










Meu SciELO
 Serviços customizados
Serviços customizadosServiços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkCultivos Tropicales
versão impressa ISSN 0258-5936versão On-line ISSN 1819-4087
cultrop vol.38 no.3 La Habana july.-set. 2017
ARTÍCULO ORIGINAL
Efecto de la 24-epibrasinólida en el crecimiento y la fotosíntesis de plantas jóvenes de arroz tratadas con NaCl
Effect of 24-epibrassinolide on growth and photosynthesis of young rice plants treated with naCl
M.Cs. Yanelis Reyes,I Lisbel Martínez,I Dr.C. José Dell´Amico,I Dra.C. María C. González,I Dr.C. Michael Deyholos,II Dra.C. Miriam NúñezI
IInstituto Nacional de Ciencias Agrícolas. Gaveta Postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32700.
IIDepartamento de Ciencias Biológicas. Universidad de Alberta. Edmonton. Alberta, Canadá.
RESUMEN
La salinidad es una de las causas de estrés más acuciantes en la actualidad y los brasinoesteroides son compuestos que, además, de estimular el crecimiento vegetal aumentan la tolerancia de las plantas al estrés salino. En el presente trabajo, se evaluó el efecto de la aspersión foliar con un brasinoesteroide natural (24-epibrasinólida, EBL) en plántulas de arroz sometidas a este tipo de estrés. Para ello, se asperjaron plantas jóvenes de arroz de los cultivares J-104 y Ginés con EBL (2 µmol L-1), la mitad de ellas se trataron con NaCl (100 mmol L-1) por siete días, y posteriormente, se transfirieron a solución nutritiva para evaluar su recuperación durante 14 días. Las evaluaciones de crecimiento (longitudes de la parte aérea y de las raíces y masas secas de la parte aérea y de las raíces) se realizaron a los tres y siete días del tratamiento salino y a los siete y 14 días de recuperación. También se determinaron algunos indicadores de la fotosíntesis como tasa fotosintética, conductancia estomática, transpiración, concentración interna de CO2 y pigmentos fotosintéticos (clorofilas y carotenos). Los resultados demostraron que, de forma general, la aspersión foliar con EBL (2 µmol L-1) estimuló la longitud y la masa seca de las plántulas de arroz de ambos cultivares en condiciones salinas, lo que parece estar asociado a un incremento de la fotosíntesis y el intercambio gaseoso, así como a un aumento de la concentración de clorofilas.
Palabras clave: brasinoesteroides, clorofilas, Oryza sativa, salinidad.
ABSTRACT
At present, salinity is one of the most pressing causes of stress, and brassinosteroids are compounds which stimulate plant growth and also, increase the plant tolerance to salt stress. In this paper, the effect of foliar spraying with a natural brassinosteroid (24-epibrassinolide, EBL) in rice seedlings subjected to this kind of stress was evaluated. To do this, rice young plants of J-104 and Gines cultivars were sprayed with EBL (2 μmol L-1), and then, half of them were treated with NaCl (100 mmol L-1) for seven days, and later, they were transferred to nutrient solution to assess recovery for 14 days. Evaluations of growth (length and dry mass of aerial parts and roots) were performed at three and seven days of saline treatment and seven and 14 days of recovery. Some indicators of photosynthesis such as photosynthetic rate, stomatal conductance, transpiration, intercellular CO2 concentration and photosynthetic pigments (chlorophylls and carotenes) were also determined. The results showed that, in general, foliar spraying with EBL 2 μmol L-1 stimulated the length and dry mass of rice seedlings of both cultivars under saline conditions which appear to be associated with increased photosynthesis, gas exchange and chlorophyll concentration.
Key words: brassinosteroids, chlorophylls, Oryza sativa, salinity.
INTRODUCCIÓN
La salinidad es una de las causas de estrés más acuciantes en la actualidad. La inadecuada irrigación de los suelos, así como el cambio climático hacen que este fenómeno alcance un nivel global (1). En Cuba, la superficie agrícola está afectada en un 14 % y otro 15 % más presenta peligros potenciales de salinizaciónA.
Para enfrentar esta problemática, se trabaja en la búsqueda de soluciones que permitan incrementar la productividad de los cultivos en estas condiciones, para lo cual no sólo se pudieran utilizar cultivares tolerantes; sino también, trabajar en la introducción de tecnologías que permitan minimizar los daños que estos tipos de estrés provocan en las plantas (2).
Una de las tecnologías para incrementar los rendimientos agrícolas que se desarrolla en los últimos años, se basa en el uso de productos naturales inocuos desde el punto de vista ecológico, como los brasinoesteroides (BRs) (3).
Estos compuestos son potentes reguladores del crecimiento vegetal de naturaleza esteroidal. Estas hormonas tienen efectos pleiotrópicos como la estimulación del alargamiento celular y desdiferenciación de protoplastos, regeneración de la pared celular, regulación de la diferenciación de elementos traquearios e incremento de la biomasa y del rendimiento (4). En los últimos años, varios autores han estudiado la protección que los BRs le confieren a las plantas ante determinados tipos de estrés tanto bióticos como abióticos (5-8).
Sin embargo, hasta donde se conoce, no se ha logrado la optimización de la aplicación de estos compuestos en la protección de los cultivos sometidos a diferentes estreses ambientales, debido fundamentalmente, a que se desconoce el mecanismo en que ellos actúan en las plantas en estas condiciones. El conocimiento del modo de acción de los brasinoesteroides en el cultivo del arroz ante condiciones de estrés salino permitiría, en un futuro, una aplicación más efectiva y racional de estos reguladores del crecimiento. Por esta razón, el objetivo de este trabajo fue determinar si la aspersión foliar con 24-epibrasinólida es capaz de proteger a plantas jóvenes de arroz de los efectos adversos que provoca el estrés salino, así como, evaluar algunos indicadores fisiológicos asociados a esta respuesta.
MATERIALES Y MÉTODOS
Los experimentos se ejecutaron en el laboratorio de Biología Molecular de la Facultad de Biología de la Universidad de Alberta en Canadá. Se utilizaron semillas de arroz (Oryza sativa L.) de los cultivares Jucarito-104 (J-104), susceptible al estrés salinoB y Ginés, tolerante (9). Las semillas se desinfectaron con hipoclorito de sodio comercial durante cinco minutos; posteriormente, se lavaron seis veces con agua destilada estéril y luego se sumergieron en agua durante 24 horas.
