










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCultivos Tropicales
versión On-line ISSN 1819-4087
cultrop vol.37 supl.1 La Habana 2016
REVISIÓN BIBLIOGRÁFICA
Biosíntesis de ácido indol-3-acético y promoción del crecimiento de plantas por bacterias
Biosynthesis of indole-3-acetic acid and plant growth promoting by bacteria
Paulina Vega-Celedón,I Hayron Canchignia Martínez, I, II Myriam González,I Michael SeegerI
IInstituto de Investigaciones Fundamentales en Agricultura Tropical Alejandro de Humboldt, INIFAT. Cuba.
IIUniversidad de La Habana. Cuba.
RESUMEN
La hormona vegetal ácido indol-3-acético (AIA) es la principal auxina en las plantas. El AIA controla diversos procesos fisiológicos como la elongación y división celular, la diferenciación de tejidos y las respuestas a la luz y la gravedad. La concentración de AIA se encuentra regulada en las plantas. Se ha descrito que las bacterias pueden modular los niveles de AIA. Las rutas biosintéticas de AIA más importantes y ampliamente distribuidas en bacterias son las vías anabólicas de indol-3-piruvato (IPA) y de indol-3-acetamida (IAM). Estas rutas metabólicas son dependientes del precursor triptófano. La vía IPA ha sido descrita principalmente en bacterias promotoras del crecimiento (PGPB), mientras que la vía IAM está presente en bacterias fitopatógenas. Las PGPB estimulan el crecimiento de las plantas mediante diversos mecanismos, que incluyen la producción de fitohormonas. Diversos estudios sobre PGPB que poseen la capacidad de producir AIA demuestran que esta fitohormona juega un rol determinante en la promoción del crecimiento vegetal.
Palabras clave: ácido indolacético, auxina, bacteria, fitohormona, planta, triptófano.
ABSTRACT
The plant hormone indole-3-acetic acid (IAA) is the main auxin in plants. IAA controls several physiological processes such as cell elongation and division, tissue differentiation and responses to light and gravity. IAA concentration is regulated in plants. Interestingly, bacteria can modulate IAA levels. The most widely distributed biosynthetic IAA routes in bacteria are indole-3-pyruvate (IPA) and indole-3-acetamide (IAM) anabolic pathways. These metabolic pathways use tryptophan as precursor. IPA pathway has been described mainly in plant growth-promoting bacteria (PGPB), whereas the IAM pathway is present in phytopathogenic bacteria. PGPB stimulate plant growth through various mechanisms including the production of plant hormones. Studies on PGPB that are capable to produce IAA indicate that this phytohormone plays a crucial role for promoting plant growth.
Key words: indole acetic acid, auxin, bacteria, phytohormone, plant, tryptophan.
INTRODUCCIÓN
Hormonas vegetales
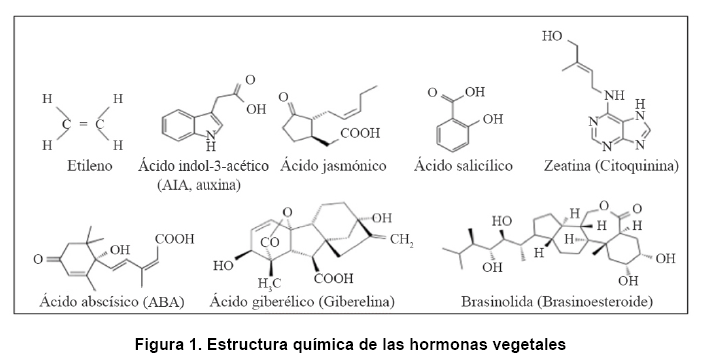
- Estudios pioneros en el siglo XIX realizados por Julius von Sachs y Charles Darwin demostraron que varios procesos del crecimiento de las plantas estaban regulados por sustancias que se transportaban de una parte de la planta a otra (1). Más de un siglo después, se conoce que estas sustancias son pequeñas moléculas estructuralmente no relacionadas que derivan de vías metabólicas esenciales de las plantas. En general, estos compuestos están presentes en concentraciones muy bajas y actúan localmente, alrededor del sitio de síntesis o en tejidos distantes. En las últimas décadas se han ido identificando nuevas hormonas vegetales, que incluyen auxinas como el ácido indol-3-acético (AIA), ácido abscísico (ABA), brasinoesteroides (BRS), citoquininas, giberelinas, etileno, ácido jasmónico (AJ) y ácido salicílico (AS) (Figura 1). Las auxinas, citoquininas, ABA, etileno, ácido salicílico y giberelinas pueden ser producidas también por bacterias (2, 3, 4, 5, 6, 7). Estas hormonas regulan todos los aspectos de la vida vegetal, desde la formación del patrón durante el desarrollo hasta las respuestas al estrés biótico y abiótico (8, 9).
