










 Servicios personalizados
Servicios personalizados
 texto en
texto en  Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
La simbiosis micorrízica arbuscular se presenta en cerca del 80 % de las especies vegetales, en las cuales se encuentran el grueso de los cultivos económicos 1 y son ampliamente reconocidos los beneficios que aportan a las plantas y al ambiente, asociados al incremento en la toma de nutrientes 2 y agua 3, al efecto de bioprotección frente al ataque de algunas plagas radicales y foliares 4, a la tolerancia a metales pesados 5, así como otros ecoservicios, entre los que se encuentran mejoras en los agregados del suelo, participación en los ciclos del Carbono y del Nitrógeno, potenciando una mayor resiliencia del ecosistema 6,7 y constituye un reto el manejo consciente de los hongos micorrízicos arbusculares (HMA), para optimizar estos beneficios en los agrosistemas 8,9.
Los trabajos desarrollados en Cuba, al respecto, se basan en la inoculación de cepas eficientes de HMA de carácter generalista con las especies vegetales y recomendadas en base al ambiente edáfico 10. De este modo se diseñan sistemas integrales de suministro de nutrientes, a partir de una micorrización efectiva de los diferentes cultivos, obteniendo altos rendimientos y satisfactorio estado nutricional, con menores dosis de fertilizantes 8,11-13.
Uno de los aspectos encontrados al trabajar con cultivos dependientes de la micorrización, ha sido la presencia de un positivo efecto de permanencia del inoculante aplicado a diferentes cultivos sobre el primer cultivo en sucesión, aunque desaparece para los cultivos posteriores 8,14. Este efecto se ha vinculado bien con el uso de los abonos verdes inoculados 15,16 o con el manejo de los inoculantes en secuencias de cultivos 14, con sustitución en ambos casos de la inoculación del primer cultivo en sucesión; aunque también ha sido evaluado en cultivos perennes inoculados 11,17, para definir el momento de la reinoculación. La desaparición del efecto de permanencia ha sido asociada en lo fundamental a la competencia con la micorriza residente 18, a las características del cultivo, su manejo agronómico y las condiciones edáficas 11,19.
Ipomea batata (L.) Lam es un cultivo con significativa respuesta a la inoculación, pero que aún utilizando la vía de recubrimiento de los esquejes 14 requiere de muy altas cantidades de inoculantes. Los trabajos desarrollados recientemente en este cultivo 20, establecieron que, utilizando como precedentes inoculados cultivos comunes en la agricultura cubana (una gramínea como maíz y leguminosas como fríjol común o Vigna unguiculata, sembrados en época lluviosa y poco lluviosa), además de los beneficios de la inoculación sobre estos precedentes, se encontraron efectos de permanencia igualmente efectivos con ambos precedentes y en las dos épocas. De este modo no es necesario inocular el boniato, con independencia de los valores mayores de esporas micorrízicas reproducidas por el maíz y por los cultivos sembrados en la época lluviosa.
En las anteriores publicaciones 14,16,20, el intervalo entre la cosecha de los precedentes inoculados y el cultivo en sucesión no fue mayor de 30 días. Por tanto, debido a la importancia práctica de los resultados reportados, a la posible influencia de las diferencias encontradas en la reproducción de esporas entre los precedentes y épocas en la duración del efecto y al hecho de que, tanto en las secuencias de cultivos 8,14, como en los cultivos perennes inoculados, la respuesta positiva a la inoculación desaparece gradualmente en el tiempo 11,17. Para definir el manejo de los precedentes inoculados en una u otra época, se consideró necesario, determinar cómo influían intervalos superiores a los 30 días sobre la efectividad del mismo.
MATERIALES Y MÉTODOS
La investigación se desarrolló en el Instituto de Investigaciones de Viandas Tropicales (INIVIT) ubicado a los 22o 35’ N, 80o18’ W y a 40 m s.n.m., en el municipio de Santo Domingo, provincia de Villa Clara, Cuba, sobre suelo Pardo argénico medianamente lavado 21, clasificado también como Cambisol eútrico sin carbonatos 22, durante los años 2015 y 2016.
El suelo en el área experimental fue homogéneo, sin presentar diferencias significativas (p<0,05) en las variables determinadas entre los diferentes experimentos, años y épocas de siembra. En la Tabla 1 se presentan los valores promedio correspondientes a cada época de siembra. El suelo presentó una reacción neutra, así como valores bajos de materia orgánica posiblemente asociados al cultivo continuo. Los contenidos de fósforo disponible fueron bajos y los de potasio fueron medios. El calcio y el magnesio intercambiable presentaron valores altos y típicos de estos suelos. Las esporas micorrízicas fueron bajas, con valores aproximados a los reportados en áreas colindantes con similares tipos de suelo y manejo 12,14.
