiene como objetivo alcanzar una alta producción de las especies cultivadas, esto significa la eliminación total de las arvenses, lo que provoca la pérdida de especies con valor científico y de utilidad potencial para el hombre (5, 76).
En Guatemala, los campesinos reconocen la competitividad de las arvenses en la parte inicial de los cultivos, los cuales utilizan ampliamente en diferentes regiones antropocéntricas. Realizan limpias de las arvenses dentro del periodo crítico, para que después de este crezcan y sean utilizadas, de no ser así, desarrollan deshierbes selectivos y dejan crecer a la par de los cultivos aquellas arvenses que tienen para ellos un valor en la alimentación humana, la animal o el uso medicinal (3).
En México alrededor de 40 especies asociadas con los campos de maíz, son consumidas como verdura por los campesinos y algunas de estas especies se les permite diseminar sus semillas para aumentar su crecimiento (6).
Definitivamente, las arvenses juegan un papel importante en los campos de una gran mayoría de agricultores tradicionales del trópico, quienes hacen un uso intensivo y variado de estas. Algunas arvenses que se utilizan para la alimentación humana son Bidens pilosa L., Solanum spp. y Portulaca oleraceae L. Para la alimentación de animales domésticos Melampodium divaricatum (Rich) DC., B. pilosa, Cynodon dactylon (L) Pers., Cynodon nlemfuensis Vandeiyst. y Setaria spp. (5).
Para uso medicinal, emplean como colirio a Argemone mexicana L., Asclepias glaucescens L., es aplicada como desinfectante, Cleome viscosa L., para baños posterior al parto, Chenopodium ambrosioides L., como antihelmíntico, Rumex crispus L., para laxante. Otras especies tienen valor ornamental como Commelina spp. y el marabú Cailliea glomerata Forsk.; además, poseen una buena protección de los terrenos contra la erosión. Para la alimentación de aves se usan semillas de A. mexicana y de Rottboellia cochichinensis (Lour) W.D.Clayton.
También las arvenses tienen mucha importancia como reservorio genético de las plantas cultivadas. El tomate es un ejemplo clásico de una planta cultivada que se ha mejorado por hibridación con especies silvestres emparentadas (78).
Otro aspecto importante de las arvenses en un agroecosistema es la acción que tienen sobre algunos insectos como Eleusine indica (L.) Gaertner., que regula a Empoasca kraemeri (Ross & More) por repelencia química o disfraz, S. halepense, disminuye a Eotetranychus cuillamettei, debido a que aumenta los ácaros depredadores (9). Amaranthus viridis L., Boerhavia erecta L. y Cucumis foetidia L., son huéspedes alternos de áfidos y frenan así considerablemente la trasmisión de la virosis (79).
Asimismo, las especies que pertenecen a la familia Fabaceae son muy importantes, por ser portadoras de nódulos de Rhizobium las cuales enriquecen el suelo con nitrógeno. En Costa Rica, hay estudios que indican, la reducción del daño que producen los insectos plagas, principalmente de Diabrotica spp., sobre leguminosas cuando se dejó Amaranthus spinosus L., en la entre línea. Mientras que en Nicaragua, el gusano cogollero S. frugiperda , causó un mayor daño en las plantas jóvenes de maíz en un hábitat libre de arvenses, el mismo se redujo cuando existió una cobertura de arvenses, compuesta por una mayor abundancia de monocotiledóneas como C. rotundus, S. halepense, Panicum spp., Rottboellia cochichinensis y Setaria spp. (23, 80).
La diversidad de un agroecosistema, beneficia la variedad de insectos parásitos depredadores. La práctica de diversificación, debe realizarse a través de la utilización de diferentes cultivos y el empleo de poblaciones de arvenses dentro del cultivo o en los bordes, para que actúen como repelentes de insectos o cultivos trampas (81).
Igualmente, las arvenses son importantes como fuentes de extractos que se usan para la elaboración de plaguicidas. B. pilosa afecta las poblaciones de nemátodos, Ch. ambrosioides , actúa como repelente de insectos. Se plantea que las sustancias extraídas de Tithonia tubaeformis (Miller) Blake y T. diversifolia son venenosas contra la polilla de la col y las de Lantana camara L. actúa como repelente sobre esta misma plaga (79).
Finalmente C. rotundus es considerada como una de las arvenses más indeseables en el trópico, aun así, esta especie por su sistema radicular que desarrolla puede jugar un papel importante en áreas susceptibles a la erosión. Además, es posible que las sustancias alelopáticas que producen, tengan un potencial como plaguicida (80).
El enfoque expuesto anteriormente sobre las arvenses, indican su rol ecológico en los agroecosistemas, sus usos múltiples desde el punto de vista biológico y el valor utilitario que le dan los pequeños productores de Mesoamérica. Esto, llama a la reflexión sobre las cualidades de las arvenses, especialmente para aquellos científicos y técnicos que quieran desarrollar una agricultura sostenible.
Queda la esperanza de que los grandes agricultores y los campesinos asimismo como los ecólogos no estén riñendo sobre la palabra arvense o planta beneficiosa sino tratan de buscar caminos y fuentes para asegurar altos y estables rendimientos también en una agricultura sostenible en el trópico.
DIFERENTES APORTES QUE REALIZAN LAS ARVENSES EN EL AGROECOSISTEMA
Aportes al equilibrio edáfico
Las prácticas para mejorar la fertilidad de los suelos pueden impactar directamente la susceptibilidad fisiológica del cultivo a los insectos plaga, ya sea al afectar la resistencia al ataque de las plantas individuales o al alterar la aceptabilidad de algunas plantas hacia ciertos herbívoros (2).
