










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Medicina Tropical
versión impresa ISSN 0375-0760versión On-line ISSN 1561-3054
Rev Cubana Med Trop v.59 n.3 Ciudad de la Habana sep.-dic. 2007
ARTÍCULO ORIGINAL
Determinación in vivo del papel de las enzimas esterasas y glutation transferasa en la resistencia a piretroides en Aedes aegypti (Diptera: Culicidae)
Determination in vivo of the role of esterase and gluthation transferase enzymes in pyrethroid resistance of Aedes aegypti (Diptera: Cullicidae)
Lic. María M. RodríguezI; Lic. Juan A. BissetII; Téc. Ditter FernándezIII
I Licenciada en Bioquímica. Investigadora Auxiliar. Instituto de Medicina Tropical ¨Pedro Kourí¨ (IPK).
II Doctor en Ciencias. Licenciado en Ciencias Biológicas. Investigador Titular. IPK.
III Técnica en Química. IPK.
RESUMEN
Se realizó un estudio in vivo a través del uso de 2 sinergistas, el trifenil fosfato (TFF) inhibidor específico de esterasas y el ácido etacrínico (AE), inhibidor específico de la enzima glutation transferasa (GST), para determinar si estas enzimas eran responsables de la resistencia a piretroides en Aedes aegypti. Para el trabajo se utilizaron 2 cepas de Aedes aegypti resistentes a insecticidas, una cepa que fue seleccionada con temefos por 6 generaciones de selección (SAN-F6) y otra con deltametrina por 12 generaciones de selección con este insecticida (SAN-F12), ambas resultaron ser resistentes a insecticidas piretroides. Se demostró a través del uso de los sinergistas TFF y AE que las enzimas esterasas y GST son responsables de la resistencia a los piretroides en estas cepas. Esos resultados demuestran la existencia de un fenómeno de resistencia cruzada y multirresistencia, lo cual debe tenerse en cuenta en las estrategias de uso de insecticidas para el control de este vector.
Palabras clave: Aedes aegypti, sinergistas, esterasas, acetilcolinesterasa, glutation transferasa.
SUMMARY
An in vivo study of two synergists, that is, Triphenil phosphate -specific esterase inhibitor- and ethacrynic acid specific gluthation transferase inhibitor- was performed to determine if these enzymes were responsible for pyrethroid resistance of Aedes aegypti. To this end, two insecticide resistant Aedes aegypti strains were used, one strain selected with temephos by six selection generations (SAN-F6) and the other strain with delmamethrin by 12 selection generations (SAN-F12), being both strains resistant to pyrethroid insecticices. Through the use of TPP and EA synergists, it was proved that esterase and gluthation-s-transferase (GST) enzymes were responsible for pryrethroid resistance of these strains. These results showed the existence of cross-resistance and multidrug resistance, which should be taken into account for insecticide use strategies aimed at vector control.
Key words: Aedes aegypti, sinergists, esterases, acetylcholinesterase, glutathion transferase.
INTRODUCCIÓN
El mosquito Aedes aegypti es el principal vector de fiebre amarilla, dengue y fiebre hemorrágica del dengue (FHD). Globalmente existe un estimado de 50 000 000 de casos de dengue al año, con 500 000 casos de FHD y más de 24 000 defunciones.1
Un control efectivo del vector es esencial para reducir la transmisión de dengue, pero Aedes aegypti ha desarrollado resistencia a una amplia variedad de insecticidas,2 lo que constituye el principal problema operacional que afecta su control.
La resistencia a piretroides en Aedes aegypti se ha asociado a un fenómeno de resistencia cruzada entre DDT y piretroides3 y se ha demostrado que este fenómeno está dado por el mecanismo de resistencia de knockdown, tipo gen Kdr.4 Más recientemente se ha asociado también la resistencia a piretroides con los mecanismos de acción metabólica, basado en el incremento de la actividad de algunas enzimas como son las esterasas y la GST.5,6 En Cuba se han seleccionado cepas de Aedes aegypti con genes para la resistencia a temefos7 y a deltametrina,8 ambas cepas muestran alta resistencia a piretroides.
El objetivo de este trabajo fue conocer, a través de estudios con sinergistas, el papel de los mecanismos de resistencia de acción metabólica, basado en la alta actividad de las enzimas esterasas y GST en la resistencia a piretroides en cepas de referencia de Aedes aegypti resistentes a piretroides.