Transcurrido este tiempo, se colocaron en placas de Petri, a temperatura ambiente en la oscuridad, para propiciar la germinación. Luego de 48 horas, las semillas germinadas se trasplantaron a macetas plásticas de 250 mL que contenían vermiculita.
Se utilizaron 60 macetas en total (cada una con cuatro plantas en crecimiento) y se colocaron en bandejas plásticas que contenían solución nutritiva de Hoagland diluida 1:1 v/v. Las bandejas se ubicaron dentro de una cámara de crecimiento con condiciones controladas de temperatura (28 ºC), humedad (70 %) y fotoperíodo (12 horas luz/12horas de oscuridad).
En el momento de emergencia de la tercera hoja verdadera, se realizó la aspersión foliar con 24-epibrasinólida a la concentración de 2 μmol L-1. Se asperjó un volumen total de 60 mL para 120 plantas. A todas las soluciones se le adicionó Tween 20 al 0,01 % como detergente. Las plantas se dividieron en dos grupos, uno se colocó en bandejas a las cuales se les adicionó solución nutritiva Hoagland diluida 1:1 v/v (SN) y el otro se colocó en bandejas que contenían solución nutritiva Hoagland diluida suplementada con NaCl 100 mmol L-1 (SN+NaCl).
A los siete días, las plantas que se sometieron a tratamiento salino, se colocaron en solución nutritiva Hoagland diluida 1:1 v/v, durante 14 días, para su recuperación.
Las evaluaciones de crecimiento (longitud de la parte aérea, longitud de las raíces, masa seca de la parte aérea y de las raíces) se realizaron a 16 plantas por tratamiento, a los tres y siete días del tratamiento salino y a los siete y 14 días de recuperación. En ese mismo momento se realizaron evaluaciones de la tasa fotosintética, la conductancia estomática, la transpiración y la concentración interna de CO2, utilizando un equipo portátil de medición de fotosíntesis (LI-6400; LI-COR, Lincoln, NE, USA). Para la determinación de las clorofilas a, b, totales y los carotenos se tomaron cuatro muestras por tratamiento y se realizó la extracción con acetona al 80 % durante 24 h en la oscuridad (10). Transcurrido este tiempo se midieron las absorbancias a las siguientes longitudes de onda 440, 649 y 665 nm.
En el experimento se utilizó un diseño completamente aleatorizado. Luego de comprobar que los datos obtenidos cumplieron con los supuestos teóricos de normalidad y homogeneidad de varianza, se les realizó un ANOVA de clasificación simple. En los casos en que hubo diferencias significativas entre tratamientos se realizó una comparación de medias mediante la prueba de Tukey con p≤0,05 (11), utilizando el programa estadístico STATGRAPHICS Plus 5.1 (12).
RESULTADOS Y DISCUSIÓN
En la Figura 1 se muestra el comportamiento de la longitud de la parte aérea de las plántulas. Como se puede observar, el tratamiento con NaCl afectó este indicador desde los tres días de estrés en ambos cultivares, mientras que la aspersión foliar con EBL logró revertir esta disminución en todos los momentos evaluados, siendo más efectiva en el cultivar J-104, donde las plantas tratadas con EBL y sometidas a estrés lograron valores semejantes al control sin sal, excepto a los 14 días de la recuperación.
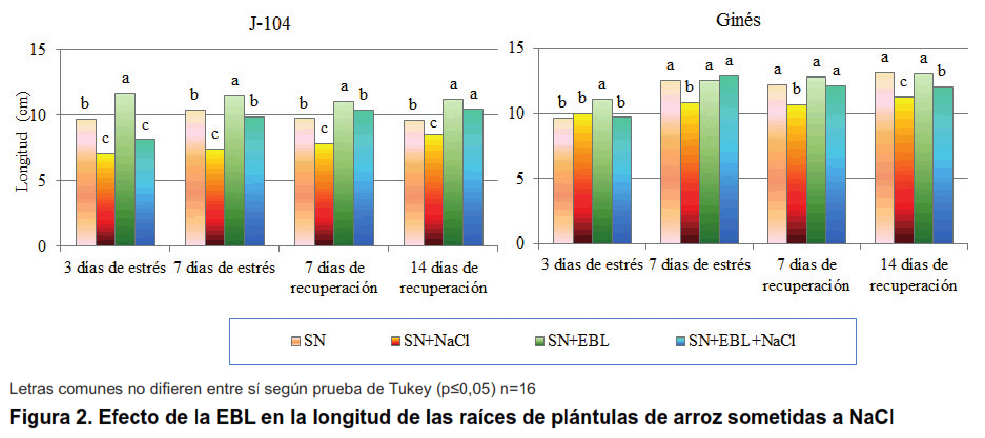
En la Figura 2 se observa el comportamiento de la longitud de las raíces en ambos cultivares. Es interesante destacar como la salinidad afectó al cultivar J-104 desde los tres días de estrés, mientras que en el cultivar Ginés esta afectación se hizo visible a los siete días de estrés. La aspersión foliar con EBL tuvo una influencia positiva en todos los momentos evaluados (Figura 2), destacándose el cultivar Ginés donde recuperó totalmente la longitud radical, excepto a los 14 días de recuperación.
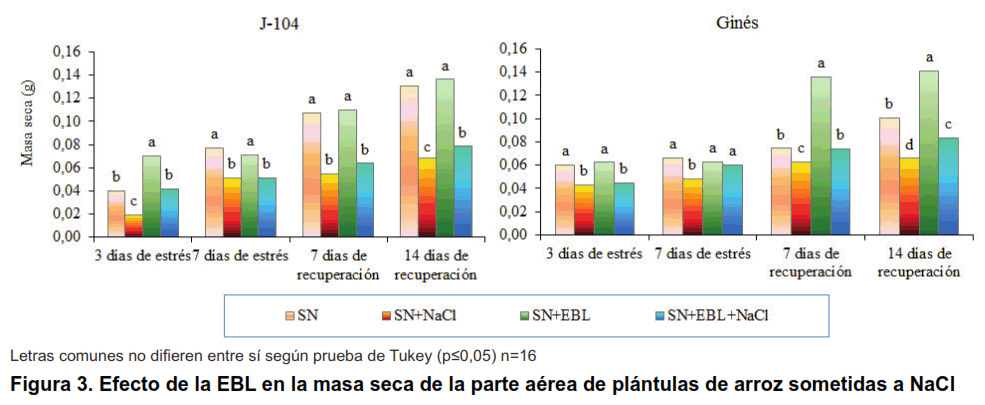
La masa seca de la parte aérea disminuyó significativamente desde los tres días del tratamiento con NaCl, siendo este efecto más marcado en el cultivar J-104 que en el Ginés (Figura 3). La aspersión foliar con EBL en el caso del cultivar J-104 solo logró revertir este efecto negativo a los tres días de estrés y a los 14 días de recuperación; sin embargo, en el cultivar Ginés la aspersión foliar (SN+EBL+NaCl) logró aumentar significativamente la masa seca de la parte aérea, en comparación con el tratamiento control (SN+NaCl), desde los siete días de estrés, manteniéndose este comportamiento hasta el fin del experimento.