Varias investigaciones demuestran que la capacidad de un cultivo de resistir o tolerar el ataque de los insectos plaga y las enfermedades está ligada a las propiedades físicas, químicas y particularmente biológicas del suelo. Suelos con alto contenido de materia orgánica y una alta actividad biológica, generalmente exhiben buena fertilidad, así como cadenas tróficas complejas y organismos benéficos abundantes que previenen la infección. Por otro lado, las prácticas agrícolas que causan desbalances nutricionales, como la aplicación excesiva de fertilizantes nitrogenados sintéticos, bajan la resistencia de las plantas a las plagas (81).
Las plantas funcionan en un ambiente complejo multitrófico, donde generalmente la flora y la fauna del suelo y los organismos de arriba del suelo (cultivos, insectos, etc.) interactúan en redes tróficas complejas, con una serie de interacciones que pueden favorecer o desfavorecer la menor incidencia de plagas (Figura 2). Las comunidades de arriba del suelo se ven afectadas directa e indirectamente por interacciones con los organismos de la red trófica del suelo (82).
Las actividades de alimentación de los descomponedores o detritívoros (básicamente bacterias y hongos) en la red trófica estimulan el movimiento de nutrientes, la adición de nutrientes por las plantas y el funcionamiento de estas, indirectamente, influyen sobre los insectos que se alimentan de los cultivos (83).
APORTES AL EQUILIBRIO DE LA ENTOMOFAUNA BENÉFICA
La tolerancia de las arvenses en los campos cultivados y sus alrededores constituyen un reto para los agricultores ya que generalmente estas plantas se consideren como arvenses y por tanto se ha establecido que deben eliminarse; pero se ha demostrado que ellas no siempre son perjudiciales (competencia, interferencia, reservorio de plagas) a los cultivos, sino que pueden contribuir a la conservación del suelo, la alimentación y ser refugio de artrópodos benéficos entre otros. Esto significa que hay que manejarlas con mucho cuidado para favorecer los efectos beneficiosos y reducir los perjudiciales (75).
La interacción entre las arvenses y las plagas asociadas debe ser objeto de correcta comprensión, para el mejor desarrollo de las prácticas de manejo integrado de plagas (MIP). A veces es aconsejable dejar una pequeña población de ciertas especies de arvenses, con el propósito de garantizar el desarrollo de depredadores importantes de insectos plaga (26).
La práctica ha demostrado que el control de arvenses suele reducir la incidencia de plagas. A pesar de que los enemigos naturales varían ampliamente en su respuesta a la distribución, densidad y dispersión de los cultivos, la evidencia señala que ciertos atributos estructurales del agroecosistema (la diversidad vegetal, los niveles de insumos, etc.) influyen marcadamente en la dinámica y la diversidad de depredadores y parasitoides (84).
La mayoría de estos atributos se relacionan con la biodiversidad y están sujetos al manejo (asociaciones y rotaciones de cultivos, presencia de arvenses en floración, diversidad genética, etc.). Basándose en la información disponible, la biodiversidad de enemigos naturales y su efectividad, se puede incrementar en los agroecosistemas de las formas antes mencionadas (7).
APORTES A LA FERTILIDAD DEL SUELO
En la naturaleza no existen “malas hierbas”, pero si plantas “adventicias e invasoras” que deben percibirse como indicadores ecológicos de gran utilidad para entender el estado de las calidades físicas, químicas y biológicas de los suelos debido a que estas favorecen la toma de elementos minerales por la planta, mejoran las propiedades físicas, químicas y las biológicas del suelo; además, aportan sustancias estimuladoras del crecimiento para la planta (83).
Las arvenses juegan un rol importante en la relación suelo-arvense, ya que mediante la acción ecológica - fisiológica de las arvenses, pueden mostrarse como indicadoras de las propiedades del suelo por diferentes elementos ya sea el fósforo, el potasio, el nitrógeno o el humus (85).
La vegetación herbácea espontánea en investigaciones realizadas permitió recopilar información con respecto al suelo donde aparecía. Por ejemplo, un suelo dominado por gramíneas estoloníferas (que disponen de tallos o estolones a lo largo de la superficie del suelo, raíces en los nodos y producen retoños nuevos), como Digitaria sanguinalis (L.) M. Scop. (pasto cuaresma o gramilla), presentaba una estructura física deficiente, o sea, no era un suelo suelto y por ello, probablemente la planta cultivada gastaba mucha energía para establecerse, pudiendo presentar deficiencia de nutrientes (Tabla I). Del mismo modo que las arvenses, las plagas indicaban el origen de las dificultades que las plantas estaban teniendo, como por ejemplo, la falta de nutrientes (Tabla II). Sin embargo, esos nutrientes podían estar presentes en el suelo más no estaban siendo aprovechados por las plantas, como es el caso de la pudrición apical en el tomate que ocurre debido a la falta de calcio en los períodos en que el suelo está demasiado seco y no necesariamente por carencia de este mineral en el ambiente (86).
Las coberturas vegetales actualmente están siendo incluidas en los sistemas agrícolas con el motivo de incrementar la fertilidad del suelo y el funcionamiento del cultivo a largo plazo, a partir del control de la erosión, el incremento de la materia orgánica y el mejoramiento de las propiedades físicas del suelo, a corto plazo, afectando el balance de radiación, la temperatura y la humedad del suelo, la disponibilidad de nutrientes, la relación escorrentía- infiltración y el establecimiento del cultivo (89).
Otros aportes (retención de la humedad, evitar la erosión del suelo)
En el trópico, la utilización de las arvenses como protección del suelo resulta vital para disminuir el efecto de factores importantes de degradación, como la insolación y el impacto directo de la lluvia.
El efecto de las arvenses como cobertura vegetal del suelo sobre la erosión puede ser dividido en tres tipos, el efecto de tipo uno, es relativo a la cobertura vegetal ofrecida por el dosel, el efecto de tipo dos, referente a la cobertura vegetal en contacto directo con la superficie del suelo y el efecto de tipo tres, es relativo a la incorporación de residuos vegetales al suelo en función de su manejo (90).