Tabla 1 Algunas propiedades iniciales del suelo en el área experimental y en ambas épocas de siembra de los precedentes. Profundidad de muestreo de 0-20 cm
| Épocas de siembra de los precedentes | pH | Nitrógeno total | MO | P2O5 | K2O | Ca | Mg | Na | K | Esp./ 50 g |
|---|---|---|---|---|---|---|---|---|---|---|
| H2O | (g kg-1) | mg kg-1 | (cmolc kg-1) | |||||||
| Lluviosa | 7,12 | 1,60 | 20,02 | 20,20 | 21,44 | 30,20 | 4,29 | 0,37 | 0,61 | 41,5 |
| Z1-α * S x̅ | ±0,09 | ±0,07 | ±0,14 | ±0,14 | ±0,07 | ±0,38 | ±0,06 | ±0,02 | ±0,02 | ±0,64 |
| Poco lluviosa | 7,12 | 1,68 | 20,30 | 20,02 | 21,45 | 27,85 | 4,35 | 0,38 | 0,62 | 42,0 |
| Z1-α * S x̅ | ±0,10 | ±0,06 | ±0,16 | ±0,14 | ±0,06 | ±0,35 | ±0,06 | ±0,02 | ±0,02 | ±0,77 |
Z1-α * S x̅ = ± intervalo de confianza (1-α = 0,05), siendo Z1 = 1,96. Cada valor es promedio de 24 muestras compuestas
Las precipitaciones anuales en los años 2015 y 2016 fueron de 1211 y 1370 mm, ligeramente inferiores en el año 2015 a la media histórica (periodo 1969-2016) de 1348 mm, aunque en los dos años las precipitaciones en el periodo lluvioso (mayo-octubre) alcanzaron el 76 y 78 % del total anual respectivamente, muy similar al porcentaje histórico de 77 %. Las temperaturas medias anuales fueron de 24,6 y 24,24 °C, respectivamente, similares a la media histórica de 24,3 °C, aunque en el periodo lluvioso fueron de 26,03 °C, superiores en 3,8 °C a las del periodo poco lluvioso.
Los experimentos se desarrollaron con los mismos cultivos precedentes y épocas referidos en una anterior publicación 20. En la época lluviosa (Tabla 2) fueron maíz (Zea mayz L./var. MC-4) y frijol vigna (Vigna unguiculata (L.) Walp./var. Guariba); en el caso de la siembra de enero se utilizaron la misma variedad de maíz y el frijol común (Phaseolus vulgaris/var. BAT-304). A continuación de la cosecha de los cultivos precedentes y en tres momentos diferentes (30, 45 y 60 días), se plantó el cultivo del boniato (Ipomea batatas (L) Lam/var.'CEMSA 78-354’).
Como los cultivos precedentes fueron diferentes en las épocas y además no poseen la misma extensión de su ciclo biológico, los experimentos no incluyeron los cultivos, ni la época como factores y se desarrollaron como experimentos independientes. Para cada cultivo y en cada época se ejecutaron tres experimentos, cada uno con la misma fecha de siembra del precedente y variando los periodos entre la cosecha de este y la plantación del boniato (Tabla 2) para un total de 12 experimentos. Los tratamientos (Tabla 3) y el diseño utilizado en cada experimento fueron similares: bloques al azar de cuatro tratamientos, que incluían tanto la etapa del cultivo precedente como la del boniato (factor A) y repetidos durante dos años (factor B), con arreglo factorial de 4 x 2 y con cuatro réplicas.
Los marcos de plantación utilizados fueron similares a los reportados en una publicación anterior 20, 0,9 x 0,3 m para el maíz, 0,7 x 0,2 m para el frijol común y 0,9 x 0,1 m para la vigna. En el cultivo del boniato el marco de plantación fue 0,9 x 0,23 m para las sucesiones comenzadas en el periodo lluvioso y 0,9 x 0,3 m para las otras.
Las parcelas estuvieron constituidas por cinco surcos de 6 metros de largo, excepto para el frijol, las cuales tenían seis surcos. En cada experimento, los cultivos precedentes y el boniato de cada tratamiento se ubicaron siempre en las mismas parcelas.
Tabla 2 Fechas de siembra y cosecha de los cultivos en las diferentes sucesiones en ambos años
| Época1 | Cultivo precedente | Siembra | Cosecha | Plantación del boniato | Cosecha | ||
|---|---|---|---|---|---|---|---|
| 30 días | 45 días | 60 días | |||||
| Lluviosa | Maíz año 1 | 11/5/2015 | 12/9/2015 | 15/10/15 | 30/10/15 | 16/11/15 | 4 meses |
| Vigna año 1 | 10/6/2015 | 12/9/2015 | 15/10/15 | 30/10/15 | 16/11/15 | 4 meses | |
| Maíz año 2 | 11/4/2016 | 12/8/2016 | 15/9/16 | 30/9/16 | 15/10/16 | 4 meses | |
| Vigna año 2 | 10/5/2016 | 12/8/2016 | 15/9/16 | 30/9/16 | 15/10/16 | 4 meses | |
| Poco lluviosa | Maíz año 1 | 15/1/2015 | 16/5/2015 | 18/6/15 | 3/7/15 | 18/7/15 | 4 meses |
| Fríjol año 1 | 9/2/2015 | 16/5/2015 | 18/6/15 | 3/7/15 | 18/7/15 | 4 meses | |
| Maíz año 2 | 16/1/2016 | 18/5/2016 | 18/6/16 | 4/7/16 | 19/7/16 | 4 meses | |
| Fríjol año 2 | 10/2/2016 | 18/5/2016 | 18/6/16 | 4/7/16 | 19/7/16 | 4 meses | |
1La época se define por el comienzo de la sucesión
Tabla 3 Tratamientos estudiados en cada sucesión y para cada uno de los periodos entre cosecha del precedente y plantación del boniato
| Tratamientos | Etapa cultivos precedentes | Etapa cultivo del boniato |
|---|---|---|
| 1 | 50 % NPK+HMA | 50 % NP 25 % K + HMA |
| 2 | 50 % NPK+HMA | 50 % NP 25 % K |
| 3 | 50 % NPK | 50 % NP 25 % K |
| 4 | 100 % NPK | 100 % NPK |
Precedentes en época lluviosa: maíz (var. MC-4) y vigna (var. Guariba). Precedentes en época poco lluviosa: maíz (var. MC-4) y frijol común (var. BAT-304). Variedad de boniato 'CEMSA 78-354’. HMA: R. irregulare/INCAM11.