MÉTODOS
Cepas
Se utilizaron 3 cepas:
San-F6: una cepa de Aedes aegypti seleccionada con temefos por 6 generaciones de selección con este insecticida.
SAN-F12: una cepa seleccionada con deltametrina por 12 generaciones de selección con este insecticida.
ROCKEFELLER: una cepa de referencia de Aedes aegypti susceptible a insecticidas, suministrada por el CDC, San Juan, Puerto Rico.
Insecticidas grado técnico utilizados
Piretroides: lambdacialotrina, cipermetrina, ciflutrina y deltametrina.
Bioensayos de larvas
En los bioensayos de la OMS,9 se realizaron 5 réplicas de cada concentración del insecticida (20 larvas por réplica), y se registró entre 2 y 98 % de mortalidad. Todas las soluciones se ajustaron a un volumen final de 1 mL con acetona. Esta concentración de acetona no causó mortalidad en los controles. La lectura de las mortalidades se realizó a las 24 h, se halló la CL50 y la CL90 con el programa probit-log.10
La acción de 2 sinergistas, trifenil fosfato (TFF) y ácido etacrínico (AE), fueron investigados mediante la exposición de las larvas de cuarto estadio a 2,5 mg/L de TFF y 5 mg/L de AE durante 4 h, previo a la adición de la solución del insecticida y se determinó la mortalidad después de 24 h de exposición; las concentraciones letales 50 (CL50) y 90 (CL90) se hallaron mediante el programa Probit-logaritmo.10 No existió mortalidad del sinergista solo a estas concentraciones. Se calculó el factor de sinergismo (FS), por la fórmula siguiente:
FS= CL50 insecticida sin sinergista/Cl50 insecticida más sinergista TFF o AE
RESULTADOS
Se determinó in vivo, la intervención de las enzimas esterasas o glutation transferasa (GST) en la resistencia a los piretroides lambdacialotrina, cipermetrina, ciflutrina y deltametrina, se utilizaron los sinergistas trifenil fosfato (TFF) y ácido etacrínico (AE). Estos sinergistas potencian la acción del insecticida inhibiendo de forma específica la actividad de esterasas (TFF) o de la GST (AE). El efecto de los sinergistas se expresó como factor de sinergismo (FS), el cual significa cuántas veces disminuye la concentración letal media (CL50) al aplicar el sinergista.
En la tabla 1 se observó que la cepa SAN-F6, la cual fue seleccionada con el insecticida temefos por 6 generaciones de selección, mostró resistencia a todos los piretroides (FR50 mayor que 10x), excepto para lambdacialotrina, al cual se observó un valor de FR50 menor que 5x (0,19x). Esta cepa mostró el valor mayor de FR50 para deltametrina (337,5 x), seguido por ciflutrina (34,61x) y cipermetrina (13,07x).
El valor de FS para lambdacialotrina resultó bajo (FS ≥ 5) para TFF (1,35x) y para el AE (1,08x); esto demuestra que esas enzimas no intervinieron en la resistencia al piretroide. Sin embargo, al aplicar el TFF la resistencia a cipermetrina disminuyó 18,47x, a ciflutrina 4 500x y a deltametrina 150x, estos son los valores correspondientes al FS mayores que 5x. Al aplicar el AE, se observó un efecto similar, el valor del FS resultó ser mayor que 5x para todos los piretroides, es decir disminuyó la resistencia a cipermetrina 3 225x, a ciflutrina 750x y a deltametrina 27x. Estos resultados indicaron que las enzimas esterasas y GST fueron responsables de la resistencia a piretroides en la cepa SAN-F6, porque al inhibir estas enzimas se potenció la acción de estos piretroides, lo cual se evidenció por la disminución de la CL50 para cada insecticida, cuando se aplicaron estos sinergistas, que incrementó el FS.
