










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Medicina Tropical
versión impresa ISSN 0375-0760versión On-line ISSN 1561-3054
Rev Cubana Med Trop v.56 n.1 Ciudad de la Habana ene.-abr. 2004
Comparación morfológica e histológica del tubo digestivo de Gambusia puncticulata y Girardinus metallicus (Cyprinodontiformes: Poeciliidae), peces utilizados en el control biológico de mosquitos
Lic. Jinnay Rodríguez Rodríguez,1 Lic. Efraín González,2 Ing. Natividad Hernández Contreras,3 Dra. Virginia Capó4 y Dr. Israel García5
Resumen
Se describió y comparó la morfología e histología del tubo digestivo de Gambusia puncticulata y Girardinus metallicus, peces utilizados en el control biológico de larvas de mosquitos en Cuba, y se relacionó con los hábitos alimentarios de cada especie. Gambusia puncticulata tiene un intestino corto y grueso, característico de las especies carnívoras y Girardinus metallicus posee un intestino alargado y delgado arrollado en forma de espiral como corresponde a los peces omnívoros. Se observó que el patrón histológico del intestino en ambas especies es muy simple, sin embargo, Gambusia puncticulata presentó gran cantidad de microvellosidades de tipo ramificadas, al compararla con Girardinus metallicus, lo que aumenta de esta forma el rendimiento digestivo de su relativamente más corto, tubo digestivo.
DeCS: CONTROL BIOLOGICO DE VECTORES/ métodos; CONTROL DE MOSQUITOS; PECES; CONTENIDO DEL TRACTO DIGESTIVO; MORFOLOGIA; HISTOLOGIA; CUBA.
Para combatir los mosquitos de importancia médico-veterinaria en Cuba se utilizan varias especies de peces larvívoros, dentro de los cuales se incluyen Girardinus metallicus y Gambusia puncticulata, sin embargo se señala que esta última es una especie carnívora y por lo tanto un magnífico biorregulador y Girardinus metallicus un pez omnívoro con menor capacidad para realizar esa función.1
Según la literatura, la morfología del tracto digestivo de los peces es muy variable, ilustrando la diversidad de regímenes alimentarios y modos de vida de estos.2 En el estudio de la morfología interna de los peces larvívoros cubanos, solo se conoce el trabajo de Poey3 referente a las características internas de los géneros Gambusia, Girardinus y Limia.
Numerosos estudios se han realizado sobre las características histológicas del tubo digestivo de varias especies de peces con valor comercial (Pereyra LA, Domitovic HA, Roux JP, Sampietro JC. Microscopia electrónica de barrido [MEB] del aparato digestivo del pacú [Piaractus mesopotamicus, Holmberg, 1887] [Pisces, Serrasalmidae]. Acta de Reunión de Comunicaciones científicas y tecnológicas de la UNNE, 2000).4 No se han encontrado trabajos de este tipo en los peces utilizados en el control biológico de mosquitos. Por las razones expuestas, el objetivo de este trabajo fue describir y comparar la morfología e histología del tubo digestivo de Gambusia puncticulata y Girardinus metallicus, y relacionarla con sus hábitos alimentarios.
Para el estudio se utilizaron 6 ejemplares de cada especie, los cuales fueron capturados en un canal de agua dulce en Batabanó, Provincia Habana, con un jamo de malla plástica y trasladados vivos al laboratorio en bolsas de nailon. Las especies de peces fueron mantenidas en peceras independientes, alimentándose con vegetación acuática y con larvas de Culex quinquefasciatus.
Tras ser sacrificados se procedió a disecar cada ejemplar mediante una incisión longitudinal que se inició en el ano y concluyó debajo de la mandíbula inferior. A continuación se extrajeron los órganos de la cavidad abdominal y se fijaron en Bouin acuoso. Se utilizaron técnicas histológicas de rutina donde los tejidos fijados fueron embebidos en parafina, cortados a 6 micrómetros con un micrótomo manual y teñidos con la técnica de hematoxilina-eosina,5 observándose en microscopio clínico con magnificación de hasta 1 000 x. Para determinar los tipos de tejidos y sus células componentes se siguió la terminología propuesta por Eliséiev y otros.6
A simple vista G. puncticulata posee un sistema digestivo en forma de conducto carente de diferenciación morfológica. No posee estómago, presenta un intestino y un hepatopáncreas. El intestino es corto y grueso, característico de las especies carnívoras, coincidiendo con lo planteado por Poey.3 El patrón histológico es muy simple, se observan al microscopio 2 capas: mucosa, capa muscular externa. En la figura 1 A se observa el tipo de epitelio de la parte más inicial, el cual es plano pseudoestratificado con células caliciformes y fibra muscular, lisa y estriada. Se continúa imperceptiblemente con epitelio cilíndrico pseudoestratificado con células caliciformes y presencia de fibra muscular lisa. La membrana basal de la mucosa se evagina originando gran cantidad de vellosidades ramificadas (fig 1 B, C), y vellosidades más simples y bajas que se van alternando con células caliciformes (fig. 1D).