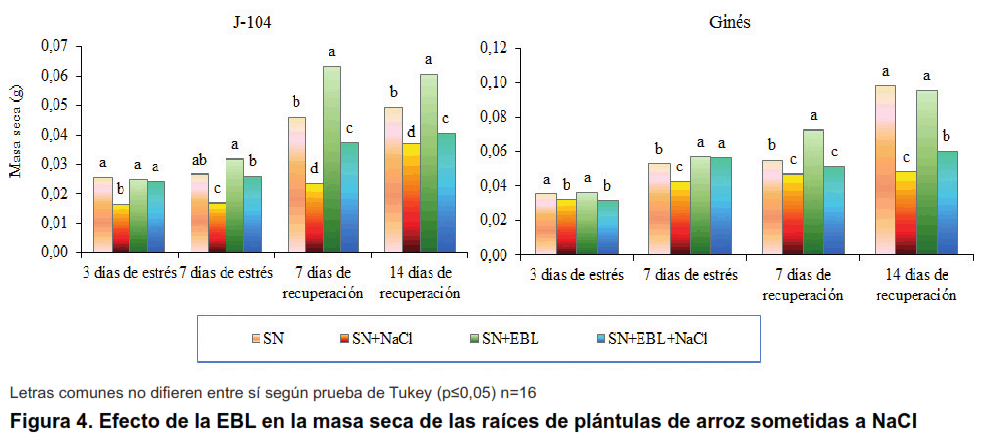
Al evaluar la masa seca de las raíces se observa que el estrés salino provocó una disminución significativa que fue revertida en las plantas asperjadas con EBL pero solamente en el cultivar J-104. El comportamiento en el cultivar Ginés fue diferente ya que la aspersión foliar solo pudo mejorar este indicador a los siete días de estrés y a los 14 días de recuperación (Figura 4).
La salinidad afectó negativamente los indicadores de crecimiento en ambos cultivares, incluso durante la recuperación estos efectos se hicieron visibles. Es bien conocido que la salinidad inhibe el crecimiento de las plantas y en muchos casos la parte aérea es más afectada que las raíces. Este efecto del estrés salino en el crecimiento es ampliamente conocido, tanto el estrés hídrico como la acumulación de iones retrasan los procesos de división y diferenciación celular (13). Por ejemplo, en cuatro cultivares de cítricos sometidos a diferentes niveles de salinidad se afectaron tanto la longitud como las masas secas de raíces y vástagos, siendo este efecto más marcado en los cultivares sensibles (14). Igualmente, se ha observado la disminución de la longitud y la masa fresca de plántulas de mostaza sometidas a diferentes concentraciones de NaCl (15). Además, en Fraxinus ornus, disminuyó la masa seca de las raíces en plantas expuestas a salinidad (16).
Por otro lado, la disminución en el crecimiento de las plantas tratadas, durante la recuperación, pudiera deberse a que los efectos tóxicos de la acumulación de sales tardan más en manifestarse pero permanecen en el tiempo, aún después de pasar al período de recuperación. Para corroborar esta hipótesis, se ha señalado el hecho de que una vez concluido el período de estrés y sometidas las plantas durante siete días al tratamiento control, estas no lograron recuperarse, lo que demuestra el carácter irreversible de los daños ocasionados (17).
Es de destacar, como el cultivar Ginés considerado tolerante no se comportó como tal, teniendo reducciones del crecimiento similares al cultivar J-104. Esto pudiera deberse a la concentración de sal usada en el experimento (100 mmol L-1), equivalente a 9,867dS m-1, considerada por las normas cubanas de salinidad como muy fuertemente salino (18), lo que ocasionaría daños severos a las plantas. Otra explicación pudiera ser que los mecanismos de tolerancia de este cultivar se desarrollan en etapas posteriores del crecimiento y esto permite obtener un mejor rendimiento, aún cuando en las primeras etapas se comporte de esta manera. De igual manera, otros trabajos donde se utilizaron los cultivares de arroz J-104 e INCA LP-7, que, a nivel de campo, responden de manera diferente al estrés salino (19), no resultaron muy contrastantes en sus respuestas a la concentración de 100 mM de NaCl en la fase inicial de crecimiento (7 días)(20).
Por otra parte, la aspersión foliar con EBL logró revertir los efectos adversos del estrés salino en el crecimiento de los dos cultivares estudiados.
Resultados similares han informado otros autores, quienes observaron una recuperación en el crecimiento de las plántulas de arroz var. IR-64 cuyas semillas fueron tratadas durante 24 h con NaCl (150 mmol L-1) y EBL (3 µmol L-1) (21).
Por otra parte, se informó el aumento en el crecimiento inicial de plántulas de arroz cultivar IR-28 ante estrés salino (120 mmol L-1) tratadas con EBL (3 µmol L-1)(22). Además, se ha observado una recuperación en el crecimiento en plantas de arroz, asperjadas durante la fase de inicio de paniculación y sometidas a estrés salino (100 mmol L-1) (23).
Más recientemente, se evaluó la influencia del tratamiento a las semillas con varias concentraciones de 24-epibrasinólida y Biobras-16 (BB-16), un análogo de brasinoesteroides, en el crecimiento de las plántulas de dos cultivares de arroz (J-104 y Ginés) en medio salino. Tanto la EBL como el BB-16 fueron capaces de revertir parcialmente la inhibición que en el crecimiento de las plántulas de ambos genotipos de arroz provocó la presencia de NaCl; siendo más efectivos los tratamientos en el cultivar tolerante Ginés (24).