El drenaje y la labranza agrícola, aceleran las pérdidas por percolación profunda; la remoción de los cultivos aumenta la cantidad de lluvia que llega al suelo y reduce la evapotranspiración; los cambios en la estructura del suelo debido al control de residuos de labranza, la rotación de cultivos o el uso de abonos afecta la tasa de percolación y el flujo lateral. Uno de los controles principales de la acumulación de humedad en el suelo es ejercido por la cobertura vegetal, puesto que influye en los insumos y en las pérdidas ejercidas hacia y desde la humedad del suelo. Por ejemplo, el dejar el follaje cortado de las arvenses como mulch, reduce las pérdidas de agua provenientes de la evapotranspiración y aumenta los contenidos de humedad del suelo (45).
Un suelo capaz de soportar una producción vegetal abundante es una mezcla de sustancias inorgánicas procedentes del sustrato original, la materia orgánica producida por las plantas y una vida intensa que transforma la materia orgánica, poniendo a disposición de las plantas una buena parte de los nutrientes que necesitan, asociándose con ellas para facilitar la toma de nutrientes, reduciendo la pérdida de éstos en el suelo y creando condiciones para la aireación, la penetración y la retención del agua en el suelo (85).
CONSIDERACIONES GENERALES
Por todo lo explicado anteriormente, se hace necesario seguir proporcionando investigaciones, para demostrar que las arvenses, que siempre han sido consideradas como perjudiciales por su interferencia en los cultivos económicos estableciendo una marcada competencia con ellos por la luz, el agua, los nutrientes, el CO2 y el espacio físico, o por la producción de sustancias nocivas para el cultivo, juegan también un papel beneficioso dentro del agroecosistema.
Ello significa que es necesario establecer normas de convivencia, mediante el manejo adecuado de las arvenses en convivencia interespecífica con los cultivos, teniendo en cuenta el período crítico de competencia (interferencia), pues está demostrado que la presencia de diferentes especies de arvenses en los cultivos, mantiene la composición de la entomofauna benéfica y, a su vez, tienen mayores posibilidades de encontrar presas alternativas, abrigo, sitios para reproducción y refugios para dormancia ya que el agroecosistema se encuentra más complejo. También manejando las arvenses, se puede determinar los principios ecológicos generales que regulan la dinámica de las arvenses y las interacciones de estas en los agroecosistemas. A través de nuevas investigaciones, se podrá profundizar sobre las combinaciones específicas: cultivo-arvenses en los agroecosistemas locales y para desarrollar lineamientos flexibles en el diseño de sistemas agrícolas.
A partir de que se conozca que la diversidad vegetal favorece de manera frecuencial los mecanismos homeostáticos de control de plagas en los cultivos, se controla la erosión del suelo entre otras, es de presumir que la diversidad insectil sea mayor y las densidades de fitófagos sean menores y más estables en los cultivos que presenten distintas especies de arvenses, que en el cultivo sin ninguna diversidad vegetal y que, a su vez, las variedades ejerzan cierta influencia sobre estos indicadores, por lo que dichas prácticas podrían ser utilizadas para la regulación de comunidades de insectos nocivos en diferentes cultivos y así manejar el agroecosistema agroecológicamente, sin tener que acudir a los productos químicos.
BIBLIOGRAFÍA
1. Altieri, M. Á. y Nicholls, C. I. Biodiversidad y manejo de plagas en agroecosistemas [en línea]. Ed. Icaria, 2007, 252 p., ISBN 978-84-7426-764-8, [Consultado: 1 de julio de 2016], Disponible en: <https://www.google.com/search?tbo= p&tbm=bks&q=isbn:8474267641>.
2. Altieri, M. A. y Nicholls, C. I. Agroecología y resiliencia al cambio climático: principios y consideraciones metodológicas. Agroecología, vol. 8, no. 1, 2013, pp. 7-20, ISSN 1989-4686.
3. Altieri, M. y Nicholls, C. Agroecología: teoría y práctica para una agricultura sustentable [en línea]. (ser. Textos básicos para la formación ambiental), vol. 4, Ed. Programa de las Naciones Unidas para el Medio Ambiente - Red de Formación Ambiental para América Latina y el Caribe, 2000, 250 p., ISBN 978-968-7913-07-0, [Consultado: 1 de julio de 2016], Disponible en: <https://books.google.com.cu/books/about/Agroecolog%C3%ADa.html?id=to-yPQAACAAJ&redir_esc=y>.
4. Blanco, Y. y Leyva, A. Las arvenses en el agroecosistema y sus beneficios agroecológicos como hospederas de enemigos naturales. Cultivos Tropicales, vol. 28, no. 2, 2007, pp. 21–28, ISSN 1819-4087.
5. Gamboa, W. y Pohlan, J. La importancia de las malezas en una agricultura sostenible del trópico. Journal of Agriculture in the Tropics and Subtropics, vol. 98, no. 1, 1 de abril de 1997, pp. 117-123, ISSN 0041-3186.
6. Thurston, H. D. Sustainable Practices for Plant Disease Management in Traditional Farming Systems [en línea]. Ed. Westview Press, septiembre de 1991, Boulder, New Delhi, 280 p., ISBN 978-0-8133-8363-7, [Consultado: 1 de julio de 2016], Disponible en: <https://www.amazon.com/Sustainable-Practices-Disease-Management-Traditional/dp/0813383633>.
7. Zamora, R.; García, F. P. y Gómez, A. L. Las interacciones planta-planta y planta animal en el contexto de la sucesión ecológica [en línea]. En: ed. Valladares F., Ecología del bosque mediterráneo en un mundo cambiante, (ser. Naturaleza y parques nacionales Serie técnica), Ed. Organismo Autónomo Parques Nacionales, Madrid, 2004, pp. 371-393, ISBN 978-84-8014-552-7, OCLC: 179982412, [Consultado: 1 de julio de 2016], Disponible en: <http://www.gbv.de/dms/goettingen/51086516X.pdf>.