Una lista parcial de las respuestas de cada fitohormona se describe a continuación (8). El gas etileno promueve la maduración de la fruta, senescencia, respuestas a patógenos y al estrés abiótico. El AIA (auxina) regula la división y expansión celular, diferenciación vascular, desarrollo de raíz lateral y dominancia apical. El ácido jasmónico es una señal volátil que modula el desarrollo del polen y las respuestas a infecciones por patógenos. El ácido salicílico participa en el crecimiento y desarrollo de la planta, en los mecanismos de resistencia a fitopatógenos, y en respuestas al estrés abiótico y biótico (10). Las citoquininas son derivados de adenina identificados por su capacidad de promover la citoquinesis. El ácido abscísico promueve la latencia de la semilla y participa en varias vías de señalización de estrés. Las giberelinas son compuestos diterpenoides que promueven la germinación, alargamiento del tallo y la inducción de la floración. Los brasinoesteroides regulan la expansión celular y la fotomorfogénesis.
Auxinas
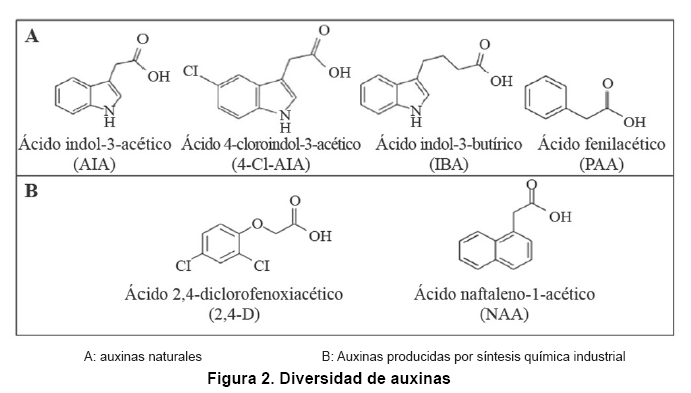
Las auxinas son hormonas de crecimiento de plantas, debido a su capacidad de estimular el crecimiento diferencial en respuesta a estímulos de luz (11). Son las fitohormonas que juegan el rol más importante en el desarrollo de las plantas (12). Se han descrito cuatro auxinas naturales sintetizadas por plantas: AIA, ácido indol-3-butírico (IBA), ácido 4-cloroindol-3-acético (4-Cl-AIA) y ácido fenilacético (PAA) (13) (Figura 2A). Ensayos in vitro han permitido determinar que el AIA es una de las principales auxinas que participan en el desarrollo de las plantas (14, 15).
La industria agroquímica ha desarrollado auxinas sintéticas que imitan la estructura de AIA. Las auxinas sintéticas más ampliamente utilizadas como herbicidas son el ácido 2,4-diclorofenoxiacético (2,4-D) y el ácido naftaleno-1-acético (NAA) (13) (Figura 2B). En contraste con AIA, estos herbicidas poseen una larga vida meda, debido a su alta estabilidad en plantas superiores y, por lo tanto, mayor efectividad que AIA. En bajas concentraciones son capaces de estimular los procesos de desarrollo y crecimiento de plantas, pero a altas concentraciones el crecimiento se ve perturbado y se daña la planta causando su muerte (16, 17).
Ácido indol-3-acético (AIA)
El AIA es la principal auxina nativa de las plantas superiores (14). El AIA está involucrado en el crecimiento y desarrollo de las plantas, principalmente en una serie de procesos fisiológicos que incluyen el alargamiento y división celular, diferenciación de tejido, fototropismo, gravitropismo y en respuestas defensivas (9, 18), destacando un importante rol en la formación del xilema y la raíz (19).
La biosíntesis de AIA no está limitada a las plantas superiores. Organismos como bacterias, hongos y algas son capaces de sintetizar AIA, lo que puede afectar el crecimiento y el desarrollo de las plantas (20). En bacterias, la producción de AIA es una capacidad relevante, tanto de PGPBs, como de bacterias fitopatógenas (21, 22). Las plantas superiores exudan, entre otros componentes el aminoácido triptófano, que es el principal precursor para la biosíntesis de AIA microbiano (23, 24, 25, 26).
