










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Medicina Tropical
versión On-line ISSN 1561-3054
Rev Cubana Med Trop v.50 n.2 Ciudad de la Habana Mayo-ago. 1998
Instituto de Medicina Tropical "Pedro Kourí"
Selección de una cepa de Culex quinquefasciatus resistente a lambdacialotrina y su espectro de resistencia cruzada a otros insecticidas
Lic. María Magdalena Rodríguez,1 Dr. Juan Bisset,2 Lic. Cristina Díaz1 y Téc. Alain Soca3
Resumen
Se seleccionó una cepa de campo de Culex quinquefasciatus resistente al insecticida piretroide lambdacialotrina, con el objetivo de poder utilizarla como cepa de referencia en el laboratorio, en los estudios de bioquímica y genética de la resistencia; evaluar la utilidad de este insectivida para el control de mosquitos en Cuba y determinar la resistencia cruzada a insectividas organofosforados, piretroides y un carbamato. Se obtuvo una alta resistencia a lambdacialotrina después de 6 generaciones de presión de selección. Se observó baja o ninguna resistencia cruzada a otros piretroides (deltametrina y cipermetrina), a un carbamato (propoxur) y a los insecticidas organofosforados (clorpirifos y metil-pirimifos); sin embargo, se observó una alta resistencia cruzada a malatión (organofosforado).
Descriptores DeCS: RESISTENCIA A INSECTICIDA; CONTROL DE MOSQUITOS, CULEX; LARVA; CUBA.
Los insecticidas organofosforados (OP) han sido usados de forma amplia contra los miembros del complejo Culex pipiens (principalmente C.p. pipiens y C.p. quinquefasciatus) en muchos países y los genes para la resistencia han sido identificados a menudo.1-5
En Cuba, se usó malatión (un OP) contra C.p. quinquefasciatus durante 7 años hasta 1986. Éste fue reemplazado por cipermetrina en 1987, debido a la aparición de resistencia, aunque se continuó usando malatión en forma alternada en algunas áreas rurales alrededor de La Habana.3,6,7 La resistencia a malatión fue debida, al menos, a 2 mecanismos de resistencia: una alta actividad de esterasas B y una acetil-colinesterasa insensitiva.3 Estos mecanismos, en apariencia, no son capaces de ofrecer resistencia cruzada a piretroides.6 La resistencia a cipermetrina apareció en el centro de La Habana en 1990,7 y los mecanismos fisiológicos responsables de esta resistencia no son conocidos aún.
Antes de usar un insecticida alternativo se debe investigar la resistencia cruzada que generan los mecanismos de resistencia ya existentes, como se realizó para metil-pirimifos.6
En este trabajo se reporta la selección de una cepa con un insecticida piretroide, lambdacialotrina, la alta resistencia lograda en pocas generaciones y la baja resistencia cruzada observada a insecticidas OP, carbamatos y piretroides.
Métodos
Mosquitos. Se utilizaron 2 cepas: S-LAB, una cepa susceptible de referencia aislada por Georghiou en 1966,8 y SANTIAGO, una población natural, colectada en Santiago de Cuba en mayo de 1995. Esta población fue seleccionada durante 6 generaciones exponiendo las larvas de cuarto estadio a dosis de lambdacialotrina que causaron el 90??5 % de mortalidad (cada generación se designó como SAN Fx donde x representa el número de la generación).