8. Altieri, M. A. y Whitcomb, W. H. “The potential use of weeds in the manipulation of beneficial insects (agents in natural crop pest control)”. HortScience, vol. 14, 1979, pp. 12-18, ISSN 0018-5345.
9. Altieri, M. A. y Letourneau, D. K. “Vegetation management and biological control in agroecosystems”. Crop Protection, vol. 1, no. 4, diciembre de 1982, pp. 405-430, ISSN 02612194, DOI 10.1016/0261-2194(82)90023-0.
10. Van Emden, H. F. y Dabrowski, Z. T. “Biodiversity and habitat modification in pest management”. International Journal of Tropical Insect Science, vol. 15, no. 6, diciembre de 1994, pp. 605-620, ISSN 1742-7584, 1742-7592, DOI 10.1017/S174275840001691X.
11. Matienzo, B. Y.; Rijo, C. E.; Milán, V. O.; Torres, N. N.; Larrinaga, L. Y.; Romero, C. N. y Plá, del C. D. Diversidad de insectos benéficos asociados a Morinda citrifolia L.”. Fitosanidad, vol. 11, no. 1, 23 de noviembre de 2012, pp. 25-28, ISSN 1818-1686.
12. Landis, D. A.; Wratten, S. D. y Gurr, G. M. “Habitat Management to Conserve Natural Enemies of Arthropod Pests in Agriculture”. Annual Review of Entomology, vol. 45, no. 1, 2000, pp. 175-201, ISSN 0066-4170, DOI 10.1146/annurev.ento.45.1.175, PMID: 10761575.
13. Asteraki, E. J.; Aina, B.; Ings, T. C. y Manley, W. J. “Factors influencing the plant and invertebrate diversity of arable field margins”. Agriculture, Ecosystems & Environment, vol. 102, no. 2, abril de 2004, pp. 219-231, ISSN 01678809, DOI 10.1016/j.agee.2003.07.003.
14. Guzmán, C. G. y Morales, H. J. Agroecología y agricultura ecológica. Aportes y sinergias para incrementar la sustentabilidad agraria. Agroecología, vol. 6, 2011, pp. 55-62, ISSN 1989-4686.
15. Vázquez, L. Manejo plagas agricultura ecológica. Boletín Fitosanitario, vol. 15, no. 1, 2010, p. 11, ISSN 1816-8604.
16. Gabriel, D.; Roschewitz, I.; Tscharntke, T. y Thies, C. “Beta diversity at different spatial scales: plant communities in organic and conventional agriculture”. Ecological Applications, vol. 16, no. 5, octubre de 2006, pp. 2011-2021, ISSN 1051-0761, DOI 10.1890/1051-0761 (2006)016[2011: BDADSS]2.0.CO;2.
17. Pino, M. de los A. Diversidad agrícola de especies de frutales en el agroecosistema campesino de la Comunidad Las Caobas, Gibara, Holguín. Cultivos Tropicales, vol. 29, no. 2, junio de 2008, pp. 5-10, ISSN 0258-5936.
18. Návar, C. J. de J. y González, E. S. Allometric equations for tree species and carbon stocks for forests of northwestern Mexico. Forest Ecology and Management, vol. 257, no. 2, enero de 2009, pp. 427-434, ISSN 03781127, DOI 10.1016/j.foreco. 2008.09.028.
19. Nicholls, C. Bases agroecológicas para diseñar e implementar una estrategia de manejo de hábitat para control biológico de plagas. Agroecología, vol. 1, no. 0, 22 de abril de 2008, pp. 37-48, ISSN 1989-4686.
20. Altieri, M. Desiertos verdes: monocultivos y sus impactos sobre la biodiversidad [en línea]. En: Courtis C., Monocultivos: en la frontera de los derechos humanos, Ed. FIAN Internacional, FIAN Suecia, HIC-AL, SAL, 2009, pp. 55-62, ISBN 978-607-95101-2-1, [Consultado: 30 de agosto de 2016], Disponible en: <http://es.slideshare.net/MercedesGmez2/azucar-roja>.
21. Jarvis, D.; Padoch, C. y Cooper, H. D. Manejo de la biodiversidad en los ecosistemas agrícolas. (trad. Walter A.), Ed. Bioversity Internacional, 2011, Roma, 503 p., ISBN 978-92-9043-823-6, OCLC: 812025908.
22. Ranaboldo, C. y Venegas, F. C. Escalondando la Agroecologi ́a: Procesos Y Aprendizajes de Cuatro Experiencias en Chile, Cuba, Honduras Y Peru ́ [en línea]. 1.a ed., Ed. Plaza y Valdés-Centro Internacional de Investigaciones para el Desarrollo, 2007, México-Ottawa, 186 p., ISBN 978-1-55250-341-6, Google-Books-ID: 7M8tcCZHDiUC, [Consultado: 30 de agosto de 2016], Disponible en: <https://books.google.com.cu/books?id=7M8tcCZHDiUC>.
23. Altieri, M. A.; Liebman, M.; Hecht, S. B.; Magdoff, F. y Sikor, T. O. Agroecología : bases científicas para una agricultura sustentable. Ed. Nordan-Comunidad, 1999, Montevideo, Uruguay, 315 p., ISBN 9974-42-052-0.
24. Altieri, M. A. y Toledo, V. M. “The agroecological revolution in Latin America: rescuing nature, ensuring food sovereignty and empowering peasants”. Journal of Peasant Studies, vol. 38, no. 3, julio de 2011, pp. 587-612, ISSN 0306-6150, 1743-9361, DOI 10.1080/03066150.2011.582947.
25. Andow, D. A. “Yield Loss to Arthropods in Vegetationally Diverse Agroecosystems”. Environmental Entomology, vol. 20, no. 5, 1 de octubre de 1991, pp. 1228-1235, ISSN 0046-225X, 1938-2936, DOI 10.1093/ee/20.5.1228.