Biosíntesis de AIA en bacterias
Se ha propuesto que el 80 % de las bacterias de la rizósfera son capaces de producir AIA. Se ha observado un alto grado de similitud en las vías de síntesis de AIA de plantas y bacterias (27).
Los microorganismos de rizósfera que interaccionan con las plantas, interfieren con su desarrollo, mediante el desbalance de auxinas, de manera que pueden afectar a la planta en forma positiva o negativa (7). Las bacterias productoras de AIA (BIP, del inglés bacterial IAA producer) tienen el potencial de interferir con los procesos de incorporación de AIA en las plantas. La consecuencia para la planta depende de la cantidad de AIA producida y la sensibilidad del tejido de la planta a los cambios en la concentración de AIA (18), lo que se asocia a la vía biosintética utilizada por las bacterias asociadas a plantas (27). Bacterias fitopatogénicas como Agrobacterium spp. y Pseudomonas savastanoi pv. savastanoi causan en el tejido vegetal tumores y corona de agallas, respectivamente, que en plantas jóvenes significan retrasos en el crecimiento, el desarrollo anormal, la susceptibilidad a otras enfermedades o la muerte de la planta (28, 29). Por otra parte, la PGPB Azospirillum favorece el desarrollo de la raíz (27, 30). El triptófano es el principal precursor en las vías de biosíntesis de AIA en bacterias (27, 31). Diferentes vías de síntesis de AIA han sido identificadas en bacterias. Cinco de estas rutas anabólicas son dependientes de triptófano. Las vías de síntesis de AIA más importantes y ampliamente distribuidas son la vía indol-3-piruvato (IPA) y la vía indol-3-acetamida (IAM) (7). La vía IPA ha sido reportada principalmente en PGPB, mientras que la vía IAM ha sido descrita en bacterias fitopatógenas (7, 27) (Figura 3). Además se han descrito vías de síntesis de AIA independientes de triptófano, que han sido estudiadas mediante mutantes y marcaje isotópico.
Estas rutas anabólicas en presencia de triptófano aumentan significativamente la producción de AIA (20). El aminoácido triptófano es uno de los principales componentes del exudado de las raíces (26). La biosíntesis bacteriana de AIA puede contribuir a la supervivencia de bacterias, mediante la destoxificación de triptófano del exudado de las plantas (27).
Vía indol-3-piruvato (IPA)
La vía IPA ha sido propuesta como la principal ruta para la biosíntesis de AIA en plantas. Sin embargo, las enzimas o genes clave no han sido aún identificados en plantas (7). La producción de AIA por la vía IPA ha sido descrita en diversas bacterias, como el fitopatógeno Pantoea agglomerans, bacterias benéficas (e.g., Bradyrhizobium, Azospirillum, Rhizobium y Enterobacter cloacae) y en cianobacterias. Esta vía tiene como primera etapa la conversión de triptófano a ácido indol-3-pirúvico (IPA) por una aminotransferasa. En una segunda etapa el IPA se descarboxila a ácido indol-3-acetaldehído (AIAld) por la indol-3-piruvato descarboxilasa (IPDC), siendo esta la etapa limitante de la síntesis. El gen ipdC que codifica para la enzima IPDC ha sido caracterizado en algunas bacterias como Azospirillum brasilense, E. cloacae, Pseudomonas putida, P. agglomerans y Paenibacillus polymyxa (22, 27, 32). En la etapa final, el AIAld es oxidado a AIA por la indol-3-acetaldehído deshidrogenasa (7, 27).
Vía indol-3-acetamida (IAM)
La vía IAM es la ruta más caracterizada en bacterias. Esta vía no ha sido descrita en plantas (27).
Esta vía metabólica está presente en diversos patógenos y contribuye a la virulencia de estas bacterias por la excesiva producción de AIA (28, 30). Los síntomas fitopatogénicos están principalmente ligados a la vía IAM, dado que sería microbiana específica (27).
Debido a que las plantas no poseen los intermediarios metabólicos de esta vía, no son capaces de mantener el AIA a niveles no tóxicos o fisiológicamente apropiados en sus tejidos por retroalimentación (30). Los genes, las proteínas y la regulación de esta ruta metabólica han sido caracterizados y comprende dos etapas. En la primera, el triptófano es convertido en IAM por la enzima triptófano-2-monooxigenasa (IaaM), que está codificada en el gen iaaM. En la segunda, el IAM es convertido a AIA y amonio, mediante una IAM hidrolasa (IaaH), que es codificada por el gen iaaH.