Inoculante e inoculación micorrízica
El inoculante se preparó en el Instituto Nacional de Ciencias Agrícolas (INCA), Mayabeque, Cuba, a base de Rhizophagus irregulare23/INCAM11 y utilizando Urochloa decumbens (Hochst. ex A. Rich.) como planta hospedera. Esta cepa se recomienda como cepa eficiente para esta condición edáfica 10. El inoculante poseía 30 esporas g-1. La inoculación se ejecutó vía recubrimiento de las semillas, en el caso de los granos con una cantidad de inoculante equivalente al 8 % del peso de las mismas 24, con dosis de 2 y 4 kg ha-1 para el maíz y el frijol, respectivamente y en el boniato se recubrió el tercio inferior de los esquejes, con una mezcla de 1 kg de inoculante en 5 litros de agua y aplicando 35 kg ha-1 (14.
Fertilización y atenciones culturales
En el caso de la fertilización para altos rendimientos (100 % NPK) se corresponden con las de los respectivos Instructivos 25-28 y fueron: 90, 130, 170 kg ha-1 de N, P2O5 y K2O, para el maíz, de 80, 60, 90 kg ha-1 para el frijol y la vigna, así como de 120, 100, 300 kg ha-1 de N, P2O5 y K2O, para el boniato. Las dosis aplicadas para obtener un funcionamiento micorrízico óptimo en los diferentes precedentes inoculados fueron del 50 % NPK 24 y en el caso del boniato se aplicaron 50 % NP 25 % K 29-31. Las labores a los cultivos precedentes y al boniato, incluyendo el riego, se realizaron según los Instructivos Técnicos de estos cultivos 25-28. Es de señalar que cuando los cultivos precedentes se cosecharon, el suelo se preparó de forma convencional, con una secuencia de labores de roturación (arado), dos pases de grada y finalmente se surcó, ajustando los tiempos entre labores a los tres intervalos estudiados.
Evaluaciones
Análisis de suelo
Se tomaron muestras de suelos compuestas en la profundidad de 0-20 cm en cada una de las réplicas de los experimentos y al inicio de estos. Las determinaciones realizadas fueron las establecidas en el laboratorio de suelos y tejido vegetal del Instituto Nacional de Ciencias Agrícolas 32 y consistieron en : pH en KCl y H2O, con una relación suelo-solución de 1:2,5; nitrógeno total (Nt) por el método de micro-Kjeldahl; materia orgánica según Walkley-Black; el P2O5 y K2O con extracción con solución de (NH4)2CO3 con pH 9 y los cationes intercambiables (Ca, Mg, Na y K) por extracción con AcNH4 1N a pH 7,0.
Colonización total de las raíces con HMA (%)
Se efectuó a los 45 días después de la siembra de maíz, vigna y frijol común y a los 90 días después de la plantación del boniato. Para la colonización total se tomaron muestras de raíces finas de ocho plantas por parcela, ubicadas en los surcos centrales. Las raíces fueron teñidas 33 y evaluadas por el método de los interceptos 34.
Conteo de esporas micorrízicas
El número de esporas micorrízicas fue determinado al inicio y en el momento de la cosecha de cada cultivo y se expresó como número total de esporas en 50 g de suelo. Se tomó una muestra compuesta de 10 submuestras (0-20 cm) por parcela y la extracción de esporas se realizó por el método del decantado húmedo 35 y el conteo en microscopio estéreo.
Rendimiento
Para su estimación en t ha-1, fueron cosechadas las plantas de cada parcela, excluyendo las que se encontraban en los dos surcos de borde. En los cultivos del maíz, vigna y frijol se realizó a los 120, 90 y 100 días, respectivamente y se expresó en base a 14 % de humedad. En el boniato se realizó a los 120 días y solo se cuantificaron las raíces comerciales (>100 g).
El efecto de permanencia (EP) en cada intervalo se estimó a partir de las respuestas en el rendimiento del boniato de los tratamientos precedente inoculado-boniato (R1) y precedente inoculado-boniato inoculado (R2), en relación al rendimiento del boniato en la sucesión homóloga no inoculada y se calculó como sigue
Análisis estadístico
Se realizaron los ANOVA de acuerdo con el diseño utilizado. Para los cultivos precedentes, al disponer de información de tres experimentos para cada combinación precedente por épocas, se procedió primeramente a evaluar si existía interacción tratamientos por repetición de experimentos por años y los resultados se presentan a partir de ese análisis. Para el cultivo del boniato los resultados se expresan de acuerdo a si la interacción tratamientos por años en cada experimento fue o no significativa. Además, en el cultivo del boniato se establecieron análisis de regresión entre el porcentaje de colonización vs el rendimiento y vs las cantidades de esporas en 50 g para cada sucesión y época, utilizando los datos correspondientes a los tres intervalos de plantación. La comparación entre experimentos se realizó con los intervalos de confianza (p<0,05), calculados a partir de los Esx obtenidos en los ANOVA realizados para cada variable en los diferentes experimentos.