Bioensayos con insecticidas. La resistencia a los insecticidas en las larvas fue analizada por los bioensayos en SANTIAGO y las sucesivas generaciones de selección SAN-F1 a SAN-F6, mediante larvas de cuarto estadio en copas con 99 mL de agua más de 1 mL de solución de insecticida en acetona. Se probaron 7 insecticidas: 3 organofosforados (clorpirifos, malatión, y metil-pirimifos), 3 piretroides (lambdacialotrina, cipermetrina y deltametrina), y 1 carbamato (propoxur). En cada prueba, fueron expuestos grupos de 20 larvas a diferentes dosis de insecticidas durante 24 h. Para cada insecticida se realizaron 5 réplicas correspondientes a 5 dosis diferentes (20 larvas por dosis), que causaron entre 2 y 98 % de mortalidad. Para estandarizar los bioensayos la concentración final de acetona fue ajustada al 1 %. Los datos de mortalidad fueron analizados con el programa Probit-Log,9 basado en el método de Finney.10
Resultados
Evolución de la resistencia en mosquitos de Santiago. La tabla 1 muestra la resistencia a lambdacialotrina en las generaciones sucesivas de selección. Los valores de factores de resistencia (FR50 y FR90) se calcularon dividiendo los valores de CL50 y CL90 obtenidos en cada generación de selección entre los correspondientes valores obtenidos para la cepa de referencia S-LAB. La CL50 de la población natural de SANTIAGO fue de alrededor de 0,0018 p.p.m., alcanzó un valor de 0,0063 p.p.m. después de 3 generaciones de selección y se elevó a un valor de 0,099 p.p.m. para SAN-F6. La última selección con lambdacialotrina incrementó la CL50 a casi 0,1 p.p.m. Esta progresión de los valores de CL50 indica que fueron seleccionados de forma progresiva diferentes genotipos resistentes y que todos los individuos susceptibles fueron rápidamente eliminados. El valor de la pendiente en SAN-F6 (tabla 1) indica que la po-blación aún es heterogénea, por lo que es posible continuar seleccionando.
Resistencia cruzada. El incremento en la resistencia a lambdacialotrina durante 6 generaciones se vio asociado con un incremento en la resistencia a otros piretroides (fig. 1), pero a mucho más bajo nivel, lo cual indica que en SAN-F6, está presente un mecanismo de resistencia específico para lambdacialotrina.
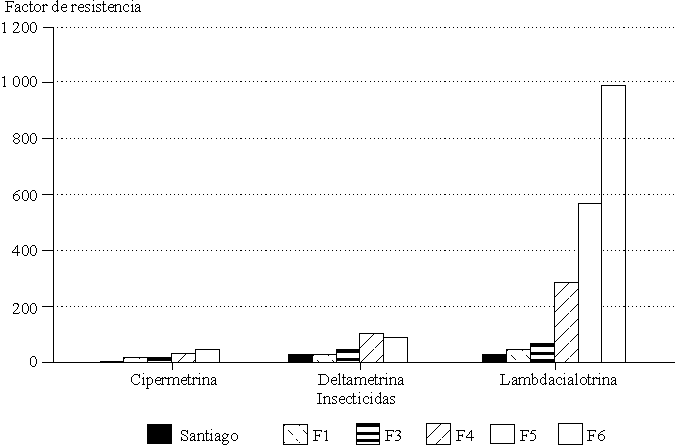
Fig.1. Efecto de la presión de selección con lambdacialotrina durante 6 generaciones (F1-F6) en la variación del factor de resistencia para 3 insecticidas piretroides (cipermetrina, deltametrina y lambdacialotrina) en Culex quinquefasciatus de Santiago de Cuba.
Dentro de los insecticidas organofosforados carbamatos (fig. 2), se observó un incremento significativo de la resistencia a malatión (rango de correlación de Spearman=0,829; p<0,05) asociado con la selección con lambdacialotrina, pero no a metil-pirimifos (rango de correlación de Spearman=0,09; p0,5), clorpirifos (rango de correlación de Spearman=0,03; p0,5) o propoxur (rango de correlación de Spearman= -0,77; p0,5). Esto indica que no se seleccionaron 2 mecanismos de resistencia diferentes, uno ofreciendo resistencia a lambdacialotrina y otro a malatión, por lo que es difícil entender cómo la resistencia diferentes, uno ofreciendo resistencia a malatión, tan específica, puede haber sido seleccionada durante la presión con lmóbdacialotrina.