26. Patro, L. R. Biodiversity conservation and management. Ed. Discovery Pub. House, 2010, New Delhi, 249 p., ISBN 978-81-8356-600-1, OCLC: 495614325.
27. Afifi, M. y Swanton, C. “Early Physiological Mechanisms of Weed Competition”. Weed Science, vol. 60, no. 4, octubre de 2012, pp. 542-551, ISSN 0043-1745, 1550-2759, DOI 10.1614/WS-D-12-00013.1.
28. Roschewitz, I.; Gabriel, D.; Tscharntke, T. y Thies, C. “The effects of landscape complexity on arable weed species diversity in organic and conventional farming: Landscape complexity and weed species diversity”. Journal of Applied Ecology, vol. 42, no. 5, 5 de septiembre de 2005, pp. 873-882, ISSN 0021-8901, 1365-2664, DOI 10.1111/j.1365-2664.2005.01072.x.
29. Fuentes, D. C. L. y Romero, C. E. Una visión del problema de las malezas en Colombia. Agronomía Colombiana, vol. 8, no. 2, 1991, pp. 364-378, ISSN 2357-3732.
30. Cerrudo, D.; Page, E. R.; Tollenaar, M.; Stewart, G. y Swanton, C. J. “Mechanisms of Yield Loss in Maize Caused by Weed Competition”. Weed Science, vol. 60, no. 2, abril de 2012, pp. 225-232, ISSN 0043-1745, 1550-2759, DOI 10.1614/WS-D-11-00127.1.
31. Powell, W.; Dean, G. J. y Dewar, A. “The influence of weeds on polyphagous arthropod predators in winter wheat”. Crop Protection, vol. 4, no. 3, septiembre de 1985, pp. 298-312, ISSN 02612194, DOI 10.1016/0261-2194(85)90032-8.
32. Barroso, J.; Miller, Z. J.; Lehnhoff, E. A.; Hatfield, P. G. y Menalled, F. D. “Impacts of cropping system and management practices on the assembly of weed communities”. Weed Research, vol. 55, no. 4, agosto de 2015, pp. 426-435, ISSN 0043-1737, DOI 10.1111/wre.12155.
33. Labrada, R.; Paredes, E. y Morales, R. Lucha química contra malezas en fríjol y soya. III. Susceptibilidad varietal. Ciencia y Técnica en la Agricultura, Protección de Plantas, vol. 9, 1987, pp. 33–47, ISSN 1010-1578.
34. Labrada, R.; Caseley, J. C.; Parker, C. y Organización de las Naciones Unidas para la Agricultura y la Alimentación. Manejo de malezas para países en desarrollo. Ed. Organización de las Naciones Unidas para la Agricultura y la Alimentación, 1996, Roma, 403 p., ISBN 978-92-5-303427-7, OCLC: 133095251.
35. Grime, J. P.; García, F. C. A. y Cervantes, R. M. Estrategias de adaptación de las plantas y procesos que controlan la vegetación [en línea]. Ed. Limusa, 1982, México, 291 p., ISBN 978-968-18-1360-4, OCLC: 893495388, [Consultado: 1 de julio de 2016], Disponible en: <https://www.worldcat. org/search?q=ti%3A Estrategias+de+ adaptaci%C3%B3n +de+las+plantas+y+procesos +que+ controlan +la +vegetaci%C3 %B3n&fq= yr%3A1982..1982+ %3E&qt= advanced&dblist =638>.
36. Maítre, A.; Bentley, J. W. y Fischler, M. ¿Qué es más urgente, el manejo integrado de plagas o el manejo sostenible de suelos?. Revista Ceiba, vol. 44, no. 1, 22 de octubre de 2012, pp. 45-59, ISSN 0008-8692, 2225-6687.
37. Blanco, V. Y.; Leyva, G. Á. y Castro, L. I. Determinación del período crítico de competencia de arvenses en el cultivo del maíz (Zea mays, L.)”. Cultivos Tropicales, vol. 35, no. 3, septiembre de 2014, pp. 62-69, ISSN 0258-5936.
38. Chávez-Servia, J. L.; Tuxill, J. y Jarvis, D. I. Manejo de la diversidad de los cultivos en los agroecosistemas tradicionales [en línea]. Ed. Instituto Internacional de Recursos Fitogenéticos, 2004, Cali, Colombia, 264 p., ISBN 978-92-9043-658-4, [Consultado: 30 de agosto de 2016], Disponible en: <https://www.google.com/search?tbo= p&tbm=bks&q= isbn:9290436581>.
39. Vázquez, L.; Matienzo, Y.; Veitia, M. y Alfonso, J. Conservación y manejo de enemigos naturales de insectos fitófagos en los sistemas agrícolas de Cuba. Ed. CIDISAV, 2008, La Habana, Cuba, ISBN 978-959-7194-17-0.
40. Blanco, Y. y Leyva, Á. Abundancia y diversidad de especies de arvenses en el cultivo de maíz (Zea mays, L.) precedido de un barbecho transitorio después de la papa (Solanum tuberosum L.)”. Cultivos Tropicales, vol. 31, no. 2, junio de 2010, pp. 12–16, ISSN 0258-5936.
41. Contreras, O. y Moreno, F. Cobertura muerta y arvenses en la asociación Lactuca sativa - Allium ampeloprasum”. Manejo Integrado de Plagas y Agroecología, vol. 74, 2005, pp. 65-68, ISSN 1659-0082.
42. Menalled, F. D. Consideraciones ecológicas para el desarrollo de programas de manejo integrado de malezas. Agroecología, vol. 5, 2010, pp. 73-78, ISSN 1989-4686.