RESULTADOS
En ninguno de los experimentos los términos de interacción fueron significativos, por lo cual solo se presentarán los efectos del factor tratamientos sobre cada una de las variables en los diferentes experimentos.
Cultivos precedentes en época lluviosa y cultivo principal en época poco lluviosa
Tanto el maíz como la vigna presentaron respuestas significativas (p<0,05) a las dosis de fertilización, como a la inoculación con la cepa eficiente de HMA, de forma tal que no existieron diferencias entre los rendimientos de los tratamientos inoculados que recibieron dosis medias de fertilización y los que recibieron las dosis superiores de fertilización en cada cultivo (Tabla 4A y B). La inoculación micorrízica también originó incrementos significativos del porcentaje de colonización micorrízica y de las cantidades de esporas, con relación a los tratamientos no inoculados, mostrando los tratamientos no inoculados que recibieron las mayores dosis de fertilización, los menores valores; no obstante, los valores de colonización y de esporas obtenidos en el precedente maíz inoculado fueron superiores (p<0,05) que cuando se utilizó vigna como tal.
Tabla 4 Efectos de la inoculación con HMA en los precedentes y efectividad del efecto de permanencia en las sucesiones iniciadas en la época lluviosa
| Maíz época lluviosa | Boniato época poco lluviosa | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 30 días | 45 días | 60 días | |||||||||||
| Tratamientos | t ha-1 | Col | # esp | Tratamientos | t ha-1 | Col | # esp | t ha-1 | Col | # esp | t ha-1 | Col | # esp |
| HMA 50 %NPK | 3,59a | 72,8a | 735a | HMA 50 %NPK | 25,75a | 72,3a | 715a | 25,1a | 71,8a | 713a | 25,9a | 73,5a | 715a |
| HMA 50 %NPK | 3,58a | 71,4a | 736a | 50 %NPK | 25,49a | 70,8a | 712a | 24,1b | 70,1a | 712a | 20,4b | 37,1b | 310b |
| 50 % NPK | 3,36b | 10,9b | 71b | 50 % NPK | 21,33b | 10,1b | 69b | 20,9c | 9,9b | 69b | 20,6b | 8,5c | 70c |
| 100 % NPK | 3,58a | 9,5b | 54c | 100 % NPK | 25,84a | 9,8b | 61c | 25,5a | 8,9b | 60c | 25,8a | 8,5c | 60d |
| EsX | 0,01 | 0,53 | 0,99 | EsX | 0,235 | 0,38 | 1,1 | 0,20 | 0,49 | 1,1 | 0,21 | 0,39 | 1,3 |
| p | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | |
| Vigna época lluviosa | Boniato época poco lluviosa | ||||||||||||
| HMA 50 %NPK | 2,47a | 58,8a | 521a | HMA 50 %NPK | 25,8a | 54,3a | 517a | 26,1a | 53,9a | 516a | 26,1a | 55,3a | 516a |
| HMA 50 %NPK | 2,48a | 57,8a | 520a | 50 %NPK | 25,9a | 54,5a | 516a | 26,1a | 52,9a | 513a | 21,1b | 41,8b | 226b |
| 50 % NPK | 2,26b | 9,3b | 56b | 50 %NPK | 20,9b | 10,4b | 56b | 21,2b | 9,8 | 56b | 21,2b | 10 c | 56 c |
| 100 % NPK | 2,49a | 8,3b | 51c | 100 % NPK | 26,1a | 8,6c | 41c | 26,3a | 8,4 | 37c | 26,3a | 8,3d | 41d |
| EsX | 0,03 | 0,27 | 0,44 | EsX | 0,14 | 0,31 | 1,0 | 0,18 | 0,33 | 1,5 | 0,21 | 0,43 | 1,5 |
| p | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | |
col (% colonización micorrízica); #esp (esporas 50 g-1). HMA: R. irregulare /INCAM 11. Las interacciones con las repeticiones y años no fueron significativas. Para cada cultivo precedente o intervalo de plantación del boniato, letras diferentes en cada columna conllevan a diferencias significativas a p< 0,05 según Prueba de Duncan. p: valor p del factor tratamientos en los diferentes ANOVA realizados.
En relación con el boniato, este cultivo siempre respondió significativamente a la inoculación y a la mayor fertilización, de forma tal que ambos tratamientos alcanzaron rendimientos similares en los diferentes experimentos con uno y otro precedente (Tabla 4A y B). En los tratamientos en que se inoculó el precedente y no se inoculó el boniato, el efecto de permanencia se mantuvo en los primeros 30 días, para ambos precedentes, con rendimientos similares con los tratamientos en que el boniato también se inoculó, pero ya a los 45 días, solo fue completamente efectivo en la sucesión vigna inoculada-boniato. Si bien con el precedente de maíz inoculado el rendimiento alcanzado fue superior al tratamiento homólogo sin inoculación, el efecto fue del 76 % comparado con el incremento en el rendimiento obtenido por el tratamiento en que ambos cultivos en la sucesión se inocularon. A los 60 días, el efecto desapareció completamente con ambos precedentes.