En otros cultivos la EBL también ha sido capaz de incrementar el crecimiento en condiciones de estrés salino como en Vigna radiata (25), Lactuca sativa L. (26) y Triticum aestivum L. (27).
El efecto osmótico del estrés salino produce el cierre de los estomas con la consecuente disminución de la fotosíntesis por deficiencia de CO2. Por esa razón en el experimento se evaluó la tasa fotosintética, la conductancia estomática, la transpiración y la concentración interna de CO2.
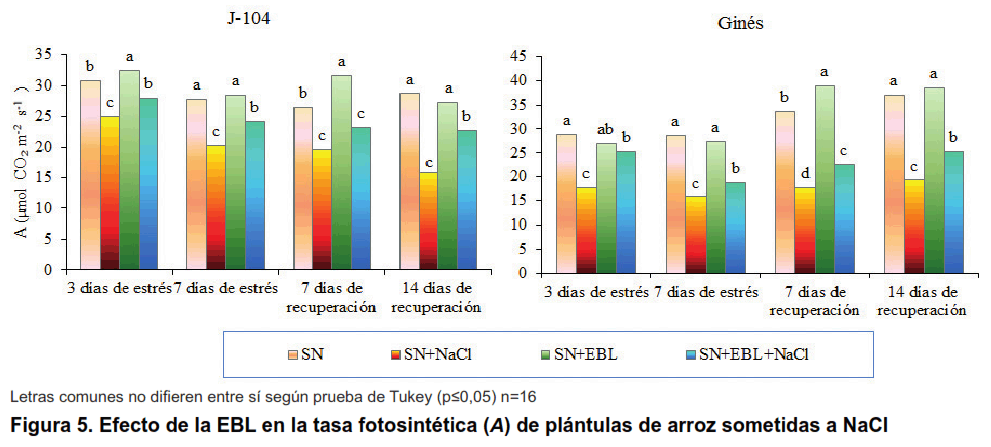
Como se puede observar en la Figura 5 la tasa fotosintética de las plantas disminuye con el estrés salino en ambos cultivares desde los tres días de impuesto el mismo y se mantiene este comportamiento durante todo el experimento, por lo que en la recuperación no se logra revertir este efecto. Algo similar ha sido informado por otros autores que refieren la disminución de la fotosíntesis en cuatro cultivares de cítricos ante diferentes concentraciones de NaCl, siendo los más afectados los cultivares sensibles (14). Igualmente, se ha observado la disminución de la tasa fotosintética en plántulas de mostaza sometidas a diferentes concentraciones de NaCl (15). También, en Fraxinus ornus disminuyó la tasa fotosintética en plantas expuestas a salinidad (16).
La aspersión foliar con EBL logró revertir parcialmente el efecto del estrés salino en la fotosíntesis, destacándose el cultivar Ginés donde se observa en todos los momentos evaluados, no siendo así en el cultivar J-104.
Otros autores han informado la influencia positiva del tratamiento con brasinoesteroides en la fotosíntesis, por ejemplo, en berenjena la concentración de 100 nmol L-1 de 24-EBL logró revertir los efectos inhibitorios del estrés salino en la fotosíntesis (28). También en frijol mungo la aspersión foliar de EBL (10-8 mol L-1) aumentó la fotosíntesis en el tratamiento con NaCl a niveles similares al control (25). Además otros brasinoesteroides naturales como la 28-homobrasinólida (28-HBL) también han tenido efectos positivos en la fotosíntesis en condiciones de estrés. Por ejemplo, en Brassica juncea la concentración de 10-6 mol L-1 de 28-HBL fue la de mejor resultado en el incremento de la fotosíntesis en las tres concentraciones de NaCl evaluadas (50,100 y 150 mmol L-1) (29).
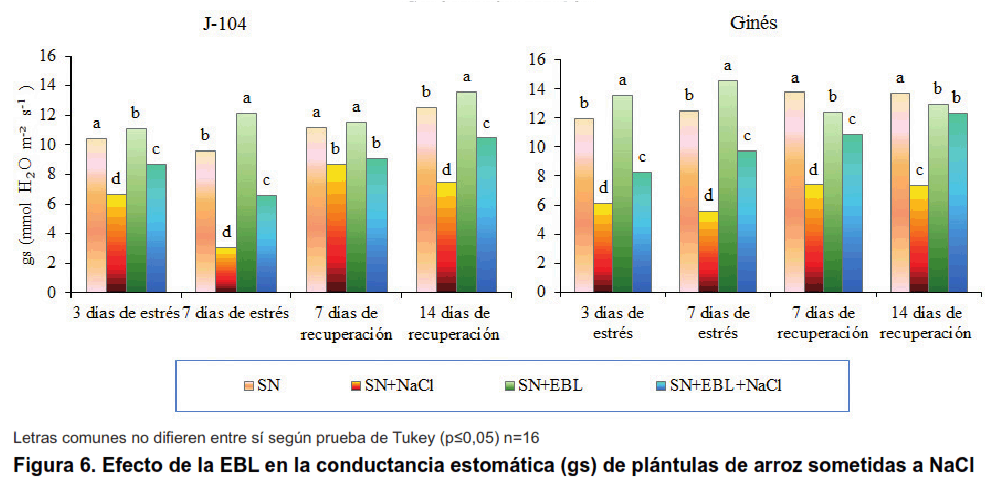
Otro de los indicadores evaluados en el experimento fue la conductancia estomática que es una medida de la apertura de los estomas para la realización del intercambio gaseoso. La regulación de la conductancia estomática en las hojas es un fenómeno clave tanto para la absorción de CO2 como para evitar la desecación en condiciones de estrés (30).
En la Figura 6, se observa que el estrés salino disminuyó significativamente la conductancia estomática en ambos cultivares, siendo este comportamiento más pronunciado en el cultivar J-104 a los siete días de estrés. Resultados similares han sido demostrados por otros autores en cuatro cultivares de cítricos ante diferentes concentraciones de NaCl, siendo los cultivares más afectados los sensibles. Igualmente, se ha observado la disminución de la conductancia estomática en plántulas de mostaza sometidas a diferentes concentraciones de NaCl. También, en Fraxinus ornus disminuyó la conductancia estomática en plantas expuestas a la salinidad (14-16).