Estos genes han sido descritos en bacterias como Agrobacterium tumefaciens, Pseudomonas savastanoi, Pseudomonas syringae, P. agglomerans, Rhizobium sp. NGR234 y Bradyrhizobium japonicum (7, 27, 33, 34).
Bacterias promotoras del crecimiento de plantas
Las bacterias promotoras del crecimiento de plantas (PGPB, plant-growth promoting bacteria) son un grupo de bacterias benéficas de plantas (35).
Las PGPB participan en el crecimiento y la salud de las plantas, suprimiendo daños efectuados por microorganismos patógenos, sintetizando fitohormonas, acelerando la disponibilidad y la asimilación de nutrientes del suelo mediante diferentes mecanismos como la fijación del nitrógeno atmosférico, la solubilización del fósforo y la síntesis de sideróforos (36, 37). En la agricultura, las PGPB han sido utilizadas como una alternativa sustentable con el medio ambiente, dado que tienen la capacidad de mejorar la calidad de los suelos y el rendimiento de los cultivos, reduciendo el impacto negativo de los fertilizantes químicos (38). Las PGPB han sido extensivamente estudiadas durante los últimos años y pertenecen a los géneros Acetobacter, Acinetobacter, Alcaligenes, Arthrobacter, Azoarcus, Azospirillum, Azotobacter, Bacillus, Beijerinckia, Burkholderia, Derxia, Enterobacter, Gluconacetobacter, Herbaspirillum, Klebsiella, Ochrobactrum, Pantoae, Pseudomonas, Rhodococcus, Serratia, Stenotrophomonas y Zoogloea (36).
Bacterias promotoras del crecimiento de plantas productoras de ácido indol-3-acético
El AIA producido por PGPB presenta un gran impacto sobre su actividad promotora del crecimiento de plantas. Diversas bacterias de las clases taxonómicas α-Proteobacteria, β-Proteobacteria, δ-Proteobacteria y Bacilli son PGPB productoras de AIA; algunos ejemplos de PGPB se muestran en la tabla. Se estudió el efecto fitoestimulante de 16 cepas de Bacillus sobre el crecimiento de la planta Vigna radiata. B. megaterium MiR-4, que correspondió a la mayor productora de AIA, presentó el mayor efecto promotor del crecimiento, aumentando la elongación de los brotes y el número de raíces (39).
Bacterias mutantes del gen ipdC pierden capacidad promotora de crecimiento de plantas. La longitud de raíces de esquejes de canola de semillas tratadas con
P. putida GR12-2 fueron un 35 % más largas que las raíces de semillas tratadas con la mutante ipdC- (22).
La bacteria ipdC- de Azospirillum brasilense SM (SMIT56s10) disminuyó el efecto promotor del crecimiento sobre el número y longitud de raíces de semillas de sorgo (40).
Burkholderia xenovorans LB400, bacteria modelo para la degradación de diversos compuestos aromáticos, incluyendo los policlorobifenilos, tiene la capacidad de producir AIA mediante la vía IPA. La cepa LB400 promovió el crecimiento sobre plantas como Nicotiana tabacum y Vitis vinifera, incrementando de esta manera su versatilidad metabólicaA (41).
CONCLUSIONES
- La auxina ácido indol-3-acético es una fitohormona de gran importancia para el desarrollo vegetal, que puede ser sintetizada mediante diversas rutas metabólicas.
- El AIA producido por PGPB juega un rol significativo en la interacción planta-microorganismo, contribuyendo en la promoción del crecimiento radicular y foliar de la planta.
AGRADECIMIENTOS
Los autores agradecen el soporte financiero otorgado por las becas doctorales CONICYT y RIABIN (PVC, HCM), FONDECYT (1110992 & 1151174) (http://www.fondecyt.cl) (MS), Universidad Técnica Federico Santa María (131109, 131342 & 131562) (http://www.usm.cl) (MS, MG) y Center for Nanotechnology and Systems Biology (http://www.usm.cl) (MS).
Nota al pie
AVega-Celedón, P.; Canchignia, H.; González, M. y Seeger, M. Metabolism of indole-3-acetic acid in Burkholderia xenovorans LB400 and its effect on Vitis vinifera growth. En: XXI Congreso Latinoamericano de Microbiología, Santos, Brasil, 2012.