Tanto los porcentajes de colonización como las esporas, respondieron significativamente a la inoculación del boniato, con valores superiores (p<0,05) cuando el maíz fue precedente. Para cada precedente y en los tratamientos con precedentes inoculados y boniato sin inocular se presentaron durante los intervalos de 30 y 45 días, valores similares para ambos indicadores micorrízicos, en relación a los tratamientos en que los dos cultivos en la sucesión se inocularon y ambos indicadores disminuyeron bruscamente (p<0,05) en el intervalo de 60 días, aunque con valores superiores a los tratamientos homólogos no inoculados. Los tratamientos no inoculados y que recibieron las mayores dosis de fertilización, exhibieron siempre los menores valores de los indicadores del funcionamiento micorrízico evaluados.
En las dos sucesiones los rendimientos del boniato se asociaron fuertemente con los porcentajes de colonización micorrízica (Figura 1). En la sucesión maíz-boniato los mayores rendimientos del boniato se alcanzaron con porcentajes de colonización en el rango de 70 a 75 %; mientras que al utilizar la vigna como cultivo precedente, se lograron rendimientos similares con porcentajes de colonización inferiores a los obtenidos en la anterior sucesión (p<0,05) y del orden de 52 a 56 %.
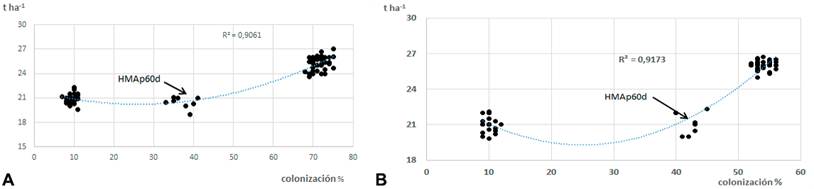
Figura 1 Relaciones en el cultivo del boniato entre el rendimiento y los porcentajes de colonización para las sucesiones maíz-boniato (A) y vigna-boniato (B)
Se utilizaron los datos correspondientes a los tres intervalos, los dos años y cuatro réplicas de los tres tratamientos que recibieron la misma dosis de fertilizantes con y sin inoculación (n=72). HMAp60d (precedente inoculado-boniato sin inocular, intervalo 60 días).
Es de señalar que en los gráficos de rendimiento vs colonización y para ambas sucesiones (Figura 1A y 1B), se puede observar cómo valores del orden de 40 % de colonización, alcanzados en el intervalo de 60 días, presentaron el mismo rendimiento que los tratamientos no inoculados con cerca de 10 % de colonización. Asimismo, en ambas sucesiones, la reproducción de esporas por el boniato se asoció significativamente con los porcentajes de colonización alcanzado por este cultivo (R2 = 0,99), con valores siempre crecientes para ambas variables, aunque la producción de esporas, cuando la vigna fue precedente, fue mucho menor que con el maíz (Tabla 4).
Cultivos precedentes en época poco lluviosa y cultivo principal en época lluviosa
Con ambos precedentes se encontró una respuesta significativa en el rendimiento a la inoculación y a la fertilización (Tabla 5A y B), de forma tal que los tratamientos inoculados con la cepa eficiente HMA y dosis media de fertilización alcanzaron rendimientos similares a los obtenidos con las mayores dosis de fertilización. Asimismo, tanto el maíz como el frijol inoculados presentaron valores muy superiores (p<0,05) de porcentajes de colonización y de cantidades de esporas que los no inoculados, aunque ambos fueron significativamente mayores en maíz que en frijol (p<0,05). A su vez, las mayores dosis de fertilización presentaron los menores valores de estos dos indicadores del funcionamiento micorrízico.
Además, en la época lluviosa el maíz alcanzó no solo mayores rendimientos que cuando fue sembrado en la poco lluviosa, sino también mayores porcentajes de colonización micorrízica y de producción de esporas. Asimismo, la vigna alcanzó valores superiores (p<0,05) de los indicadores de funcionamiento micorrízico evaluados que el frijol.
El boniato asimismo, presentó una respuesta positiva a la inoculación y a la fertilización mineral (Tabla 5A y B) con rendimientos similares entre ambos en los diferentes experimentos, aunque mayores (p<0,05) cuando el cultivo precedente fue frijol. De forma similar a la anterior época, el efecto de permanencia se alcanzó con ambos precedentes en el intervalo de 30 días; a los 45 solo se mantuvo completamente con el precedente de frijol inoculado, aunque con el maíz fue del orden del 81 % y desapareció totalmente en ambas sucesiones a los 60 días.
Tanto los porcentajes de colonización como las esporas, respondieron significativamente a la inoculación del boniato, con valores superiores (p<0,05) cuando el maíz fue precedente. Para cada precedente y en los tratamientos con precedentes inoculados y boniato sin inocular y con los intervalos de 30 y 45 días, se presentaron valores similares de ambos indicadores micorrízicos, en relación a los tratamientos en que los dos cultivos en sucesión se inocularon. Sin embargo, ambos indicadores disminuyeron bruscamente (p<0,05) en el intervalo de 60 días, aunque con valores superiores a los tratamientos homólogos no inoculados. En los tratamientos no inoculados, las mayores dosis de fertilización redujeron (p<0,05) los valores de los indicadores del funcionamiento micorrízico evaluados.
