










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Medicina Tropical
versión On-line ISSN 1561-3054
Rev Cubana Med Trop v.59 n.3 Ciudad de la Habana sep.-dic. 2007
ARTÍCULO ORIGINAL
Subtipos de virus dengue serotipos 2, 3 y 4 aislados en el Departamento de Santander, Colombia
Subtypes of dengue virus serotypes 2, 3 and 4 isolated in Santander District, Colombia
Dr. Fabián M. CortésI; Lic. Sergio Y. GómezII; Dra. Raquel E. OcazionezIII
I Candidato a Maestro en Ciencias Básicas Biomédicas. Universidad de Antioquia, Medellín, Colombia.
II Licenciado en Microbiología. Investigador Auxiliar. Centro de Investigaciones en Enfermedades Tropicales. Universidad Industrial de Santander.
III Doctora en Virología. Profesora Titular. Investigadora Titular. Universidad Industrial de Santander.
RESUMEN
Se analizaron virus de los serotipos 2, 3 y 4 que circularon en el Departamento de Santander, Colombia, entre 1998-2004. Identificar el subtipo de un serotipo del virus del dengue es una herramienta para vigilar factores de riesgo de severidad, porque las cepas se diferencian en el potencial para causar dengue hemorrágico. Se usó la técnica de amplificación simultánea de secuencias reconocidas por endonucleasas de restricción o restriction site-specific polymerase chain reaction (RSS-PCR) y el subtipo se determinó comparando el patrón electroforético del aislado local con el del virus de referencia. Virus del serotipo 2 resultaron subtipo A igual a un aislado de Tailandia (1996) y uno de Puerto Rico (1986); los del serotipo 3 fueron subtipo C como virus de"Sri-Lanka"(1990), Honduras (1995) y Puerto Rico (2000); los del serotipo 4 resultaron una variante del subtipo B igual que un virus de Puerto Rico (1987) y uno de Tahití (1985). Se confirmó la presencia en Colombia de los subtipos de virus dengue que circulan en América.
Palabras clave: Genotipo, dengue, serotipo, flavivirus, RSS-PCR.
SUMMARY
Virus serotypes 2, 3 and 4 that had circulated in Santander District, Colombia in the period 1998-2004 were analyzed. Identifying the subtype of a dengue virus serotype is a useful tool for surveillance of severe risk factors because the strain potential to cause hemorrhagic dengue makes the difference among them. Simultaneous sequence amplification technique known as restriction site specific-polymerase chain reaction (RSS-PCR) was used to determine the subtype by comparing the electrophoretic pattern of the local isolate to the reference virus. Virus serotype 2 corresponded to subtype A similar to the one isolated in Thailand (1996) and to the other isolated in Porto Rico (1986); virus serotypes 3 were of subtype C like the virus found in Sri Lanka (1990), Honduras (1995) and Porto Rico (2000); virus serotypes 4 were a variant of subtype B similar to a virus from Porto Rico (1987) and to another virus from Tahiti (1985). The study confirmed the presence in Colombia of dengue virus subtypes circulating now in the Americas.
Key words: Genotype, dengue, serotype, flavivirus, RSS-PCR.
INTRODUCCIÓN
El dengue es causado por el virus del mismo nombre y es la enfermedad transmitida por un artrópodo más frecuente en países tropicales. En la mayoría de los casos se manifiesta como un síndrome leve autolimitado conocido como fiebre dengue (FD) y en algunos de forma severa o fiebre hemorrágica del dengue (FHD), que puede llegar a ser fatal cuando se acompaña de choque hipovolémico (FHD/SCD). En Colombia, entre 2000-2004 se reportaron 236 319 casos de dengue a la Organización Panamericana de la Salud.1
Existen 4 serotipos del virus del dengue, conocidos como DEN-1, 2, 3 y 4; cada uno agrupa variantes genéticas de aislados. La divergencia entre virus del mismo serotipo se detectó comparando secuencias completas o parciales de los genes E o NS1. De esta manera, virus DEN-1 y DEN-2 se han agrupado en 5 subtipos,2 DEN-3 en 43 y DEN-4 en 2 subtipos.4 Otros métodos diferentes a la secuenciación nucleotídica se han usado para genotipificar virus dengue, entre estos, la amplificación simultánea de secuencias reconocidas por endonucleasas de restricción o restriction site specific polymerase chain reaction (RSS-PCR).5,6 La técnica permite identificar homología o divergencia genética entre aislados comparando el patrón electroforético de los fragmentos amplificados. Así, los virus DEN-1 se agrupan en 3 subtipos (A-C), DEN-2 en 7 (A - G), DEN-3 en 3 (A - C) y DEN-4 en 2 (A y B). En cada subtipo RSS-PCR se agrupan aislados de localidades y año de aislamiento similares, que corresponden a 1 de los subtipos generados por secuenciación nucleotídica o más.