BIBLIOGRAFÍA
1. Darwin, C. The Power of Movement in Plants. edit. HardPress Publishing, 29 de enero de 2010, 352 p., ISBN 978-1-4076-3307-7.
2. Press, C. M.; Wilson, M.; Tuzun, S. y Kloepper, J. W. Salicylic Acid Produced by Serratia marcescens 90-166 Is Not the Primary Determinant of Induced Systemic Resistance in Cucumber or Tobacco. Molecular Plant-Microbe Interactions, vol. 10, no. 6, 1 de agosto de 1997, pp. 761-768, ISSN 0894-0282, DOI 10.1094/MPMI.1997.10.6.761.
3. Weingart, H. y Volksch, B. Ethylene Production by Pseudomonas syringae Pathovars In Vitro and In Planta.. Applied and Environmental Microbiology, vol. 63, no. 1, 1 de enero de 1997, pp. 156-161, ISSN 0099-2240, 1098-5336, PMID: 16535480.
4. Bottini, R.; Cassán, F. y Piccoli, P. Gibberellin production by bacteria and its involvement in plant growth promotion and yield increase. Applied Microbiology and Biotechnology, vol. 65, no. 5, 28 de julio de 2004, pp. 497-503, ISSN 0175-7598, 1432-0614, DOI 10.1007/s00253-004-1696-1.
5. Boiero, L.; Perrig, D.; Masciarelli, O.; Penna, C.; Cassán, F. y Luna, V. Phytohormone production by three strains of Bradyrhizobium japonicum and possible physiological and technological implications. Applied Microbiology and Biotechnology, vol. 74, no. 4, 29 de noviembre de 2006, pp. 874-880, ISSN 0175-7598, 1432-0614, DOI 10.1007/s00253-006-0731-9.
6. Frébort, I.; Kowalska, M.; Hluska, T.; Frébortová, J. y Galuszka, P. Evolution of cytokinin biosynthesis and degradation. Journal of Experimental Botany, vol. 62, no. 8, 2011, pp. 2431–2452, ISSN 1460-2431.
7. Spaepen, S. y Vanderleyden, J. Auxin and Plant-Microbe Interactions. Cold Spring Harbor Perspectives in Biology, vol. 3, no. 4, 4 de enero de 2011, p. a001438, ISSN 1943-0264, DOI 10.1101/cshperspect.a001438, PMID: 21084388.
8. Gray, W. M. Hormonal Regulation of Plant Growth and Development. PLoS Biology, vol. 2, no. 9, 14 de septiembre de 2004, p. 311, ISSN 1544-9173, 1545-7885, DOI 10.1371/journal.pbio.0020311.
9. Santner, A.; Calderon-Villalobos, L. I. A. y Estelle, M. Plant hormones are versatile chemical regulators of plant growth. Nature chemical biology, vol. 5, no. 5, 2009, pp. 301–307, ISSN 1552-4450, 1552-4469.
10. Rivas, S. V. M. y Plasencia, J. Salicylic acid beyond defence: its role in plant growth and development. Journal of Experimental Botany, vol. 62, no. 10, 2011, pp. 3321-3338, ISSN 0022-0957, 1460-2431.
11. Zhao, Y. Auxin biosynthesis and its role in plant development. Annual Review of Plant Biology, vol. 61, 2010, p. 49, ISSN 15452123, 15435008.
12. Wisniewska, J.; Xu, J.; Seifertová, D.; Brewer, P. B.; Ruicka, K.; Blilou, I.; Rouquié, D.; Benková, E.; Scheres, B. y Friml, J. Polar PIN localization directs auxin flow in plants. Science, vol. 312, no. 5775, 2006, pp. 883–883, ISSN 0036-8075, 1095-9203.
13. Simon, S. y Petráek, J. Why plants need more than one type of auxin. Plant Science, vol. 180, no. 3, marzo de 2011, pp. 454-460, ISSN 0168-9452, DOI 10.1016/j.plantsci.2010.12.007.
14. Bonner, J. y Bandurski, R. S. Studies of the Physiology, Pharmacology, and Biochemistry of the Auxins. Annual Review of Plant Physiology, vol. 3, no. 1, 1 de junio de 1952, pp. 59-86, ISSN 0066-4294, DOI 10.1146/annurev.pp.03.060152.000423.