Asimismo, en las dos sucesiones, el rendimiento del boniato se asoció fuertemente con los porcentajes de colonización micorrízica, obtenidos en este cultivo (Figura 2). Cuando el precedente fue maíz los mayores rendimientos del boniato se alcanzaron con porcentajes de colonización en el rango de 59 a 64 %; mientras que en la sucesión frijol-boniato los porcentajes de colonización que se asociaron con los mayores rendimientos fueron inferiores a los alcanzados con el maíz (p<0,05) y estuvieron en el rango de 51 a 57 %; no obstante, los rendimientos del boniato fueron superiores en esta última sucesión. En ambas sucesiones, las reproducciones de esporas realizadas por el boniato se asociaron fuertemente con los porcentajes de colonización alcanzados por este cultivo (R2 = 0,98), con valores siempre crecientes para ambas variables, aunque la esporulación con el frijol precedente fue mucho menor que con el maíz (Tabla 5).
Tabla 5 Efecto de la inoculación con HMA en los precedentes y efectividad del efecto de permanencia en las sucesiones iniciadas en la época poco lluviosa
| Maíz época poco lluviosa | Boniato época lluviosa | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 30 días | 45 días | 60 días | |||||||||||
| Tratamientos | t ha-1 | col | # esp | Tratamientos | t ha-1 | col | # esp | t ha-1 | col | # esp | t ha-1 | col | # esp |
| HMA 50 %NPK | 3,14a | 60,6a | 562a | HMA 50 %NPK | 29,7a | 62,5a | 553a | 29,4a | 61,75a | 552a | 29,8a | 61,5a | 552a |
| HMA 50 %NPK | 3,13a | 59,25a | 562a | 50 %NPK | 29,3a | 61,3a | 551a | 28,4a | 60,6a | 549a | 25,5b | 36,5b | 229b |
| 50 % NPK | 2,61b | 10,5b | 62b | 50 % NPK | 24,9b | 10b | 61b | 24,1b | 9,5b | 60,8b | 24,2b | 10c | 61c |
| 100 % NPK | 3,15a | 9c | 58c | 100 % NPK | 30,2a | 8,6b | 57c | 29,3a | 8,5b | 55,6c | 29,3a | 9,8c | 55d |
| EsX | 0,03 | 0,36 | 0,5 | EsX | 0,14 | 0,41 | 0,7 | 0,14 | 0,36 | 0,9 | 0,21 | 0,4 | 0,8 |
| p | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | |
| Frijol época poco lluviosa | Boniato época lluviosa | ||||||||||||
| HMA 50 %NPK | 2,42a | 51,5a | 449a | HMA 50 %NPK | 31,5a | 54,0a | 438a | 31,1a | 53,8a | 437a | 31,03a | 55,5a | 437a |
| HMA50 %NPK | 2,41a | 50,8a | 448a | 50 %NPK | 31,2a | 53,5a | 437a | 30,7a | 53,1a | 436a | 25,08b | 35,5b | 139b |
| 50 % NPK | 2,06b | 9,1b | 49b | 50 % NPK | 25,4b | 11,1b | 47 b | 25,4b | 10,1b | 47 b | 25,11b | 9,1c | 46c |
| 100 % NPK | 2,42a | 8,4b | 44c | 100 % NPK | 31,4a | 9,5c | 40c | 31,2a | 8,6 | 39c | 31,38a | 8,6c | 38d |
| EsX | 0.02 | 0,38 | 0,70 | EsX | 0,235 | 0,40 | 0,81 | 0,271 | 0,42 | 1,0 | 0,21 | 0,36 | 1,0 |
| p | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | 0,000 | |
col (% colonización micorrízica); #esp (esporas 50 g-1). HMA: R. irregulare /INCAM 11. Las interacciones con las repeticiones y años no fueron significativas. Para cada cultivo precedente o intervalo de plantación del boniato, letras diferentes en cada columna conllevan a diferencias significativas a p< 0,05 según Prueba de Duncan. p:valor p del factor tratamientos en los diferentes ANOVA realizados.
De forma similar a la otra época, los valores del orden de 40 % de colonización, alcanzados con ambos precedentes en el intervalo de 60 días, resultaron en el mismo rendimiento que los tratamientos no inoculados con porcentajes de colonización cercanos al 10 % (Figura 2A y 2C). Hubo diferencias significativas entre épocas (p<0,05) para los porcentajes de colonización y cantidad de esporas en el boniato, con los mayores valores cuando los precedentes se sembraron en la época lluviosa; no obstante, el rendimiento del boniato fue superior al plantar el mismo en la época lluviosa; es decir, con los precedentes sembrados en la poco lluviosa.

Figura 2 Relaciones entre el rendimiento y el porcentaje de colonización micorrízica en el cultivo del boniato, para las sucesiones maíz-boniato (A) y para la sucesión vigna-boniato (B)
Se utilizaron los datos correspondientes a los tres intervalos, los dos años y cuatro réplicas de los tres tratamientos que recibieron la misma dosis de fertilizantes (n=72 en cada caso). HMA p60d (precedente inoculado-boniato sin inocular en intervalo 60 días).
DISCUSIÓN
La respuesta a la inoculación con una cepa eficiente de HMA de los cultivos precedentes en ambas épocas, expresada de que con solo el 50 % de la fertilización se garantizan altos rendimientos, superiores al homólogo no inoculado (p<0,05) y similares a los obtenidos con altas dosis de fertilizantes (100 % NPK) corrobora los resultados obtenidos anteriormente en diferentes cultivos y condiciones edáficas 8,24 y deja claro uno de los beneficios de la inoculación.