Varios estudios han demostrado que la frecuencia de casos hemorrágicos durante una epidemia de dengue depende de la cepa del virus, por lo tanto, el monitoreo de la distribución temporal de serotipos y subtipos en áreas endémicas provee información del riesgo de FHD. El DEN-2 es el serotipo que con mayor frecuencia produce casos severos seguido por el DEN-3, DEN-1 y DEN-4, pero existen diferencias en el potencial virulento entre cepas.7 Los virus DEN-2 subtipo III y DEN-3 subtipo III son los que más se aíslan de casos severos.8,9
En Colombia, la presencia del virus dengue se detectó por primera vez en los años setenta cuando se aislaron los serotipos 1, 2 y 3, y posteriormente, en 1986 con la introducción del serotipo 4.10 Desde entonces los virus se continúan aislando aunque el DEN-3 estuvo ausente entre 1978 y 2000.11 Algunas cepas de los serotipos 1, 2 y 4 se han tipificado por secuenciación nucleotídica, resultando de los subtipos III, III y II, respectivamente.8,12,13 De los virus del serotipo 3 no hay información documentada del subtipo generado por análisis de secuencias. No obstante, algunas cepas que circularon en el Departamento de Santander luego de su reintroducción en 2001, fueron tipificadas usando la RSS-PCR y resultaron del subtipo C o"Sri-Lanka".11 El subtipo de virus serotipos 2 y 4 que han circulado en Santander no se conoce. El objetivo de este estudio fue tipificar virus aislados entre 1998 y 2004 en períodos con epidemia y sin esta, usando la RSS-PCR, incluidas cepas del serotipo 3 diferentes a las analizadas previamente.
MÉTODOS
Área del estudio
Santander es uno de los 32 departamentos de Colombia, ubicado en la región nororiental cerca de la frontera con Venezuela. Está conformado por 87 municipios en un área de 165 000 km2 y 2 086 459 habitantes (2005). Su capital es Bucaramanga, localizada a 380 km de Bogotá, la capital del país.
Virus
Las cepas de cada serotipo incluidas en el estudio, el año de aislamiento y la fuente se describieron en la tabla 1. Los virus fueron aislados del suero de pacientes con dengue en células de mosquito Aedes albopictus (clon C6/36-HT) como previamente se describió.11 Para el estudio, las cepas locales y las de referencia se replicaron de nuevo en las mismas células por 7 d a 32 °C. El sobrenadante de los cultivos se colectó, se clarificó por centrifugación y luego de la adición de suero bovino fetal (20 %) se almacenó a -70 °C hasta usarlo.
Extracción de ARN viral y síntesis de ADN
El sobrenadante (125 µL) de cultivos infectados se mezcló con 375 µL de Trizol (Invitrogen Corp.) y la mezcla se incubó a temperatura ambiente por 5 min. Al término se adicionaron 100 µL de cloroformo y después de extracción se centrifugó a 12 000 rpm. La fase acuosa recuperada se mezcló con 250 µL de isopropanol y luego de centrifugación a 12 000 rpm el precipitado se lavó con 500 µL de etanol 75 % y se secó a temperatura ambiente. El ARN viral se resuspendió en 50 µL de agua tratada con dietil-pirocarbonato (DEPC) y luego se almacenó a - 70 °C. Para síntesis de ADN se usaron 5 µL del ARN extraído que se adicionaron a una mezcla de reacción que contenía 200 µM de cada dNTP, 0,5-1,0 µM de iniciadores anti-sentido,5,6 1 mM de MnCl2, 0,25 U de enzima Tth DNA Polimerasa (Promega), 2,5 µL de buffer, en un volumen final de 25 µL. La reacción se llevó a cabo a 90 °C por 2 min y luego a 60 °C por 30 min.