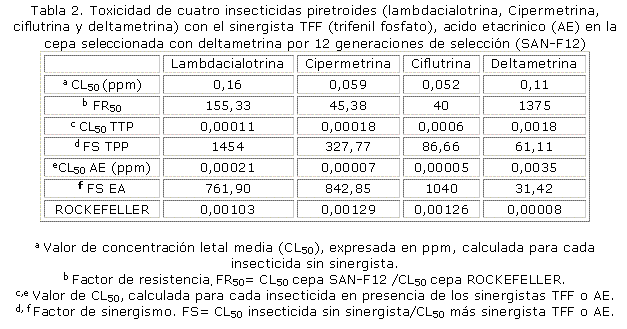
Este mismo análisis se realizó en otra cepa de Aedes aegypti que fue seleccionada con el piretroide deltametrina por 12 generaciones (SAN-F12). Esta cepa fue resistente a todos los piretroides (tabla 2), dado por el valor de FR50 mayor que 10x, se observó el mayor valor de FR50 para deltametrina (1 375x), seguido por lambdacialotrina (155,33x), cipermetrina (45,38x) y ciflutrina (40x). Para todos los piretroides, excepto para lambdacialotrina, se observó mayor valor de FS al aplicar el AE que al aplicar TFF, pero para ambos se observaron valores muy elevados. Al aplicar el TFF (inhibidor de esterasas), la resistencia disminuyó a lambdacialotrina 1 454x, a cipermetrina 327,77x, a ciflutrina (86,66x) y a deltametrina (61,11x). Al aplicar AE (inhibidor de la GST), la resistencia a lambdacialotrina disminuyó 761,90x, a cipermetrina 842,85x, a ciflutrina 1 040x y a deltametrina 31,42x. Estos resultados confirmaron que ambas enzimas, las esterasas y la GST intervinieron en la resistencia a piretroides en esta cepa.
DISCUSIÓN
Los resultados indicaron que la resistencia a piretroides observada tanto en la cepa SAN-F6 como en la cepa SAN-F12, se asoció a la incrementada actividad de las enzimas esterasas y glutation transferasa (GST). Durante el proceso de selección con temefos para la obtención de la cepa SAN-F6 aumentó la frecuencia de la actividad acrecentada de ambas enzimas y se infirió que la resistencia cruzada a piretroides podría estar asociada con el incremento de la actividad de la GST.7
En este trabajo se demostró a través de sinergistas, inhibidores específicos de esterasas (TFF) y de GST (AE), que ambas enzimas fueron responsables de la resistencia generada a los piretroides durante el proceso de selección con temefos. En la cepa SAN-F12 seleccionada con deltametrina por 12 generaciones de selección con este insecticida,8 también se demostró el fenómeno de multirresistencia, donde el incremento de la actividad de estas enzimas generó resistencia no solo a deltametrina, sino también a los otros piretroides evaluados, lambdacialotrina, cipermetrina y ciflutrina. De acuerdo con los resultados de Rodríguez y otros,8 la cepa SAN-F12 mostró resistencia a DDT, por lo que un mecanismo que también generó resistencia a los piretroides fue el gen Kdr, aunque este se encontró en esta cepa a baja frecuencia.
La resistencia cruzada entre DDT y piretroides está demostrada que existe en Aedes aegypti a través del mecanismo de knockdown o gen Kdr.4,11 Un segundo grupo de mecanismos de resistencia que existe con alta prevalencia, está basado en el incremento del metabolismo de los insecticidas. Existen 3 grupos de enzimas envueltas en el metabolismo de insecticidas, las esterasas, las monooxigenasas y la GST.5 La elevada actividad de estas enzimas ha sido asociada con la resistencia cruzada entre DDT y piretroides en algunas especies de insectos.5,6,12,13 Lumjuan y otros14 demostraron la presencia de 8 clases diferentes de GST en Aedes aegypti, el tipo GST epsilon 2 (GSTe2) en Aedes aegypti (AeGSTe2) se sobre-expresó en mosquitos resistentes a DDT y al piretroide permetrina. Existen reportes en la literatura donde implican las esterasas como mecanismo principal de resistencia a temefos.6,15-18
Se identificó en este trabajo cómo los mecanismos de acción metabólica basados en el incremento de la actividad de esterasas y GST intervienen tanto en la resistencia a temefos como a piretroides, lo cual hay que tener muy en cuenta porque se pueden generar fenómenos de resistencia cruzada y multirresistencia que afectan la efectividad de un producto químico, aun cuando se haya aplicado poco o no se haya aplicado nunca.
REFERENCIAS BIBLIOGRÁFICAS
1. WHO. Dengue and dengue haemorrhagic fever. Fact sheet No. 117. Geneva:World Health Organization; 2002.
2. ————. Resistance of vectors and reservoirs of disease to pesticides. Geneva: World Health Organization; 1986. (Tech. Rep. Ser. 737)
3. Chadwick PR, Invest JF, Bowron MJ. An example of cross-resistance to pyrethroids in DDT-resistant Aedes aegypti. Pestic Sci. 1977;15:112-20.