La aspersión foliar con EBL logró revertir este efecto, incrementando significativamente la conductancia estomática en ambos cultivares, en casi todos los momentos evaluados, excepto a los siete días de recuperación en el cultivar J-104.
Otros autores han obtenido incrementos en la conductancia estomática con la utilización de la EBL, por ejemplo en lechuga el tratamiento a la semilla y la posterior aspersión foliar de tres concentraciones de EBL (1, 2 y 3 µmol L-1) incrementó la conductancia estomática en un 75 % respecto al control, siendo el tratamiento más efectivo el de 3 µmolL-1 tanto a la concentración de NaCl de 50 mmol L-1 como a la de 100 mmol L-1 (26). También en frijol mungo la aspersión foliar de EBL (10-8 mol L-1) aumentó la conductancia estomática en las plantas tratadas con NaCl a niveles similares al control (25). En berenjena, las concentraciones de EBL de 50 y 100 nmol L-1 lograron revertir la disminución provocada por NaCl a la concentración de 90 mmol L-1; sin embargo, las concentraciones de EBL de 200 y 400 nmol L-1 potenciaron el efecto adverso de la salinidad (28).
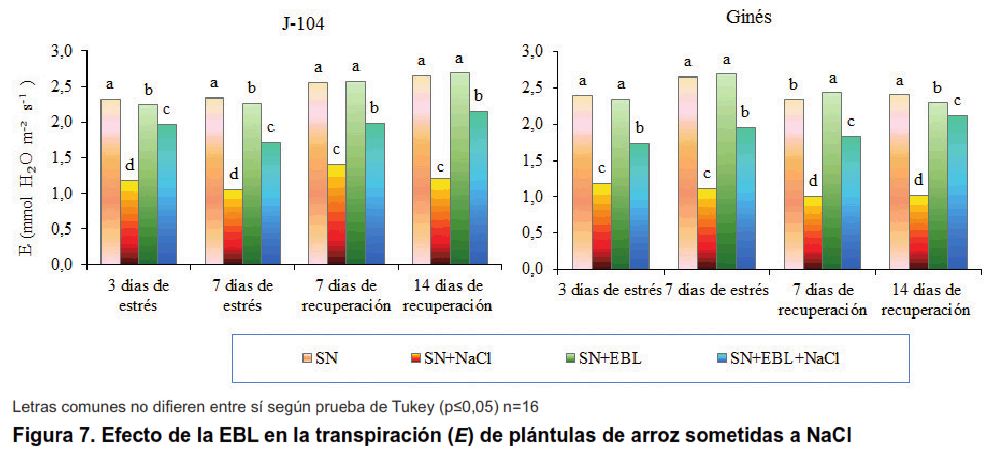
El estrés salino disminuyó significativamente la transpiración en ambos cultivares, manteniéndose este comportamiento durante la recuperación (Figura 7). Algo similar se obtuvo en cuatro cultivares de cítricos ante diferentes concentraciones de NaCl, disminuyendo la transpiración más en los cultivares sensibles que en los tolerantes (14). Igualmente, se ha observado la disminución de la transpiración en plántulas de mostaza sometidas a diferentes concentraciones de NaCl (15). También en Fraxinus ornus disminuyó este indicador en plantas expuestas a salinidad (16).
La aspersión foliar con EBL logró revertir este efecto, incrementando significativamente la transpiración, en este caso no se encontraron diferencias entre las plantas de los dos cultivares estudiados, en ninguno de los momentos evaluados.
Este comportamiento fue similar al encontrado en melón de agua, donde la aspersión foliar con EBL (10-6 mol L-1) revirtió la disminución en la transpiración provocada por el tratamiento con agua salinizada (70 mmol L-1 de NaCl) (31). Igualmente, en uva, la aplicación de EBL (3 y 6 µmol L-1) fue capaz de restituir la transpiración de plantas estresadas con varios niveles de NaCl (25, 50, 75 y 100 mmol L-1) (32).
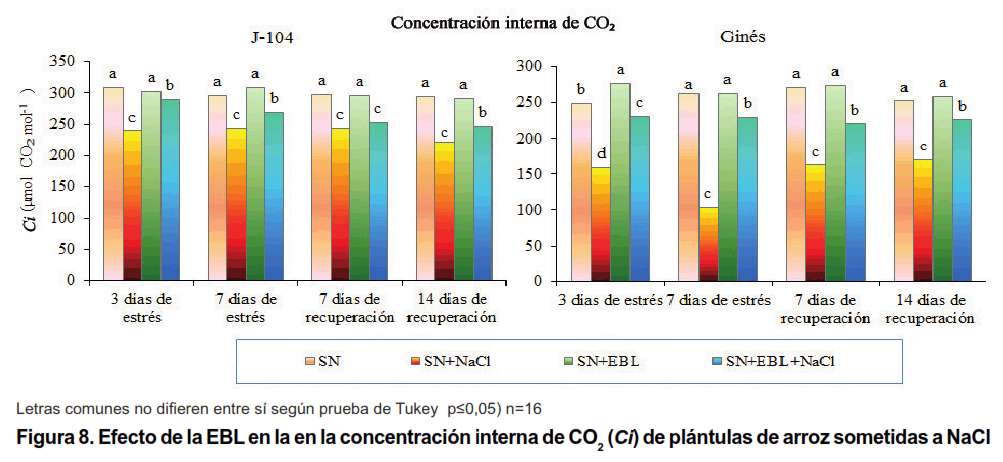
Como se puede apreciar, en la Figura 8, el estrés salino disminuyó significativamente la concentración interna de CO2 en ambos cultivares, siendo este efecto más marcado en el cultivar Ginés. Durante la recuperación se mantuvo este comportamiento. Otros autores han informado la disminución de la Ci en diferentes cultivares de arroz expuestos a estrés salino durante siete días (33).
La aspersión foliar con EBL revirtió este efecto negativo de la sal, particularmente, en el cultivar Ginés, donde hubo un mayor incremento de la Ci en todos los momentos evaluados, mientras que en el cultivar J-104 el incremento fue menor y a los siete días de recuperación no se encontraron diferencias significativas.