15. Mashiguchi, K.; Tanaka, K.; Sakai, T.; Sugawara, S.; Kawaide, H.; Natsume, M.; Hanada, A.; Yaeno, T.; Shirasu, K. y Yao, H. The main auxin biosynthesis pathway in Arabidopsis. Proceedings of the National Academy of Sciences, vol. 108, no. 45, 2011, pp. 18512-18517, ISSN 1091-6490.
16. Grossmann, K. Mediation of Herbicide Effects by Hormone Interactions. Journal of Plant Growth Regulation, vol. 22, no. 1, 21 de agosto de 2003, pp. 109-122, ISSN 0721-7595, 1435-8107, DOI 10.1007/s00344-003-0020-0.
17. Grossmann, K. Auxin herbicide action: lifting the veil step by step. Plant Signaling & Behavior, vol. 2, no. 5, 2007, pp. 421–423, ISSN 1559-2316, 1559-2324, DOI 10.4161/psb.2.5.4417.
18. Leveau, J. H. J. y Lindow, S. E. Utilization of the Plant Hormone Indole-3-Acetic Acid for Growth by Pseudomonas putida Strain 1290. Applied and Environmental Microbiology, vol. 71, no. 5, 5 de enero de 2005, pp. 2365-2371, ISSN 0099-2240, 1098-5336, DOI 10.1128/AEM.71.5.2365-2371.2005, PMID: 15870323.
19. Davies, P. J. Plant hormones: physiology, biochemistry and molecular biology [en línea]. edit. Springer Netherlands, Dordrecht, 1995, ISBN 978-0-7923-2985-5, [Consultado: 25 de enero de 2016], Disponible en: <http://link.springer.com/10.1007/978-94-011-0473-9>.
20. Lee, S.; Flores-Encarnación, M.; Contreras-Zentella, M.; Garcia-Flores, L.; Escamilla, J. E. y Kennedy, C. Indole-3-Acetic Acid Biosynthesis Is Deficient in Gluconacetobacter diazotrophicus Strains with Mutations in Cytochrome c Biogenesis Genes. Journal of Bacteriology, vol. 186, no. 16, 15 de agosto de 2004, pp. 5384-5391, ISSN 0021-9193, 1098-5530, DOI 10.1128/JB.186.16.5384-5391.2004, PMID: 15292139.
21. Patten, C. L. y Glick, B. R. Bacterial biosynthesis of indole-3-acetic acid. Canadian Journal of Microbiology, vol. 42, no. 3, 1 de marzo de 1996, pp. 207-220, ISSN 0008-4166, DOI 10.1139/m96-032.
22. Patten, C. L. y Glick, B. R. Role of Pseudomonas putida Indoleacetic Acid in Development of the Host Plant Root System. Applied and Environmental Microbiology, vol. 68, no. 8, 8 de enero de 2002, pp. 3795-3801, ISSN 0099-2240, 1098-5336, DOI 10.1128/AEM.68.8.3795-3801.2002, PMID: 12147474.
23. Kravchenko, L. V.; Makarova, N. M.; Azarova, T. S.; Provorov, N. A. y Tikhonovich, I. A. Isolation and Phenotypic Characterization of Plant Growth–Promoting Rhizobacteria with High Antiphytopathogenic Activity and Root-Colonizing Ability. Microbiology, vol. 71, no. 4, julio de 2002, pp. 444-448, ISSN 0026-2617, 1608-3237, DOI 10.1023/A:1019849711782.
24. Kravchenko, L. V.; Azarova, T. S.; Makarova, N. M. y Tikhonovich, I. A. The Effect of Tryptophan Present in Plant Root Exudates on the Phytostimulating Activity of Rhizobacteria. Microbiology, vol. 73, no. 2, marzo de 2004, pp. 156-158, ISSN 0026-2617, 1608-3237, DOI10.1023/B:MICI. 0000023982.76684.9d.
25. Kamilova, F.; Kravchenko, L. V.; Shaposhnikov, A. I.; Azarova, T.; Makarova, N. y Lugtenberg, B. Organic Acids, Sugars, and l-Tryptophane in Exudates of Vegetables Growing on Stonewool and Their Effects on Activities of Rhizosphere Bacteria. Molecular Plant-Microbe Interactions, vol. 19, no. 3, 1 de marzo de 2006, pp. 250-256, ISSN 0894-0282, DOI 10.1094/MPMI-19-0250.