Fig.2. Efecto de la presión de selección con lambdacialotrina durante 5 generaciones (F1-F5) en la variación del factor de resistencia para 3 insecticidas organofosforados (malatión, metil-pirimifos y clorpirifos) y un carbamato (propoxur) en Culex quinquefasciatus de Santiago de Cuba.
Tabla 1. Variación del factor de resistencia (FR), calculado a partir de los valores CL50 y CL90 en una cepa de Culex quinquefasciatus de Santiago de Cuba y en cada una de las cepas resultantes de la selección con lambdacialotrina (F1,F2,F3,F4,F5 y F6)
| Cepa | CL50 | FR50 | CL90 | FR | b* |
|
| (p.p.m.) |
| (p.p.m.) |
|
|
| Santiago de Cuba | 0,0018 | 18,0 | 0,008 | 40,0 | 1,9 |
| F1 | 0,0036 | 36,0 | 0,0062 | 31,0 | 5,46 |
| F2 | 0,0036 | 36,0 | 0,012 | 60,0 | 2,45 |
| F3 | 0,0063 | 63,0 | 0,013 | 65,0 | 3,76 |
| F4 | 0,032 | 320,0 | 0,062 | 310,0 | 4,45 |
| F5 | 0,058 | 580,0 | 0,16 | 800,0 | 2,86 |
| F6 | 0,09 | 990,0 | 1,45 | 7 250,0 | 1,09 |
| S-LAB | 0,0001 - |
| 0,0002 - |
| 1,6 |
* Pendiente de la curva de mortalidad vs. dosis.
La alta resistencia a lambdacialotrina después de sólo 6 generaciones de selección indica que en corto período, este insecticida no será buen candidato para el control de los mosquitos en Cuba, por lo que es recomendable utilizarlo como alternativo, dentro de una buena política de uso de insecticidas. Esta conclusión está basada en la alta resistencia a malatión existente en Cuba y la resistencia cruzada entre la lambdacialotrina y malatión observada en el presente estudio. Cualquier piretroide usado en esta área debe ser evaluado con cuidado para una posible resistencia cruzada a malatión y metil-pirimifos, la resistencia a malatión está ampliamente diseminada en Cuba y el metil-pirimifos es aún bueno como alternativo para el uso con piretroides y malatión.6
En nuestro estudio no se observó incremento de la resistencia a propoxur con la selección con lambdacialotrina. La sobreproducción de esterasas no puede ser excluida, ya que la frecuencia de este mecanismo de resistencia se incrementó durante el proceso de selección (tabla 2), aunque se requieren trabajos futuros para identificar los mecanismos de resistencia específicos.
Tabla 2. Variación de la frecuencia del gen de esterasa y acetilcolinesterasa durante 6 generaciones (F1, F2, F3, F4, F5 y F6) de selección con el insecticida piretroide lambdacialotrina en Culex quinquefasciatus de Santiago de Cuba
| Cepa | Esterasa | Acetilcolinesterasa |
| Santiago de Cuba | 0,54 | 0,67 |
| F1 | 0,77 | 0,73 |
| F2 | 1,0 | 0,77 |
| F3 | 1,0 | 0,68 |
| F4 | 1,0 | 0,58 |
| F5 | 1,0 | 0,87 |
| F6 | 1,0 | 1,0 |
Discusión
En Cuba se conocen 2 mecanismos de resistencia: una sobreproducción de esterasas B y una acetil-colinesterasa (AchE) insensitiva.3 El último no es responsable de la resistencia observada a lambdacialotrina; se ha demostrado que la AchE insensitiva provee una alta resistencia a propoxur.3
La resistencia cruzada a piretroides y organofosforados debido a una superproducción de esterasas ha sido reportada previamente en aphidos.11 En fecha reciente, a través de estudios con sinergistas, se ha reportado en Culex quinquefasciatus una asociación entre la resistencia a piretroides y la actividad de esterasas.12,13
SUMMARY
A field strain of Culex quinquefasciatus resistant to pyrethroid lambda-cyhalothrin insecticide was selected to be used as a reference strain in the laboratory for conducting studies of biochemistry and genetics of resistance, to evaluate the utility of this insecticide for the control of mosquitoes in Cuba, and to determine the cross resistance to organophosphate insecticides, pyrethroids and a carbamate. A hig resistance to lambdacyhalothrin was obtained after 6 generations of selective pressure. Lowe or no cross resistance to other pyrethroids (deltamethrin and cyper-methrin), to a carbamate (propoxur) and to the organophosphate insecticides (clorpirifos and methyl-pyrimifos) was observed. A high cross resistance to malathion (organophosphate) was detected.