Este resultado concuerda con lo informado por otros autores, que observaron un aumento de la Ci en plantas de frijol mungo asperjadas con EBL (10-8 mol L-1) y sometidas a estrés por NaCl (25). También, en berenjena, las concentraciones de EBL de 50 y 100 nmol L-1 lograron revertir la disminución en la Ci provocada por NaCl a la concentración de 90 mmol L-1; sin embargo, las concentraciones de EBL de 200 y 400 nmol L-1 no tuvieron diferencias con el control salinizado (28). Por otra parte, en trigo se observó muy poco incremento de este indicador en las plántulas estresadas con varias concentraciones de EBL (0,052; 0,104; 0,156 µmol L-1) (34).
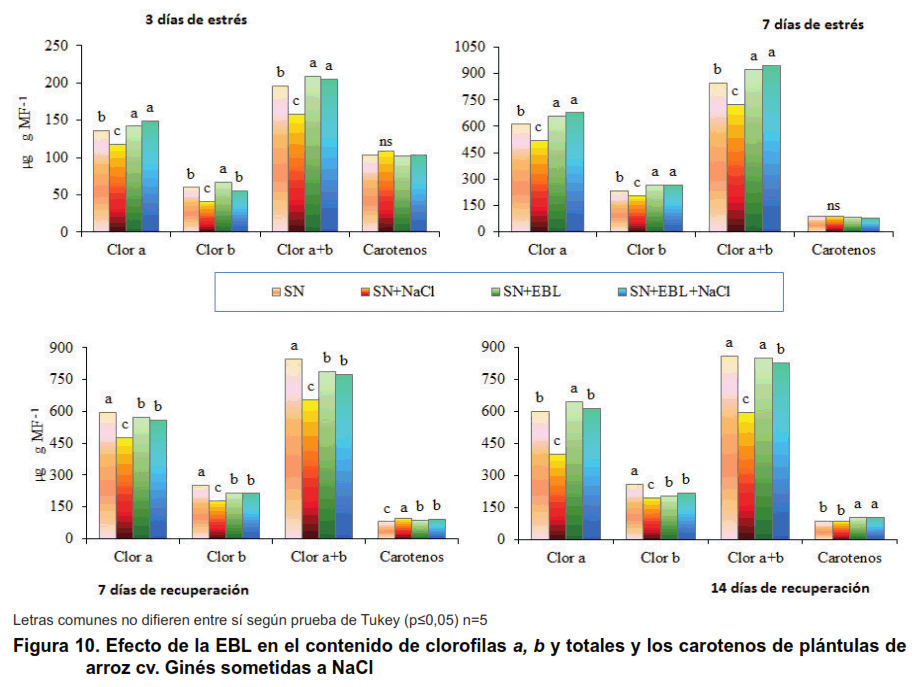
Al analizar los resultados sobre los pigmentos clorofílicos en el cultivar J-104, a los tres días del tratamiento con NaCl, se observó una disminución de la clorofila a y de las clorofilas totales en las plántulas estresadas; al finalizar el periodo de estrés esta disminución alcanzó también la clorofila b y los carotenos. En la recuperación, este comportamiento se mantuvo en todos los pigmentos evaluados (Figura 9). Sin embargo, en el cultivar Ginés, la disminución de las clorofilas comenzó desde los tres días de estrés hasta el final del experimento, mientras que los carotenos no sufrieron cambios durante el periodo de estrés, pero disminuyeron en la recuperación (Figura 10).
Este resultado concuerda con diferentes autores que han observado la disminución de la concentración de este pigmento ante estrés salino en numerosos cultivos como: albahaca, arroz, uva y tomate (32,35-37).
La reducción en el contenido de clorofilas en las hojas en condiciones de estrés salino ha sido atribuida a varias causas: la destrucción de los pigmentos clorofílicos por el incremento de la actividad clorofilasa y la supresión de enzimas de la biosíntesis como la porfobilinógeno desaminasa. También puede estar ocasionada por la interferencia de los iones salinos con la síntesis de novo de proteínas y componentes estructurales de la clorofila (30).
La aspersión foliar con EBL logró revertir los efectos adversos del estrés salino en la concentración de clorofilas desde los tres días de estrés hasta el final del experimento en ambos cultivares.
En cuanto a los carotenos la aspersión foliar de EBL aumentó la concentración de los mismos en las plántulas estresadas en el cultivar J-104 en todos los momentos evaluados. En el cultivar Ginés, la aspersión foliar logró aumentar la concentración de carotenos durante la recuperación.
La estimulación de la concentración de clorofilas en las hojas de las plantas asperjadas con EBL y sometidas al tratamiento con NaCl, confirma lo obtenido por otros autores quienes han informado que la aspersión foliar con EBL a varias concentraciones (50, 100, 200 y 400 nmol L-1) disminuyó la pérdida de los pigmentos fotosintéticos provocada por el estrés salino (NaCl 90 mmol L-1) en plántulas de berenjena (28).También, en Phaseolus vulgaris y Hordeum vulgare el pre-tratamiento a las semillas con brasinoesteroide (5 µmol L-1) revirtió la disminución del contenido de clorofila provocado por el estrés salino (150 mmol L-1), alcanzando niveles similares a las plantas controles (38). De igual forma, la aspersión foliar de EBL en plantas de frijol sometidas a estrés salino revirtió la disminución en las clorofilas provocadas por este estrés (39).
También se ha demostrado el efecto de otros BRs en la restitución de las moléculas de clorofila ante estrés salino. Por ejemplo, en Brassica juncea la concentración de 10-6 mol L-1 de 28-HBL fue la de mejor resultado en el incremento de la concentración de clorofilas en las tres concentraciones de NaCl evaluadas (50,100 y 150 mmol L-1) (29).
De forma general, el cultivar Ginés respondió mejor a la aplicación de la EBL. Estos resultados concuerdan con estudios anteriores donde se utilizaron estos mismos cultivares y la respuesta del cultivar Ginés a la aplicación de EBL a las semillas fue superior a la del cultivar J-104 (24). Por otra parte, varios investigadores han informadoque la respuesta de las plantas a la aplicación exógena de los brasinoesteroides depende de la sensibilidad de las mismas a determinado tipo de estrés; respondiendo mejor los cultivares tolerantes que los sensibles (27,32).