26. Idris, E. E.; Iglesias, D. J.; Talon, M. y Borriss, R. Tryptophan-Dependent Production of Indole-3-Acetic Acid (IAA) Affects Level of Plant Growth Promotion by Bacillus amyloliquefaciens FZB42. Molecular Plant-Microbe Interactions, vol. 20, no. 6, 30 de mayo de 2007, pp. 619-626, ISSN 0894-0282, DOI 10.1094/MPMI-20-6-0619.
27. Spaepen, S.; Vanderleyden, J. y Remans, R. Indole-3-acetic acid in microbial and microorganism-plant signaling. FEMS Microbiology Reviews, vol. 31, no. 4, 2007, pp. 425–448, ISSN 1574-6976.
28. Jameson, P. E. Cytokinins and auxins in plant-pathogen interactions – An overview. Plant Growth Regulation, vol. 32, no. 2-3, noviembre de 2000, pp. 369-380, ISSN 0167-6903, 1573-5087, DOI 10.1023/A:1010733617543.
29. Mole, B. M.; Baltrus, D. A.; Dangl, J. L. y Grant, S. R. Global virulence regulation networks in phytopathogenic bacteria. Trends in Microbiology, vol. 15, no. 8, agosto de 2007, pp. 363-371, ISSN 0966-842X, DOI 10.1016/j.tim.2007.06.005.
30. Persello-Cartieaux, F.; Nussaume, L. y Robaglia, C. Tales from the underground: molecular. Plant, Cell & Environment, vol. 26, no. 2, 1 de febrero de 2003, pp. 189-199,
ISSN 1365-3040, DOI 10.1046/j.13653040.2003.00956.x.
31. Tsavkelova, E. A.; Cherdyntseva, T. A.; Botina, S. G. y Netrusov, A. I. Bacteria associated with orchid roots and microbial production of auxin. Microbiological Research, vol. 162, no. 1, 29 de enero de 2007, pp. 69-76, ISSN 0944-5013, DOI 10.1016/j.micres.2006.07.014.
32. Phi, Q.-T.; Park, Y.-M.; Ryu, C.-M.; Park, S.-H. y Ghim, S.-Y. Functional identification and expression of indole-3-pyruvate decarboxylase from Paenibacillus polymyxa E681. Journal of Microbiology and Biotechnology, vol. 18, 2008, pp. 1235–1244, ISSN 1738-8872, 1017-7825.
33. Glickmann, E.; Gardan, L.; Jacquet, S.; Hussain, S.; Elasri, M.; Petit, A. y Dessaux, Y. Auxin Production Is a Common Feature of Most Pathovars of Pseudomonas syringae. Molecular Plant-Microbe Interactions, vol. 11, no. 2, 1 de febrero de 1998, pp. 156-162, ISSN 0894-0282, DOI 10.1094MPMI.1998.11.2.156.
34. Yang, S.; Zhang, Q.; Guo, J.; Charkowski, A. O.; Glick, B. R.; Ibekwe, A. M.; Cooksey, D. A. y Yang, C.-H. Global Effect of Indole-3-Acetic Acid Biosynthesis on Multiple Virulence Factors of Erwinia chrysanthemi 3937. Applied and Environmental Microbiology, vol. 73, no. 4, 15 de febrero de 2007, pp. 1079-1088, ISSN 0099-2240, 1098-5336,
DOI 10.1128/AEM.01770-06, PMID: 17189441.
35. Lugtenberg, B. y Kamilova, F. Plant-Growth-Promoting Rhizobacteria. Annual Review of Microbiology, vol. 63, no. 1, 2009, pp. 541-556, ISSN 0066-4227, 1545-3251, DOI 10.1146/annurev.micro.62.081307.162918, PMID: 19575558.
36. Babalola, O. O. Beneficial bacteria of agricultural importance. Biotechnology Letters, vol. 32, no. 11, 16 de julio de 2010, pp. 1559-1570, ISSN 0141-5492, 1573-6776, DOI 10.1007/s10529-010-0347-0.
37. Adesemoye, A. O.; Torbert, H. A. y Kloepper, J. W. Plant Growth-Promoting Rhizobacteria Allow Reduced Application Rates of Chemical Fertilizers. Microbial Ecology, vol. 58, no. 4, 23 de mayo de 2009, pp. 921-929, ISSN 0095-3628, 1432-184X, DOI 10.1007/s00248-009-9531-y.
38. Castro-Sowinski, S.; Herschkovitz, Y.; Okon, Y. y Jurkevitch, E. Effects of inoculation with plant growth-promoting rhizobacteria on resident rhizosphere microorganisms. FEMS Microbiology Letters, vol. 276, no. 1, 2007, pp. 1–11, ISSN 0378-1097, 1574-6968.