Subject headings: INSECTICIDE RESISTANCE; MOSQUITO CONTROL; CULEX; LARVA; CUBA.
REFERENCIAS BIBLIOGRÁFICAS
- Pasteur N, Iseki A, Georghiou GP. Genetic and biochemical studies of the highly active esterases Aand B associated with organophosphate resistance in mosquitoes of the Culex pipiens complex. Biochem Genet 1981;19:909-19.
- Raymond M, Forunier D, Bride JM, Cuany A, Berge J, Magnim M, et al. Identification of resistance mechanisms in Culex pipiens (Diptera: Culicidae) from southern France: insensitive acetylcholinesterase and detoxifying oxidases. J Econom Entomol 1986;79:1452-8.
- Bisset JA, Rodríguez MM, Díaz C, Ortiz E, Marquetti MC, Hemingway J. The mechanisms of organophosphate and carbamate resistance in Culex quinquefasciatus from Cuba. Bull Entomol Res 1990;80:245-50.
- Poirie M, Raymond M, Pasteur M. Identification of two distinct amplifications of the esterase B locus in Culex pipiens (L.) mosquitoes from Mediterranean countries. Biochem Genet 1992;30:13-26.
- Bourguet D, Capela R, Raymond M. An insensitive acetylcholinesterase in Culex pipiens L. mosquitoes from Portugal. J Econom Entomol 1996;89:1060-6.
- Bissett JA, Rodríguez MM, Hemingway J, Díaz C, Small GJ, Ortiz E. Malathion and pyrethroid resistance in Culex quinquefasciatus from Cuba: efficacy of pirimiphos-methyl in the presence of at least resistance mechanisms. Med Vet Entomol 1991;5:223-8.
- Rodríguez M, Ortiz E, Bisset JA, Hemingway J, Salcedo E. Changes in malathion and pyrethroid resistance after cypermethrin selection of Culex quinquefasciatus field populations of Cuba. Med Vet Entomol 1993;7:117-21.
- Georghiou GP, Metcalf RL, Gidden FE. Carbamate resistance in mosquitoes. Selection of Culex pipiens fatigans Wied for resistance to Baygon. Bull World Health Organ 1966;35:691-708.
- Raymond M. PROBIT, CNRS-UMII, License L93019. Avenix, 34680 St. Georges dOrques, France 1993.
- Finney DJ. Probit analysis. London: Cambridge University Press, 1971.
- Devonshire AL, Moores GD. A carboxylesterase with broad substrate specificity causes organophosphate, carbamate and pyrethroid resistance in peach-potato aphids (Myzus persicae). Pest Biochem Physiol 1982;18:235-46.
- Bisset JA, Rodríguez, Dayamí L. Determinación de los mecanismos de resistencia en Culex quinquefasciatus Say 1823 y su implicación operacional en el uso correcto de insecticidas para el control. Rev Cubana Med Trop 1994;46:108-14.
- Rodríguez MM, Bisset JA, Mastrapa L, Díaz C. Asociación de la resistencia a insecticidas organofosforados, carbamatos y piretroides con los mecanismos de resistencia observados en cepas de Culex quinquefasciatus de Ciudad de La Habana. Rev Cubana Med Trop 1995;47:154-60.
Recibido: 11 de abril de 1997. Aprobado: 12 de diciembre de 1997.
Lic. María Magdalena Rodríguez. Instituto de Medicina Tropical "Pedro Kourí". Apartado 601, Marianao 13, Ciudad de La Habana, Cuba.