RSS - PCR
Virus de referencia y aislados locales de cada serotipo fueron procesados simultáneamente, usando una modificación de los protocolos descritos por Harris y otros5 y Miagostovich y otros.6 Brevemente, 10 µL de ADN viral copia se adicionaron a una mezcla de reacción que contenía 2 mM de MgCl2, 200 µM de cada dNTP, 0,35-0,8 µM de los iniciadores, 5 µL de buffer quelante, 0,25 U de Tth DNA Polimerasa (Promega), 5 µL de buffer de enzima, en un volumen final de 50 µL. La amplificación se realizó por 32 ciclos de 94 °C por 1 min, 54 °C por 1 min y 72 °C por 2 min con una extensión final a 72 °C por 10 min. El ARN de cada cepa fue amplificado por más de una vez para confirmar la reproducibilidad del resultado.
Subtipo RSS- PCR
Los productos de la amplificación se analizaron mediante electroforesis en gel de agarosa 2 % disuelto en tampón trisborato-EDTA (Tris-HCl 89 mM pH 8, ácido bórico 89 mM, EDTA 2,5 mM) y usando un estándar de tamaños de 100-pb (DNA ladder. Promega). En un mismo gel se colocaron 15-20 µL tanto de los amplificados obtenidos con virus locales como con los de referencia del mismo serotipo. Para determinar el subtipo se comparó el patrón electroforético (tamaño y número de fragmentos) generado por la cepa local versus la prototipo.
RESULTADOS
Virus DEN-2
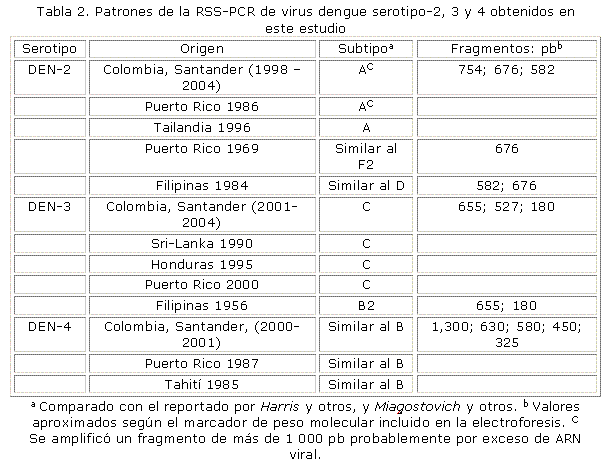
Se visualizaron 3 patrones electroforéticos que fueron iguales o similares a los generados por virus de los subtipos A, D y F2, según el número y tamaño de los fragmentos amplificados (tabla 2). Las cepas de Santander generaron el patrón del subtipo A como un virus de Puerto Rico aislado en 1986 y uno de Tailandia de 1996. Los virus locales resultaron diferentes a un aislado de Filipinas en 1986 (similar al subtipo D) y uno de Puerto Rico de 1969 (similar al subtipo F2). Un ejemplo representativo de los patrones electroforéticos con virus DEN-2 se muestra en la figura 1.
Virus DEN-3
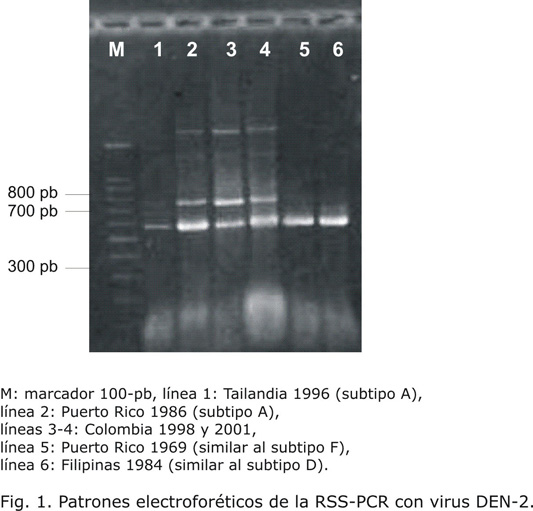
La amplificación del genoma de los virus incluidos en el estudio resultó en los patrones electroforéticos de los subtipos B2 y C (tabla 2). Los virus de Santander resultaron del subtipo C igual al de un virus de "Sri-Lanka" (India) aislado en 1990, uno de Puerto Rico de 2000 y uno de Honduras de 1995. El patrón generado por los virus locales fue diferente al de uno de Filipinas aislado en 1956 (cepa H 87) previamente reportado como subtipo B2. Un ejemplo representativo de los patrones electroforéticos de virus DEN-3 se muestra en la figura 2.