La reducción en el crecimiento en las plántulas estresadas en el presente trabajo, al parecer es debido a una disminución de la fotosíntesis, lo que pudiera deberse a factores estomáticos ya que se afectaron la conductancia estomática y la transpiración y por tanto la Ci disponible para la asimilación de CO2. Además, el trabajo presenta evidencias de que esta disminución de la fotosíntesis pudiera estar asociada también a factores no estomáticos puesto que las clorofilas también disminuyeron, tanto la a como la b, lo que podría dificultar la absorción de luz.
El efecto positivo de la aspersión foliar de EBL en las variables del crecimiento de las plántulas estresadas, al parecer se debe a un incremento de la conductancia estomática, la transpiración y la Ci, lo que aumentaría la tasa fotosintética y la producción de biomasa. Igualmente la restitución de las moléculas de clorofila por el tratamiento con EBL pudiera explicar el aumento de la fotosíntesis.
CONCLUSIONES
La aspersión foliar con EBL estimula el crecimiento de las plantas de arroz sometidas a estrés salino, lo que pudiera estar asociado a un incremento de la tasa fotosintética, la conductancia estomática, la transpiración y la concentración interna de CO2, así como al aumento en la concentración de clorofilas. El cultivar Ginés respondió mejor a la aplicación exógena de la EBL, que el cultivar J-104.
AGRADECIMIENTOS
El colectivo de autores quiere agradecer a la beca Emerging Leaders in Americas Program (ELAP) otorgada por el gobierno de Canadá, por el financiamiento para realizar la estancia de investigación en la Universidad de Alberta.
NOTAS AL PIE
AOrellana R, Febles J, Ortega F, et al. GEO Cuba. Evaluación del medio ambiente cubano. Santo Domingo. República Dominicana: Centenario SA; 2009. 293 p.
BAlfonso, R. Determinación de parámetros genético-fisiológicos indicadores del estrés hídrico para su empleo en el mejoramiento genético del arroz (Oryza sativa L.) y la estabilidad varietal [Tesis de Doctorado]. IIA; 1998.
BIBLIOGRAFÍA
1. FAO. FAO Land and Plant Nutrition Management Service[Internet]; 2016. [cited 2016 Sep 7] Available from: http://www.fao.org/ag/agl/agll/spush
2. Kumar K, Kumar M, Kim S-R, Ryu H, Cho Y-G. Insights into genomics of salt stress response in rice. Rice. 2013;6(27):1-15. doi: 10.1186/1939-8433-6-27.
3. Núñez VM, Reyes GY, Rosabal AL, Martínez GL. Análogos espirostánicos de brasinoesteroides y sus potencialidades de uso en la agricultura. Cultivos Tropicales. 2014;35(2):34-42.
4. Fariduddin Q, Yusuf M, Ahmad I, Ahmad A. Brassinosteroids and their role in response of plants to abiotic stresses. Biologia Plantarum. 2014;58(1):9-17. doi: 10.1007/s10535-013-0374-5.
5. Behnamnia M. Protective Roles of Brassinolide on Tomato Seedlings under Drought Stress. International Journal of Agriculture and Crop Sciences. 2015;8(3):455-62.
6. Swamy KN, Vardhini BV, Ramakrishna B, Anuradha S, Siddulu N, Rao SSR. Role of 28-homobrassinolide on growth biochemical parameters of Trigonella foenu-graecum L. plants subjected to lead toxicity. International Journal of Multidisciplinary and Current Research. 2014;2:317-21.
7. Jiang W, Bai J, Yang X, Yu H, Liu Y. Exogenous Application of Abscisic Acid, Putrescine, or 24-Epibrassinolide at Appropriate Concentrations Effectively Alleviate Damage to Tomato Seedlings from Suboptimal Temperature Stress. HortTechnology. 2012;22(1):137-44.
8. Kumar S, Sirhindi G, Bhardwaj R. 28-Homobrassinolide-Induced Exaggerated Growth, Biochemical Molecular Aspects of Brassica Juncea L. RLM-619 Seedlings under High Temperature Stress. J Plant Biochem Physiol. 2014;2(127):1-10. doi: 10.4172/2329-9029.1000127.
9. González MC, Pérez N, Cristo E. Gines: primer mutante de arroz obtenido a partir de la irradiación con protones. Cultivos Tropicales. 2009;30(3):59.
10. Arnon DI. Copper enzyme in isolated chloroplasts: Polyphenol oxidase in Beta vulgaris. Plant Physiology. 1949;24:1-15.
11. Tukey JW. Bias and confidence in not quite large samples. The Annals of Mathematical Statistics. 1958;29(2):614–23.
12. Statistical Graphics Crop. STATGRAPHICS® Plus [Internet]. 2000. (Profesional). Available from: http://www.statgraphics.com/statgraphics/statgraphics.nsf/pd/pdpricing
13. Gupta B, Huang B. Mechanism of Salinity Tolerance in Plants: Physiological, Biochemical, and Molecular Characterization. International Journal of Genomics. 2014;1-18. doi: 10.1155/2014/701596.
14. Balal RM, Khan MM, Shahid MA, Mattson NS, Abbas T, Ashfaq M, Garcia-Sanchez F, Ghazanfer U, Gimeno V, Iqbal Z. Comparative Studies on the Physiobiochemical, Enzymatic, and Ionic Modifications in Salt-tolerant and Salt-sensitive Citrus Rootstocks under NaCl Stress. J. Amer. Soc. Hort. Sci. 2012;137(2):86-95.
15. Jamil M, Rha ES. NaCl Stress-Induced Reduction in Growth, Photosynthesis and Protein in Mustard. Journal of Agricultural Science. 2013;5(9):114-27. doi: 10.5539/jas.v5n9p114.
16. Fini A, Guidi L, Giordano C, Baratto MC, Ferrini F, Brunetti C, Calamai L, Tattini M. Salinity stress constrains photosynthesis in Fraxinus ornus more when growing in partial shading than in full sunlight: consequences for the antioxidant defence system. Annals of Botany. 2014;114:525-38. doi: 10.1093/aob/mcu130.
17. Morales D, Bolarín MC, Cayuela E. Respuesta de plantas de arroz (Oryza sativa L.) a la aplicación de diferentes niveles de NaCl. I. Crecimiento y relaciones hídricas. Cultivos Tropicales. 2006;27(4):27-32.
18. Oficina Nacional de Normalización: Calidad del suelo - Evaluación de la afectación por salinidad. La Habana, Cuba; 2010.