Asimismo, los cultivos precedentes inoculados alcanzan altos porcentajes de colonización propios de un funcionamiento micorrízico efectivo 8. Así, en maíz, los registros son superiores a los de las leguminosas y, a su vez, en la época lluviosa de siembra los porcentajes son mayores que los obtenidos en la poco lluviosa y corroboran lo obtenido con anterioridad 20. Esto último posiblemente esté asociado con los mayores crecimientos y rendimientos obtenidos en la época lluviosa, que puede conllevar a un incremento de las necesidades de estructuras fúngicas que satisfagan los requerimientos de las plantas, tal como ocurre con diferentes especies de forrajeras inoculadas cuando se cultivan en una y otra época 36.
Los mayores conteos de esporas obtenidos en las siembras de los precedentes en la época lluviosa pueden ser explicados por los mayores porcentajes de colonización y un mayor crecimiento existentes en ambos cultivos inoculados en esta época. De forma similar en el maíz, su sistema radical más profuso de conjunto con las mayores colonizaciones alcanzadas, deben explicar la mayor cantidad de esporas asociadas. Además, de que estos resultados fueron similares a los obtenidos anteriormente con estos cultivos 20, diversos autores han reportado en diferentes cultivos inoculados con cepas eficientes de HMA, mayores cantidades de esporas asociadas con los mayores porcentajes de colonización micorrízica y con las mayores producciones de biomasa o rendimiento 29,30,36,37, así como para un mismo cultivo, mayores en periodo lluvioso que en poco lluvioso 36. En correspondencia con lo anterior otros autores 38 plantean que un funcionamiento micorrízico eficiente puede conllevar a que las ganancias de C obtenidas por el hongo se inviertan en la producción de esporas, mecanismo que no solo es favorecido en las especies de carácter generalista que se caracterizan por una alta producción de esporas 39, sino que se presenta para un grupo amplio de géneros y especies de Glomeromicota 40.
La información corroboró el efecto de permanencia encontrado en los primeros 30 días para los diferentes precedentes en ambas épocas 20, pero solo se hace completamente extensivo hasta los 45 días con los precedentes de leguminosas, pese a que en ese caso, se producen menos esporas que cuando el precedente fue el maíz. No obstante, con el maíz inoculado se alcanza a los 45 días un rendimiento equivalente a cerca del 80 % del efecto, lo cual tiene una notable importancia práctica. Es de destacar que el efecto se logra aun cuando se ejecutaron las labores comunes de preparación del suelo, previo a la plantación del boniato.
Se reconoce que las labores al suelo afectan la estabilidad de la micorrización residente 41, aunque en los diversos trabajos realizados en Cuba sobre manejo de la inoculación en secuencias de cultivos 8,14,20 este efecto se ha reportado en presencia de las labores del suelo e incluso en un trabajo en que se compararon laboreo mínimo y convencional, se obtuvieron efectos de permanencia similares entre ambos 42 y posiblemente relacionado con el hecho de que las cepas generalistas como R. irregulari se caracterizan por la alta producción de esporas 39, propágulos más resistentes al laboreo que el micelio extraradical.
En diferentes trabajos realizados con cultivos micótrofos e inoculando una cepa eficiente 11,14,17, el efecto desaparece en algún momento, dependiente su extensión de la interacción con las poblaciones de hongos micorrízicos residentes y la propia competitividad de la cepa inoculada 18, aunque también influyen la condición edáfica, el tipo de cultivo y su manejo 11,19. La desaparición del efecto parece ser importante, ya que disminuye posibles riegos ecológicos asociados a la inoculación de cepas no residentes 18, aunque en el caso específico de R. irregulare se ha reportado muy bajo endemismo 43 y dichos autores consideran que el riesgo de introducción de una cepa “exótica” de esta especie parece menor que con otras especies.
Un aspecto interesante es que las magnitudes de la colonización y las cantidades de esporas reproducidas por los precedentes inoculados parecen mantener una influencia directa en los porcentajes de colonización y la cantidad de esporas obtenidas en el boniato en sucesión. Muy probablemente la mayor cantidad de esporas micorrízicas existentes al plantar el boniato y derivadas de la inoculación del cultivo precedente, originan mayores colonizaciones del mismo, tal y como ha sido encontrado en experimentos anteriores con estos mismos cultivos 20 o por otro autor con variedades de forrajes inoculadas en la siembra y sometidas a cortes periódicos 44, el cual fundamenta con la cantidad de esporas presentes al realizar un corte, la permanencia o no de una micorrización efectiva para la nueva etapa de crecimiento.
Asimismo la mayor colonización del boniato, debe originar altas cantidades de esporas, lo cual es un resultado similar al encontrado en los precedentes inoculados y ha sido encontrado en este mismo cultivo 29-31 y, además, en diversos experimentos de comparación de cepas de HMA, en que la inoculación de la cepa eficiente no solo origina mayores porcentajes de colonización y rendimiento, sino también mayores cantidades de esporas 45-47; sin embargo, no explica porque se mantienen cantidades similares de esporas a lo largo de cada sucesión en los tratamientos inoculados.
Además, aún los cultivos inoculados no se colonizan solo con la cepa aplicada y los diferentes cultivos poseen diferentes capacidades, tanto para colonizarse con una u otra cepa 6,38,40, por lo cual, aunque los porcentajes de colonización son un indicador del funcionamiento y se relacionan, de forma general, positivamente con el rendimiento 48,49, el funcionamiento micorrízico es un fenómeno muy complejo y no se pueden inferir mejores funcionamientos al comparar cultivos inoculados, solo por los valores de estos porcentajes.