Virus DEN-4
Las cepas locales y de referencia generaron un único patrón electroforético que fue similar al del subtipo B previamente reportado por Miagostovich y otros.6 Se observó variación en el tamaño de 2 de los 5 fragmentos amplificados. Esto es producto de 580 y 320 pb vs. 600 y 280 pb de los esperados (tabla 2). Las cepas de Santander resultaron iguales a un virus de Puerto Rico aislado en 1987 y uno de Tahití de 1985. Un ejemplo representativo de los patrones electroforéticos generados por virus DEN-4 se muestra en la figura 3.
DISCUSIÓN
De todos los virus del dengue, las cepas del serotipo 2 asiático son las que con mayor frecuencia producen casos severos.7,8,11 La circulación permanente en países de América del subtipo III de este serotipo, conocido como "Americano-Asiático"o "Jamaica", se ha considerado una de las causas del incremento notorio de la FHD en la región.8 Este virus produjo la epidemia de mayor severidad que se haya registrado en Cuba ocurrida en Santiago en 1997, con 205 casos de FHD de los cuales 12 murieron.14,15 Los virus DEN-2 subtipo III se agrupan en el subtipo A de acuerdo con el patrón electroforético de la RSS-PCR.5 En Santander, el DEN-2 ha sido el serotipo prevalente, su dominancia entre 2000-2001 estuvo asociada con incremento de casos hemorrágicos. De los infectados con el virus, 27,5 % desarrollaron FHD y 13,7 % fueron fatales.11 Los resultados de este estudio muestran que los aislados entre 1998 y 2004 fueron subtipo A donde se agrupan cepas del genotipo "Americano-Asiático". Este hallazgo fue el mismo que con virus de Nicaragua de la epidemia de 1998 y de Brasil de 2000 a 2001 tipificados por la RSS-PCR.16,17
De los virus DEN-3, las cepas del subtipo III o "Sri-Lanka"se consideran las más virulentas3 y se agrupan en el subtipo C según la técnica de la RSS-PCR.5 Como se muestra en la figura 2, los aislados colombianos fueron subtipo C y este mismo resultado se reportó con virus del Brasil de la epidemia de 2000 y de Nicaragua de 1998.16,17 La circulación permanente del subtipo III en la isla de"Sri-Lanka"desde 1990 se ha relacionado con la aparición de la FHD9 y su introducción en Brasil en 2000 produjo la epidemia con más casos severos y fatales que se haya registrado en ese país.18 En Colombia, el DEN-3 estuvo ausente entre 1978 y 2000, reapareciendo en 2001 por primera vez en Santander. Aquí, la dominancia del virus luego de su introducción se asoció con disminución de la FHD, 89,1 % de los infectados con este serotipo desarrollaron FD.11 Una situación similar se vio en Venezuela luego de la aparición del virus en 2000.19
La causa que explique la diferencia entre la severidad de las epidemias por DEN-3 subtipo III en países de América se desconoce. Diferentes factores relacionados con el individuo y diferencias genéticas de los virus pueden haber contribuido. En relación con el virus, es conocido que una simple mutación no sinónima puede causar incremento de virulencia o atenuación de una cepa de dengue. En el gen E de virus DEN-3 se han reportado sustituciones de residuos de la proteína que participan en el reconocimiento antígénico y esto podría resultar en cambios de susceptibilidad del virus a eliminación por la respuesta inmune.20 Algunas evidencias sugieren evolución genética in situ de la cepa del virus DEN-3 introducida en Nicaragua y Panamá en 1994, que pudieron modificar la virulencia y originaron cepas con diferente potencial para producir casos severos.19