19. González MC, Cristo E, Pérez N, Delgado P. INCA LP-7, nueva variedad de arroz para suelos afectados por la salinidad. Cultivos Tropicales. 2002;23(3):89.
20. Núñez M, Mazorra LM, Martínez L, González MC, Robaina C. Análogos de brasinoesteroides revierten parcialmente el impacto del estrés salino en el crecimiento inicial de las plántulas de dos genotipos de arroz (Oryza sativa L.). Cultivos Tropicales. 2007;28(2):95-9.
21. Anuradha S, Rao S. Effect of brassinosteroids on salinity stress induced inhibition of seed germination and seedling growth of rice (Oryza sativa L.). Plant Growth Regulation. 2001;33(2):151-3. doi: 10.1023/A:1017590108484.
22. Özdemir F, Bor M, Demiral T, Turkan I. Effects of 24-epibrassinolide on seed germination, seedling growth, lipid peroxidation, proline content and antioxidative system of rice (Oryza sativa L.) under salinity stress. Plant Growth Regulation. 2004;42(3):203-11. doi: 10.1023/B:GROW.0000026509.25995.13.
23. Phap VA. Induction of salt tolerance in rice (Oryza sativa L.) by brassinosteroids. Cantho, Vietnam: Institut für Molekulare Physiologie und Biotechnologie der Pflanzen - der Universität Bonn; 2006. 100 p.
24. Núñez M, Reyes Y, Rosabal L, Martínez L, González MC, Pieters A. Brasinoesteroides y sus análogos estimulan el crecimiento de plántulas de dos genotipos de arroz (Oryza sativa L.) en medio salino. Cultivos Tropicales. 2013;34(1):74-80.
25. Mir BA, Khan TA, Fariduddin Q. 24-epibrassinolide and spermidine modulate photosynthesis and antioxidant systems in Vigna radiata under salt and zinc stress. International Journal of Advanced Research. 2015;3(5):592-608.
26. Ekinci M, Yildirim E, Dursun A. Mitigation of salt stress in lettuce (Lactuca sativa L. var. Crispa) by seed and foliar 24-epibrassinolide treatments. HortScience. 2012;47(5):631-6.
27. Talaat NB, Shawky BT. 24-Epibrassinolide alleviates salt-induced inhibition of productivity by increasing nutrients and compatible solutes accumulation and enhancing antioxidant system in wheat (Triticum aestivum L.). Acta Physiologiae Plantarum. 2013;35:729-40.
28. Wu X-X, Ding H-D, Zhu Z-W, Yang S-J, Zha D-S. Effects of 24-epibrassinolide on photosynthesis of eggplant (Solanum melongena L.) seedlings under salt stress. African Journal of Biotechnology. 2012;11(35):8665-71. doi: 10.5897/AJB11.3416
29. Alyemeni MN, Hayat S, Wijaya L, Anaj A. Foliar application of 28-homobrassinolide mitigates salinity stress by increasing the efficiency of photosynthesis in Brassica juncea. Acta Botanica Brasilica. 2013;27(3):502-5.
30. Ashraf M, Harris PJC. Photosynthesis under stressful environments: An overview. Photosynthetica. 2013;51(2):163-90. doi: 10.1007/s11099-013-0021-6.
31. Cheng W, Huang Y, Meng C, Zhang N, Zeng H, Ren J, et al. Effect of Exogenous 24-Epibrassinolide on Salt Resistance of watermelon (Citrullus lanatus L.) under Salinity Stress. In: V International Conference on Advanced Design and Manufacturing Engineering [Internet]. Atlantis Press; 2015 [cited 2017 May 7]. Available from: http://atlantis-press.com/php/paper-details.php?id=25840093
32. Seif SN, Tafazzoli E, Talai, A-R, Aboutalebi A, Abdosi V. Evaluation of two grape cultivars (Vitis vinifera L.) against salinity stress and surveying the effect of methyl jasmonate and epibrassinolide on alleviation the salinity stress. International Journal of Biosciences. 2014;5(7):116-25. doi: 10.12692/ijb/5.7.116-125.
33. Moradi F, Ismail AM. Responses of photosynthesis, chlorophyll fluorescence and ROS-scavenging systems to salt stress during seedling and reproductive stages in rice. Annals of Botany. 2007;99:1161-73. doi: 10.1093/aob/mcm052.
34. Ali Q, Atar H-R, Ashraf M. Modulation of growth, photosynthetic capacity and water relations in salt stressed wheat plants by exogenously applied 24-epibrassinolide. Plant Growth Regulation. 2008;56:107-16. doi: 10.1007/s10725-008-9290-7.
35. Heidari M. Effects of salinity stress on growth, chlorophyll content and osmotic components of two basil (Ocimum basilicum L.) genotypes. African Journal of Biotechnology. 2012;11(2):379-84. doi: 10.5897/AJB11.2572.
36. Senguttuvel P, Vijayalakshmi C, Thiyagarajan K, Kannanbapu JR, Kota S, Padmavathi G, Geetha S, Sritharan N, Viraktamath BC. Changes in photosynthesis, chlorophyll fluorescence, gas exchange parameters and osmotic potentialto salt stress during early seedling stage in rice (Oryza sativa L.). SABRAO Journal of Breeding and Genetics. 2014;46(1):120-35.
37. Furdi F, Velicevici G, Petolescu C, Popescu S. The effect of salt stress on chlorophyll content in several Romanian tomato varieties. Journal of Horticulture, Forestry and Biotechnology. 2013;17(1):363-7.
38. Ali AA, Abdel-Fattah RI. Osmolytes-antioxidant behaviour in Phaseolus vulgaris and Hordeum vulgare whit brassinosteroid under salt stress. Journal of Agronomy. 2006;5(1):167-74.
39. Rady MM. Effect of 24-epibrassinolide on growth, yield, antioxidant system and cadmium content of bean (Phaseolus vulgaris L.) plants under salinity and cadmium stress.Scientia Horticulturae. 2011;129:232-7. doi: 10.1016/j.scienta.2011.03.0.
Recibido: 05/12/2016
Aceptado: 14/03/2017
Ms.C. Yanelis Reyes, Instituto Nacional de Ciencias Agrícolas. Gaveta Postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32700. Email: yanelisrg@inca.edu.cu


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}