43. Ruffo, M. L. y Parsons, A. T. Cultivos de cobertura en sistemas agrícolas. Informaciones Agronómicas del Cono Sur, vol. 21, 2004, pp. 13–16, ISSN 1666-7115.
44. Avilés, B. W. I. y Santamaría, B. F. Periodo crítico de competencia entre maleza y henequen Agave fourcroydes Lem.”. Revista Mexicana de Ciencias Agrícolas, vol. 22, no. 1, 1 de enero de 2012, pp. 21-37, ISSN 2007-0934.
45. Altieri, M. A. y Nicholls, C. I. Una perspectiva agroecológica para una agricultura ambientalmente sana y socialmente más justa en la América Latina del siglo XXI [en línea]. En: Leff E., La transición hacia el desarrollo sustentable: perspectivas de América Latina y el Caribe, (ser. Pensamiento ambiental latinoamericano, no. ser. 6), Ed. Secretaría de Medio Ambiente y Recursos Naturales - Instituto Nacional de Ecología - Universidad Autónoma Metropolitana - PNUMA, México, D.F., 2002, pp. 281304, ISBN 978-968-817-544-6, OCLC: 52449393, [Consultado: 4 de julio de 2016], Disponible en: <http://www2.inecc.gob.mx/publicaciones/libros/363/cap12.html>.
46. Holway, D. A.; Lach, L.; Suarez, A. V.; Tsutsui, N. D. y Case, T. J. “The Causes and Consequences of Ant Invasions”. Annual Review of Ecology and Systematics, vol. 33, 2002, pp. 181-233, ISSN 0066-4162, DOI 10.1146/annurev.ecolsys.33.010802.150444.
47. Sherwood, S. y Bentley, J. “Rural farmers explore causes of plant disease”. ILEIA Newsletter, vol. 11, no. 1, 1995, pp. 20–22, ISSN 0920-8771.
48. Altieri, M. A.; Ponti, L. y Nicholls, C. I. El manejo de las plagas a través de la diversificación de las plantas. LEISA Revista de Agroecología, vol. 22, no. 4, 2007, pp. 9–13, ISSN 1729-7419.
49. Pérez, M. y Marasas, M. E. Servicios de regulación y prácticas de manejo: aportes para una horticultura de base agroecológica. Revista Ecosistemas, vol. 22, no. 1, 2013, pp. 36-43, ISSN 1697-2473, DOI 10.7818/re.2014.22-1.00.
50. García, T. L. y Fernández, Q. C. Fundamentos sobre malas hierbas y herbicidas. Ed. Ediciones Mundi-Prensa, 1991, Madrid, 348 p., ISBN 978-84-7114-331-0, OCLC: 434868704.
51. Najul, C. y Anzalone, A. “Control de malezas con cobertura vegetal en el cultivo de la Caraota negra (Phaseolus vulgaris L.)”. Bioagro, vol. 18, no. 2, 2006, pp. 75–82, ISSN 1316-3361.
52. Romero, A.; Chamorro, L. y Sans, F. X. Weed vegetation of organic and conventional dryland cereal fields in the Mediterranean region [en línea]. En: eds. Köpke U., Niggli U., Neuhoff D., Cornish P., Lockeretz W., y Willer H., Researching sustainable systems : proceedings of the First Scientific Conference of the International Society of Organic Agriculture Research (ISOFAR), Ed. ISOFAR-IFOAM-NASAA, Adelaide, South Australia, 23 de septiembre de 2005, pp. 127-130, ISBN 978-3-906081-76-2, OCLC: 255184119, [Consultado: 4 de julio de 2016], Disponible en: <http://www.isofar.org/isofar/images/conferences/Proceedings/ISOFAR_Proceedings_2005_Adelaide_AUS.pdf#page=127>.
53. Rajcan, I.; Chandler, K. J. y Swanton, C. J. “Red–far-red ratio of reflected light: a hypothesis of why early-season weed control is important in corn”. Weed Science, vol. 52, no. 5, septiembre de 2004, pp. 774-778, ISSN 0043-1745, 1550-2759, DOI 10.1614/WS-03-158R.
54. Leyva, A. y Pohlan, J. Reflexiones sobre la Agroecología en Cuba. Análisis de la Biodiversidad. Ed. Ediciones INCA, 2007, La Habana, Cuba, 290 p., ISBN 978-9597023-8.
55. Reynolds, M. P.; Pask, A. J. D.; Mullan, D. M. y Chavez-Dulanto, P. N. Fitomejoramiento fisiológico I: enfoques interdisciplinarios para mejorar la adaptación del cultivo [en línea]. Ed. Centro Internacional de Mejoramiento de Maíz y Trigo Internacional (CIMMYT), 2013, México, D. F, 188 p., ISBN 978-607-8263-18-9, [Consultado: 4 de julio de 2016], Disponible en: <https://books.google.com.cu/books?id=FgWhMhLxhzoC>.
56. Monson, R. K.; Teeri, J. A.; Ku, M. S. B.; Gurevitch, J.; Mets, L. J. y Dudley, S. “Carbon-isotope discrimination by leaves of Flaveria species exhibiting different amounts of C3-and C4-cycle co-function”. Planta, vol. 174, no. 2, mayo de 1988, pp. 145-151, ISSN 0032-0935, 1432-2048, DOI 10.1007/BF00394765.
57. Andrade, J. L.; de la Barrera, E.; Reyes, G. C.; Ricalde, M. F.; Vargas, S. G. y Cervera, J. C. El metabolismo ácido de las crasuláceas: diversidad, fisiología ambiental y productividad. Boletín de la Sociedad Botánica de México, vol. 81, no. 1, 2007, pp. 37–50, ISSN 0366-2128.
58. Villagra, P. E.; Giordano, C.; Álvarez, J. A.; Bruno, C. J.; Guevara, A.; Sartor, C.; Passera, C. B. y Greco, S. Ser planta en el desierto: estrategias de uso de agua y resistencia al estrés hídrico en el Monte Central de Argentina. Ecología Austral, vol. 21, no. 1, 2011, pp. 29–42, ISSN 0327-5477, 1667-782X.