El virus DEN-4 fue introducido en América en 1986 y de los 2 subtipos de este serotipo el II ha sido el único detectado.4,13 Los aislados de América se agrupan en el subtipo B usando la técnica de la RSS-PCR.6 En este estudio se analizaron cepas de Santander, Puerto Rico y Tahití que produjeron un mismo patrón electroforético, diferente en el tamaño de 2 de los 5 fragmentos esperados para el subtipo B (tabla 2). Para confirmar el subtipo de virus de Santander, se hizo secuenciación nucleotídica de un segmento del gen prM de uno de los aislados en 2000 (Laboratorio de Enfermedades Infecciosas y Transmitidas por Vectores, Universidad del Estado de Colorado, Fort Collins, Colorado, EE. UU.). Como se esperaba, el virus resultó subtipo II como virus de otros de países de América (Diaz F. Epidemiología molecular de virus dengue de América. En: 1er. Simposio Nacional de Virología, Medellín, Colombia, 2004). El subtipo II también fue el de una cepa colombiana aislada en 1996 en un departamento distinto a Santander.13 A diferencia de los virus serotipos 2 y 3, el 4 se considera el que menos casos hemorrágicos produce.7 En Santander, fue el serotipo menos aislado entre 1998 y 2004; su emergencia no pareció contribuir a la FHD.11
Varios factores influyen en la reproducibilidad de los patrones de la RSS-PCR; principalmente mutaciones en la secuencia blanco y condiciones del ensayo como temperatura, concentración y tipo de reactivos. Como resultado puede ocurrir incremento o disminución del número y tamaño de los productos amplificados y esto requiere que el ensayo se implemente usando virus prototipo de 2 patrones reproducibles o más, con diferentes protocolos.5 En este estudio no se amplificaron fragmentos menores a 500 pb de virus DEN-2, subtipos F2 y D, y se detectaron cambios en el tamaño de amplificados del DEN-4 (tabla 2). Este resultado pudo deberse a variaciones en el tipo de reactivos y porque algunas de las cepas de referencia fueron diferentes a las usadas por los autores que originalmente reportaron el método. Variaciones en los patrones de cepas de un mismo subtipo como las encontradas en este estudio han sido reportadas por otros.16,17
Aunque la secuenciación nucleotídica ha sido el método de referencia para tipificar virus dengue, la necesidad de personal especializado, equipo, materiales y reactivos costosos, dificulta su uso rutinario en laboratorios encargados de la vigilancia virológica. Adicionalmente, la información, en general, se conoce tiempo después de la epidemia cuando el virus se aisló. La RSS-PCR es una alternativa para tipificar virus en tiempo real en laboratorios poco desarrollados, por su bajo costo y simplicidad. La utilización de esta técnica permitió conocer el subtipo del DEN-3 durante la epidemia que causó en Santander en 2001.11
Los resultados de este estudio confirman la presencia en Colombia del subtipo A ("Americano-Asiático") del virus DEN-2, del subtipo C ("Sri-Lanka") del DEN-3 y del subtipo B (II) del DEN-4, iguales a virus aislados en otros países de América. Por otro lado, muestran la utilidad de la técnica de la RSS-PCR para soporte rutinario de la vigilancia virológica del dengue en áreas endémicas.
AGRADECIMIENTOS
Al doctor Vancen Vorndam (CDC, Puerto Rico) por su colaboración con los virus de referencia y al doctor Javier Díaz por el secuenciamiento de un aislado del virus DEN-4. Este trabajo fue financiado por el Instituto Colombiano de Ciencia y Tecnología (Colciencias) proyecto 11020-413042.
REFERENCIAS BIBLIOGRÁFICAS
1. Pan American Health Organization. Number of Reported Cases of Dengue and Dengue Hemorrhagic Fever (DHF). Region of the Americas (by country and subregion). Acceso el 8 de agosto de 2006. Disponible en: http://www.paho.org/english/ad/dpc/cd/dengue
2. Rico-Hesse R. Molecular evolution and distribution of dengue viruses type 1 and 2 in nature. Virology. 1990;174:479-93
3. Lanciotti RS, Lewis JL, Gubler DJ, Trent DW. Molecular evolution and epidemiology of dengue-3 viruses. J Gen Virol. 1994;75:65-75.
4. Lanciotti RS, Gubler DJ, Trent DW. Molecular evolution and phylogeny of dengue-4 viruses. J Gen Virol. 1997;78:2279-84.
5. Harris E, Sandoval E, Xet-Mull AM, Johnson M, Riley LW. Rapid subtyping of dengue viruses by restriction site-specific (RSS)-PCR. Virology. 1999;253:86-95.