4. Soderlund DM, Knipple DC. The molecular biology of knockdown resistance to pyrethroid insectides. Insecticide Biochem Mol Biol. 2003;33:563-77.
5. Hemingway J, Hawkes NJ, McCarroll L, Ranson H. The molecular basis of insecticide resistance in mosquitoes. Insect Biochem Mol Biol. 2004;34:653-6.
6. Prapanthadara L, Promtet N, Koottathep S, Somboon P, Suwonkerd W, McCarroll L, et al. Mechanisms of DDT and permethrin resistance in Aedes aegypti from Chiang Mai, Thailand. Dengue Bull. 2002;26:185-9.
7. Rodríguez MM, Bisset JA, Ruiz M, Soca A. Cross-resistance to pyrethroid and organophosphate insecticides induced by selection with temephos in Aedes aegypti (Diptera: Culicidae) from Cuba. J Med Entomol. 2002;39:882-8.
8. Rodríguez MM, Bisset JA, De Armas Y, Ramos F. Pyrethroid insecticide-resistant strain of Aedes aegypti from Cuba induced by deltamethrin selection. J Am Mosq Control Assoc. 2005;21:437-45.
9. World Health Organization. Instructions for determining the susceptibility or resistance of mosquito larvae to insecticides. Geneva:WHO/VBC/81.80; 1981. p. 6.
10. Raymond M. Presentation d'une programme d'analyse log-probit pour microordinateur cahiers. Orstrom Sér Ent Méd Parasitol. 1985;23:117-21.
11. Brengues C, Hawkes NJ, Chandre F, Mccarroll L, Duchon S, Guillet P, et al. Pyrethroid and DDT cross-resistance in Aedes aegypti is correlated with novel mutations in the voltage-gated sodium channel gene. Med Vet Entomol. 2003;17:87-94.
12. Kasai S, Weerashinghe IS, Shono T, Yamagawa M. Molecular cloning, nucleotide sequence and gene expression of a cytochrome P450 (CYP6F1) from the pyrethroids resistance mosquito, Culex quinquefasciatus Say. Insect. Biochem Mol Biol. 2000;30:163-71.
13. Vaughan A, Hemingway J. Mosquito carboxylesterases Est alpha 2 (1) (A2). Cloning and sequence of the full-length cDNA for a major insecticide resistance gene worldwide in the mosquito Culex quinquefasciatus. J Biol Chem. 1995;270:17044-9.
14. Lumjuan N, McCarroll L, Prapanthadara L, Hemingway J, Ranson H. Elevated activity of an Epsilon class glutathione transferase confers DDT resistance in the dengue vector, Aedes aegypti. Insect Biochem Mol Biol. 2005;35:861-71.
15. Mazarri MB, Georghiou GP. Characterization of resistance to organophosphate, carbamate, and pyrethroid insecticides in field populations of Aedes aegypti from Venezuela. J Am Mosq Control Assoc. 1995;11:315-22.
16. Rodríguez MM, Bisset JA, Fernández D, Pérez O. Resistencia a insecticidas en larvas y adultos de Aedes aegypti: prevalencia de la esterasa A4 asociada con la resistencia a temefos. Rev Cubana Med Trop. 2004;56:54-60.
17. Bisset JA, Rodríguez MM, Fernández D, Pérez O. Estado de la resistencia a insecticidas y mecanismos de resistencia en larvas del municipio Playa, colectadas durante la etapa intensiva contra el Ae. aegypti en Ciudad de La Habana, 2001-2002. Rev Cubana Med Trop. 2004;56:61-6.
18. Braga IA, Lima JB, Soares S, Valle D. Aedes aegypti resistance to temephos during 2001 in several municipalities in the state of Río de Janeiro, Sergipe, and Alagoas, Brazil. Mem Inst Oswaldo Cruz. 2004;99:199-203.
Recibido: 27 de noviembre de 2006.
Aprobado: 12 de febrero de 2003.
Lic. María Magdalena Rodríguez. Instituto de Medicina Tropical "Pedro Kourí". AP 601. Marianao 13. Ciudad de La Habana, Cuba. Correo electrónico: mrodriguez@ipk.sld.cu


{kind=link}
{kind=link}