59. Hatfield, R. D.; Marita, J. M.; Frost, K.; Grabber, J.; Ralph, J.; Lu, F. y Kim, H. “Grass lignin acylation: p-coumaroyl transferase activity and cell wall characteristics of C3 and C4 grasses”. Planta, vol. 229, no. 6, mayo de 2009, pp. 1253-1267, ISSN 0032-0935, 1432-2048, DOI 10.1007/s00425-009-0900-z.
60. Engelmann, S.; Blasing, O. E.; Gowik, U.; Svensson, P. y Westhoff, P. “Molecular evolution of C4 phosphoenolpyruvate carboxylase in the genus Flaveria-a gradual increase from C3 to C4 characteristics”. Planta, vol. 217, no. 5, 1 de septiembre de 2003, pp. 717-725, ISSN 0032-0935, 1432-2048, DOI 10.1007/s00425-003-1045-0.
61. Novellino, P.; Gil, A.; Neme, G. y Durán, V. El consumo de maíz en el Holoceno tardío del oeste argentino: isótopos estables y caries. Revista Española de Antropología Americana, vol. 34, 1 de enero de 2004, pp. 85-110, ISSN 1988-2718.
62. Afifi, M. y Swanton, C. “Maize seed and stem roots differ in response to neighbouring weeds: Maize roots differ in response to weeds”. Weed Research, vol. 51, no. 5, octubre de 2011, pp. 442-450, ISSN 0043-1737, DOI 10.1111/j.1365-3180.2011.00865.x.
63. Pardé, J.; Bouchon, J.; Prieto, R. A. y López, Q. M. Dasometria. 2.a ed., Ed. Paraninfo, 1994, Madrid, ISBN 978-84-283-2068-9, OCLC: 34206080.
64. Vadell, J.; Pascual, P. y Adrover, M. Evaluación de especies cultivadas y arvenses como abonos verdes [en línea]. En: VIII Congreso SEAE Agricultura y Alimentacion Ecologica, Ed. Sociedad Española de Agricultura Ecologica, Bullas, Murcia, España, 2008, OCLC: 912860265, [Consultado: 4 de julio de 2016], Disponible en: <http://www.agroecologia.net/recursos/publicaciones/publicaciones-online/2009/eventos-seae/cds/congresos/actas-bullas/seae_bullas/verd/sesiones/8%20S2B.%20FERTIL%28II%29/sesion8_4.pdf>.
65. Alemán, F. Manejo de malezas. 1.a ed., Ed. NUFFIC, 1991, Managua, Nicaragua, 164 p., OCLC: 49190804.
66. Sans, F. X. “La diversidad de los agroecosistemas”. Revista Ecosistemas, vol. 16, no. 1, 2007, ISSN 1697-2473, DOI 10.7818/re.2014.16-1.00, [Consultado: 4 de julio de 2016], Disponible en: <http://revistaecosistemas.net/index.php/ecosistemas/article/view/137>.
67. Rodríguez, C. C. N.; Díaz, C. J.; Villasana, R.; Pérez, P. D. y Romero, A. M. Flora segetal cañera de la Provincia de Matanzas, Cuba. Agrociencia. Serie Recursos Naturales Renovables, vol. 1, 1991, pp. 123-136, ISSN 0188-3062.
68. Yong, A. La biodiversidad florística en los sistemas agrícolas. Cultivos Tropicales, vol. 31, no. 4, diciembre de 2010, pp. 5-11, ISSN 0258-5936.
69. Boström, U. y Fogelfors, H. Long-term effects of herbicide-application strategies on weeds and yield in spring-sown cereals. Weed Science, vol. 50, no. 2, marzo de 2002, pp. 196-203, ISSN 0043-1745, 1550-2759, DOI 10.1614/0043-1745 (2002) 050[0196: LTEOHA]2.0.CO;2.
70. Boström, U. y Fogelfors, H. Response of weeds and crop yield to herbicide dose decision-support guidelines. Weed Science, vol. 50, no. 2, marzo de 2002, pp. 186-195, ISSN 0043-1745, 1550-2759, DOI 10.1614/0043- 1745(2002)050 [0186:ROWACY]2.0.CO;2.
71. Tomita, S.; Nawata, E.; Kono, Y.; Nagata, Y.; Noichana, C.; Sributta, A. y Inamura, T. “Differences in weed vegetation in response to cultivating methods and water conditions in rainfed paddy fields in north-east Thailand”. Weed Biology and Management, vol. 3, no. 2, 1 de junio de 2003, pp. 117-127, ISSN 1445-6664, DOI 10.1046/j.1445-6664.2003.00093.x.
72. Canizales, S. A.; Celemín, J. S. y Mora-Delgado, J. Diversidad y uso de arvenses en pasturas de fincas ganaderas del Alto Magdalena (Tolima, Colombia). Zootecnia Tropical, vol. 28, no. 3, septiembre de 2010, pp. 427-437, ISSN 0798-7269.
73. De la Cruz, R.; Rojas, E. y Merayo, A. “Manejo de la caminadora (Rottboellia cochinchinensis)(Lour) WD Clayton) en el cultivo de maíz y el período de barbecho con leguminosas de cobertura. Manejo Integrado de Plagas, vol. 31, 1994, pp. 29–35, ISSN 1016-0469.
74. Gliessman, S. R. Agroecología: procesos ecológicos en agricultura sostenible. Ed. CATIE, 2002, Turrialba, Costa Rica, 359 p., ISBN 978-9977-57-385-4, OCLC: 638970680.
75. Vázquez, M. L. L. “Los insectos, los agricultores y el manejo de la finca”. LEISA. Revista de Agroecología, vol. 28, no. 1, 2012, pp. 5-8, ISSN 1729-7419.