A: se observa epitelio plano pseudoestratificado con fibra muscular lisa y estriada y epitelio cilíndrico pseudoestratificado, 400x; B, C: mucosa intestinal con vellosidades ramificadas 400x, 1 000x; D: se muestran vellosidades simples con células caliciformes 1 000x.
Fig 1. Epitelio intestinal de Gambusia puncticulata.
A todo lo largo del intestino pero con más intensidad en la porción media, se encuentran células con gránulos acidófilos similares a las células de Paneth localizadas en el epitelio de la mucosa. Estos gránulos densos son intensamente refringentes y con eosina se tiñen de color rojo brillante.
Macroscópicamente G. metallicus posee igualmente un sistema digestivo en forma de conducto carente de diferenciación morfológica, típico de los teleósteos vivientes más primitivos. No posee estómago, presenta un intestino y un hepatopáncreas. El intestino es alargado y delgado arrollado en forma de espiral, como corresponde a los peces omnívoros coincidiendo con lo descrito por Poey.3 En el patrón histológico del tubo digestivo se destacan 3 capas: mucosa, submucosa y muscular. Como se muestra en la figura 2 la membrana basal de la mucosa se evagina originando vellosidades de diferente longitud y forma: romas cortas (fig. 2 A), largas (fig. 2 C, D) y ramificadas (fig. 2B). De forma esquemática, se puede decir que en la primera parte del tubo digestivo se observan vellosidades cortas romas, en la parte media vellosidades largas y al final vellosidades ramificadas. La mucosa está constituida por epitelio cilíndrico pseudoestratificado con células caliciformes. También presenta células con gránulos acidófilos similares a las células de Paneth en la submucosa intercalada a todo lo largo del tubo digestivo.
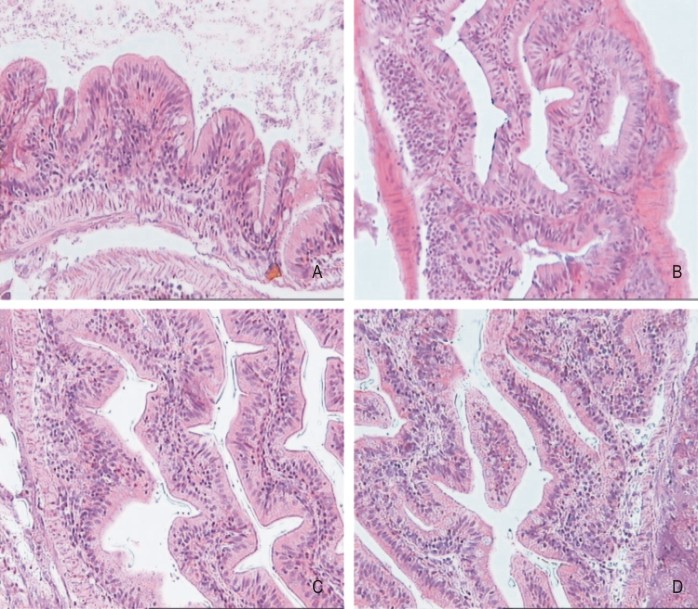
A: vellosidades romas cortas; B: vellosidades ramificadas; C, D: vellosidades largas.
Fig. 2. Epitelio intestinal de Girardinus metallicus, 400x.