De forma similar en un experimento con 17 especies de crotalaria 50 para evaluar la respuesta a la inoculación micorrízica y el efecto de permanencia sobre un cultivo de cebolla (Allium cepa L.) en sucesión, los mayores porcentajes de colonización en la cebolla no se asociaron con las especies de crotalaria que presentaron los mayores porcentajes de colonización.
Por otro lado, aunque algunos autores han relacionado positivamente la existencia del efecto de permanencia con la cantidad de esporas asociadas al cultivo inoculado; por ejemplo, en la sucesión Canavalia ensiformis inoculada-banano 12 y en áreas de forraje de varias especies de braquiarias inoculadas 44, las diferentes características en la reproducción de esporas por los diversos cultivos 51, las diferencias en la comunidad micorrízica residente, la influencia de las épocas y el propio hecho de que las esporas no son el único propágulo infectivo 6,38, si bien pueden permitir hacer valoraciones para cada cultivo inoculado en un agrosistema en particular, no permiten comparar la potencialidad de los diferentes cultivos precedentes inoculados para obtener el efecto de permanencia, a partir de las esporas totales reproducidas por cada uno de ellos.
La información obtenida confirma la existencia de un efecto de permanencia del inoculante aplicado con una alta importancia para su uso en las tecnologías de los cultivos y algunos autores han establecido, utilizando técnicas moleculares, una confirmación positiva de las cepas aplicadas en los cultivos inoculados hasta un periodo de dos años posterior a la inoculación 18; sin embargo, otros plantean que el efecto positivo de la inoculación no se vincula necesariamente a un efecto directo de la cepa inoculada 52 y, por tanto, es imprescindible desarrollar los protocolos para el seguimiento de la cepa INCAM11, mediante marcadores moleculares específicos, para establecer el mecanismo presente en el efecto de permanencia encontrado.
La utilización del frijol inoculado como precedente parece presentar otro beneficio adicional, asociado a los mayores rendimientos encontrados en el boniato, en comparación a los obtenidos en la sucesión maíz-boniato, lo cual puede estar relacionado, entre otros, con la influencia positiva de los residuos de las leguminosas con mayor facilidad para mineralizarse que los residuos de maíz con mayor relación C/N y, además, la importancia del nitrógeno para el boniato 29.
Resultó interesante que los valores intermedios de colonización alcanzados en el boniato en el intervalo de 60 días por las diferentes sucesiones con precedentes inoculados y boniato sin inocular no originaran rendimientos, aunque fuera ligeramente superiores a los obtenidos en los tratamientos homólogos sin inocular, indicando que esta cantidad de estructuras fúngicas es insuficiente para garantizar una respuesta positiva en el rendimiento 53. El porcentaje de colonización, tal como se evaluó, es inespecífico y en esta situación de desaparición del efecto de permanencia, es lógico considerar que también ha disminuido la proporción en que la cepa eficiente inoculada está presente en la colonización del boniato, lo cual enmascara posibles valoraciones sobre eficiencia de la cepa inoculada. No obstante, resultados similares han sido reportados en un experimento de manejo de la inoculación con esta misma cepa en secuencia de cultivos y en el mismo tipo de suelo 14, en que el boniato era el tercer cultivo en sucesión y en el cual los porcentajes de colonización alcanzados de 38 % no originaban rendimientos superiores a los controles no inoculados, los cuales presentaban valores muy inferiores de colonización.
Sin embargo, en cultivos perennes inoculados se ha reportado una desaparición gradual del funcionamiento micorrízico con decrecimientos proporcionales entre las colonizaciones y los rendimientos asociados 17,44. La aparente contradicción puede estar relacionada, entre otros, con características diferentes del funcionamiento micorrízico en los cultivos 38, como con el hecho de que quizás no sea lo mismo establecer una colonización efectiva, que debe requerir de un número mínimo de propágulos de cepas eficientes 53, que ir disminuyendo la efectividad de un funcionamiento existente en un cultivo perenne, en el cual sin duda, está presente un efecto acumulado sobre el crecimiento, que puede enmascarar las relaciones entre el porcentaje de colonización actual y la respuesta de la planta 54.
En los diferentes cultivos las dosis superiores de fertilizantes presentaron los menores valores de los indicadores de funcionamiento, corroborando los aspectos más generales ya reportados de que las cantidades de fertilizantes diseñadas para altos rendimientos, disminuyen o inhiben el funcionamiento micorrízico, tanto en cultivos inoculados 8, como el proveniente de las comunidades micorrízicas residentes 6,38.
CONCLUSIONES
La inoculación con cepas eficientes de HMA de los cultivos precedentes, resulta una vía adecuada para lograr un funcionamiento micorrízico efectivo en el boniato en sucesión en las dos épocas de plantación de este cultivo.
La utilización de la vigna y frijol común como cultivos precedentes, permite aprovechar este efecto de permanencia de la inoculación, hasta 45 días después de la cosecha de estos cultivos y superior a los resultados obtenidos con el maíz.
La potencialidad de los precedentes inoculados para desarrollar el efecto de permanencia no puede ser establecida a través de la simple comparación de los indicadores de funcionamiento obtenidos en cada uno de estos.