76. Vázquez, L. L. Desarrollo agroecológico de la adopción de tecnologías y la extensión para la sanidad vegetal en los sistemas agrarios de Cuba. Revista Brasileira de Agroecologia, vol. 3, no. 1, 8 de junio de 2008, pp. 3-12, ISSN 1980-9735.
77. Perdomo, F.; Vibrans, H.; Romero, A.; Domínguez, A. y Medina, L. Análisis de SHE, una herramienta para estudiar la diversidad de malezas. Revista Fitotecnia Mexicana, vol. 27, no. sp. 1, 2004, pp. 57-61, ISSN 0187-7380.
78. Altieri, M. A. y Nicholls, C. I. Conversión agroecológica de sistemas convencionales de producción: teoría, estrategias y evaluación. Revista Ecosistemas, vol. 16, no. 1, 2007, ISSN 1697-2473, DOI 10.7818/133, [Consultado: 4 de julio de 2016], Disponible en: <http://revistaecosistemas.net/index.php/ecosistemas/article/view/133> .
79. Cabellero, A. y Montes, J. Agricultura sostenible: un acercamiento a la permacultura. Ed. Praxis, 1990, Álvaro Obregón, DF, OCLC: 254449048.
80. Sagar, G. R. “On the ecology of weed control, biology pest and the disease control”. En: eds. Price J. D. y Solomon M. E., Biology in pest and disease control: the 13th Symposium of the British Ecological Society, Ed. Blackwell Scientific, Oxford, 1974, p. 450, ISBN 978-0-632-09070-9, OCLC: 1119034.
81. Uphoff, N.; Ball, A. S.; Fernandes, E.; Herren, H.; Husson, O.; Laing, M.; Palm, C.; Pretty, J.; Sanchez, P.; Sanginga, N. y Thies, J. Biological Approaches to Sustainable Soil Systems [en línea]. 1.a ed., Ed. CRC Press, 3 de marzo de 2006, Boca Raton, 784 p., ISBN 978-1-57444-583-1, [Consultado: 4 de julio de 2016], Disponible en: <https://www. amazon.com/Biological-Approaches -Sustainable- Systems -Environment/dp /1574445839?ie= UTF8&SubscriptionId= 0JRA4J6WAV0RTAZVS6R2 &camp =2025&creative= 165953&creativeASIN= 1574445839&linkCode= xm2&tag= worldcat-20>.
82. Wardle, D. A.; Bardgett, R. D.; Klironomos, J. N.; Setälä, H.; van der Putten, W. H. y Wall, D. H. Ecological Linkages Between Aboveground and Belowground Biota. Science, vol. 304, no. 5677, 11 de junio de 2004, pp. 1629-1633, ISSN 0036-8075, 1095-9203, DOI 10.1126/science.1094875.
83. Nicholls, C. I. y Altieri, M. A. Suelos saludables, plantas saludables: la evidencia agroecológica. LEISA. Revista de Agroecología, vol. 24, no. 2, 2008, pp. 6-8.
84. Jordan, N. “Prospects for Weed Control Through Crop Interference”. Ecological Applications, vol. 3, no. 1, febrero de 1993, pp. 84-91, ISSN 1051-0761, DOI 10.2307/1941794.
85. Pawar, R. K. Weed management. Ed. Oxford Book Company, 2009, Jaipur, 300 p., ISBN 978-81-89473-61-7, OCLC: 837065319.
86. Guazzelli, M. J.; Meirelles, L.; Barreto, R. y Goncalves, A. “Servicios del agroecosistema: una experiencia en la Sierra Gaucha”. LEISA. Revista de Agroecología, vol. 22, no. 4, 2007, pp. 5-8, ISSN 1729-7419.
87. Primavesi, A. Agricultura sustentável: manual do produtor rural, maior produtividade, maiores lucros, respeito à terra. Ed. Nobel, 1992, São Paulo, Brazil, 142 p., ISBN 978-85-213-0730-3, OCLC: 36213448.
88. Primavesi, A. Manejo ecologico de pragas e doencas: tecnicas alternativas para a producao agropecuaria e defesa do meio ambiente. Ed. Nobel, 1988, Sao Paulo, Brazil, 137 p., ISBN 978-85-213-0546-0, OCLC: 708562606.
89. Bunch, R. “El manejo del suelo vivo”. LEISA. Revista de Agroecología, vol. 24, no. 2, 2008, p. 5, ISSN 1729-7419.
90. Altieri, M. A. El estado del arte en la agroecología: revisando avances y desafíos. En: León S. T. E. y Altieri M. A., Vertientes del pensamiento agroecológico: fundamentos y aplicaciones, Ed. Universidad Nacional de Colombia, Bogotá, 2010, pp. 77104, ISBN 978-958-719-572-9, OCLC: 777063181.
Recibido: 05/08/2015
Aceptado: 29/03/2016
M.Cs. Yaisys Blanco Valdes, Instituto Nacional Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32700. Email: yblanco@inca.edu.cu


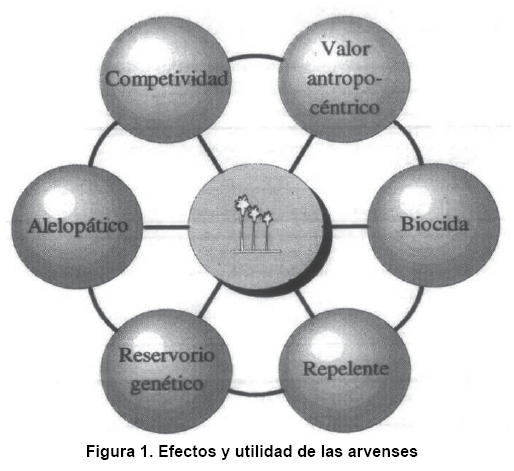{kind=link}
{kind=link}
{kind=link}
{kind=link}