39. Ali, B.; Sabri, A. N.; Ljung, K. y Hasnain, S. Quantification of indole-3-acetic acid from plant associated Bacillus spp. and their phytostimulatory effect on Vigna radiata (L.). World Journal of Microbiology and Biotechnology, vol. 25, no. 3, 16 de diciembre de 2008, pp. 519-526, ISSN 0959-3993, 1573-0972, DOI 10.1007/s11274-008-9918-9.
40. Malhotra, M. y Srivastava, S. An ipdC gene knock-out of Azospirillum brasilense strain SM and its implications on indole-3-acetic acid biosynthesis and plant growth promotion. Antonie van Leeuwenhoek, vol. 93, no. 4, 20 de octubre de 2007, pp. 425-433, ISSN 0003-6072, 1572-9699, DOI 10.1007/s10482-007-9207-x.
41. Vega-Celedón, P.; Canchignia, H.; González, M. y Seeger, M. Biosíntesis de ácido indol-3-acético y promoción del crecimiento de plantas por la bacteria Burkholderia xenovorans LB400. En: X Congreso Internacional de Biotecnología Vegetal, edit. Centro de Bioplantas. Universidad de Ciego de Ávila, Ciego de Ávila, Cuba, 2015.
42. Spaepen, S.; Bossuyt, S.; Engelen, K.; Marchal, K. y Vanderleyden, J. Phenotypical and molecular responses of Arabidopsis thaliana roots as a result of inoculation with the auxin-producing bacterium Azospirillum brasilense. New Phytologist, vol. 201, no. 3, 2014, pp. 850–861, ISSN 1469-8137.
43. Sevilla, M.; Burris, R. H.; Gunapala, N. y Kennedy, C. Comparison of Benefit to Sugarcane Plant Growth and 15N2 Incorporation Following Inoculation of Sterile Plants with Acetobacter diazotrophicus Wild-Type and Nif¯ Mutant Strains. Molecular Plant-Microbe Interactions, vol. 14, no. 3, 1 de marzo de 2001, pp. 358-366, ISSN 0894-0282,
DOI 10.1094MPMI.2001.14.3.358.
44. Poupin, M. J.; Timmermann, T.; Vega, A.; Zuñiga, A. y González, B. Effects of the Plant Growth-Promoting Bacterium Burkholderia phytofirmans PsJN throughout the Life Cycle of Arabidopsis thaliana. PLoS ONE, vol. 8, no. 7, 15 de julio de 2013, p. 69435, ISSN 1932-6203, DOI 10.1371/journal.pone.0069435.
45. Singh, R. K.; Malik, N. y Singh, S. Improved Nutrient Use Efficiency Increases Plant Growth of Rice with the Use of IAA-Overproducing Strains of Endophytic Burkholderia cepacia Strain RRE25. Microbial Ecology, vol. 66, no. 2, 25 de abril de 2013, pp. 375-384, ISSN 0095-3628, 1432-184X, DOI 10.1007/s00248-013-0231-2.
46. Apine, O. A. y Jadhav, J. P. Optimization of medium for indole-3-acetic acid production using Pantoea agglomerans strain PVM. Journal of Applied Microbiology, vol. 110, no. 5, 2011, pp. 1235–1244, ISSN 1365-2672.
47. Idris, E. E.; Bochow, H.; Ross, H. y Borriss, R. Use of Bacillus subtilis as biocontrol agent. VI. Phytohormone-like action of culture filtrates prepared from plant growth-promoting Bacillus amyloliquefaciens FZB24, FZB42, FZB45 and Bacillus subtilis FZB37. Journal of Plant Diseases and Protection, vol. 111, no. 6, 2004, pp. 583–597, ISSN 0340-8159.
48. Ryu, C.; Kim, J.; Choi, O.; Park, S.; Park, S. y Park, C. Nature of a root-associated Paenibacillus polymyxa from field-grown winter barley in Korea. Journal of Microbiology and Biotechnology, vol. 15, no. 5, 2005, p. 984, ISSN 1017-7825.
Recibido: 15 de mayo de 2015
Aceptado: 20 de enero de 2016
Paulina Vega-Celedón, Universidad Técnica Federico Santa María, Valparaíso, Chile. Email: michael.seeger@usm.cl


{kind=link}
{kind=link}
{kind=link}
{kind=link}