6. Miagostovich MP, dos Santos FB, Gutierrez CM, Riley LW, Harris E. Rapid subtyping of dengue virus serotypes 1 and 4 by restriction site-specific PCR. J Clin Microbiol. 2000;38:1286-9.
7. Vaughn DW, Green S, Kalajanarooj S, Innis BL, Nimmannitya S, Suntayakorns S, et al. Dengue viremia titer, antibody, response pattern and virus serotype correlate with disease severity. J Infect Dis. 2000;181:2-9.
8. Rico-Hesse R, Harrison L, Salas R. Tovar D, Nisalak A, Ramos C, et al. Origins of dengue type 2 viruses associated with increased pathogenicity in the Americas. Virology. 1997;230:244-51.
9. Messer BW, Vitarana UT, Sivananthan K, Vigala JE, Preethimala LD, Ramesh R, et al. Epidemiology of dengue in"Sri-Lanka"before and after of emergence of epidemic dengue hemorrhagic fever. Am J Trop Med Hyg. 2002;66:765-73.
10. Boshell J, Groot H, Gacharna M, Márquez G, González M, Gaitan MO, et al. Dengue en Colombia. Biomédica. 1986;6:101-2.
11. Ocazionez R, Cortés F, Villar L, Gómez S. Temporal distribution of dengue virus serotypes in Colombian endemic area and dengue incidence. Re-introduction of dengue-3 associated to mild febrile illness and primary infection. Mem Inst Oswaldo Cruz. 2006;101:725-31.
12. Gonçalvez A, Escalante A, Pujol FH, Ludert J, Tovar D, Salas R, et al. Diversity an evolution of the envelope gene of dengue virus type 1. Virology. 2002;303:110-9.
13. Díaz FJ, Black IV WC, Farfán-Ale JA, Loroño-Pino MA, Olson KE, Beaty BJ. Dengue virus circulation and evolution in Mexico: A Phylogenetic perspective. Arch Med Res. 2006;37:760-73.
14. Valdes L, Guzmán MG, Kourí G, Delgado J, Carbonell I, Cabrera MV, et al. Epidemiology of dengue and hemorrhagic dengue in Santiago, Cuba 1997. Rev Panam Salud Publica. 1999;6:16-25.
15. Rodríguez-Roche R, Alvarez M, Gritsun T, Halstead S, Kouri G, Gould EA, et al. Virus evolution during a sever dengue epidemic in Cuba,1997. Virology. 2005;334:154-9.
16. Balmaseda A, Sandoval E, Pérez L, Gutiérrez CM, Harris E. Application of molecular typing techniques in the 1998 dengue epidemic in Nicaragua Am J Trop Med Hyg. 1999;61:893-7.
17. De Simone TS, Nogueira RM, Araújo ES, Guimarães FR, Santos FB, Schatzmayr HG, et al. Dengue virus surveillance: the co-circulation of DENV-1, DENV-2 and DENV-3 in the State of Rio de Janeiro, Brazil. Trans R Soc Trop Med Hyg. 2004;98:533-62.
18. Nogueira RM, Schatzmayr HG, Filippis AMB, Santos FV, Cunha RV, Coehlo JO, et al. Dengue Virus Type 3, Brazil, 2002. Emerg Infect Dis. 2005;11:1376-81.
19. Uzcategui NY, Comach G, Camacho D, Salcedo M, Cabello de Quintana M, Jimenes M, et al. Molecular epidemiology of dengue virus type 3 in Venezuela. J Gen Virology. 2003;84:1569-75.
20. Sánchez IJ, Ruiz BH. A single nucleotide change in the E protein gene of dengue virus Mexican strain affects neurovirulence in mice. J Gen Virology. 1996;77:2541-5.
Recibido: 7 de octubre de 2006.
Aprobado: 17 de enero de 2007.
Dr. Fabián M. Cortés. UIS sede Guatiguará, CINTROP, Km 2 Autopista Piedecuesta, Santander, Colombia. Fax: 57-7-6540177. Correo electrónico: fabiancortesm@yahoo.com
Universidad de Antioquia, Medellín, Colombia.
Centro de Investigaciones de Enfermedades Tropicales, Universidad Industrial de Santander, Bucaramanga, Colombia.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}