El patrón histológico del intestino de ambas especies es muy simple en correspondencia con su escaso desarrollo morfológico, en ninguno de los casos se observa esófago ni estómago. En las 2 especies se presentan microvellosidades, como ocurre en forma similar en otros peces (Pereyra LA, Domitrovic HA, Sampietro JC. Microscopia electrónica de barrido en el tubo digestivo del Surubí [Pseudoplatystoma coruscans y Pseudoplatystoma fasciatum] [Pisces: Siluriformes]. Acta de Reunión de Comunicaciones Científicas y tecnológicas de la UNNE. 1999; Tomo IV:35-8). Sin embargo, Gambusia puncticulata presenta gran cantidad de microvellosidades de tipo ramificadas, al compararla con Girardinus metallicus. Esto coincide con Kuzmina7 el cual plantea que los peces carnívoros presentan un gran desarrollo de la superficie intestinal, cargada de enzimas proteolíticas; esto aumenta el rendimiento de sus relativamente más cortos, tubos digestivos.
El epitelio que se identifica en el intestino de ambas especies es pseudoestratificado cilíndrico, similar a lo reportado por González (González AR. Morfometría del tracto digestivo en especies cubanas del género Lucífuga [Ophidiiformes: Bythitidae]. Trabajo de Diploma, Universidad de La Habana, Facultad de Biología. 1986.) para otros peces; se señala además, presencia de células caliciformes coincidiendo con Bertin8 quien plantea que estas células son muy numerosas en los peces.
Los resultados obtenidos en el presente trabajo demuestran que existe una diferenciación morfológica e histológica del tubo digestivo de G. puncticulata y G. metallicus, relacionado con sus hábitos alimentarios, lo que hace superior a G. puncticulata sobre G. metallicus como biorregulador, y por lo tanto se recomienda esta especie como predadora de larvas de mosquitos de importancia médico epidemiológica en Cuba.
Summary
The morphology and histology of the digestive tract of Gambusia puncticulata and Girardinus metallicus fishes used in the biological control of mosquito larvae in Cuba, were described and compared. They were related to the food habits of each species. Gambusia puncticulata has a short and thick intestine, which is characteristic of the carnivorous species, whereas Girardinus metallicus has an elongate and thin spiral rolled up intestine as it corresponds to the omnivorous fishes. It was observed that the histological pattern of the intestine in both species is very simple; however, Gambusia puncticulata presented a great number of ramified microhairiness on comparing it with Girardinus metallicus, which increases this way the digestive yield of its relatively shorter digestive tract.
Subject headings: PEST CONTROL, BIOLOGICAL/methods; MOSQUITO CONTROL; FISHES; DIGESTIVE TRACT CONTENTS; MORPHOLOGY; HISTOLOGY; CUBA.
Referencias bibliográficas
- Koldenkova L, García I. Clave pictórica para las principales especies de peces larvívoros de Cuba. Nueva Gerona: Poligráfico Pablo de la Torriente Brau; 1990.p.56.
- Espinosa de los Monteros J, Labarta U. Nutrición en acuicultura I. Madrid;1987.p.272.
- Poey AF. Los Guajacones. En: Memorias sobre la historia natural de la Isla de Cuba. La Habana: Imp. Viuda de Barcina;1858.p.374-87.
- Flores Quintana C, Pereyra LA, Domitrovic HA, Roux JP, Sampietro JC. Estructura histología del aparato digestivo del Surubí (Pseudoplatystoma coruscans y Pseudoplatystoma fasciatum) (Pisces, Siluriformes). Reunión de Comunicaciones Científicas y Tecnológicas UNNE. Actas 1987;2:17-20.
- Klein A. Manual de métodos parasitológicos e histológicos en Piscicultura. Proyecto FAO/ PNND/URN/78/007. Asistencia al INAPE. Montevideo Uruguay, 1982:49.
- Eliséiev V G, Afanásiev Yu I, Yúrina NA. Histología. Editorial Mir. Moscú, 1985:583.
- Kuzmina VV. Membrane digestion in cyclostomes and fishes. J Ichthyol 1978;18(4):599-611.
- Bertin L. Ecologie en Grassé, pp (ed) Traité de zoologié, tomo XIII, fasc. 3: Agnates et poissons, Masson. París, 1958:1884-933.
Recibido: 10 de noviembre de 2003. Aprobado: 8 de diciembre de 2003.
Lic. Jinnay Rodríguez Rodríguez. Instituto de Medicina Tropical Pedro Kourí. Apartado 601, Marianao 13, Ciudad de La Habana, Cuba. Teléf: 2020426. Correo electrónico: jinnay@ipk.sld.cu
1 Licenciada en Biología. Aspirante a Investigadora.
2 Licenciado en Tecnología de la Salud.
3 Ingeniera Agrónoma.
4 Doctora en Ciencias Médicas.
5 Doctor en Ciencias Biológicas.

