










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Medicina Tropical
versión impresa ISSN 0375-0760
Rev Cubana Med Trop vol.64 no.2 Ciudad de la Habana Mayo-ago. 2012
Propiedades biológicas de cepas de dengue virus-3 aisladas durante la epidemia ocurrida en La Habana, 2001-2002
Biological properties of virus dengue-3 strains isolated during the epidemic ocurred in Havana, 2001-2002
Dra. C. Rosmari Rodríguez-Roche,I MSc. Liudmila López Matilla,II Dra. C. Mayling Álvarez Vera,I MSc. Luis Morier Díaz,I Dra. C. María G. Guzmán TiradoI
I Instituto de Medicina Tropical "Pedro Kourí". La Habana, Cuba.
II Centro de Inmunología Molecular. La Habana, Cuba.
RESUMEN
Introducción: durante epidemias de dengue ocurridas en Cuba se ha observado de forma reiterada, un incremento de severidad clínica con la progresión de las epidemias en el tiempo, particularmente en infecciones secundarias. Se considera que este incremento pudiera estar relacionado con cambios genéticos del virus circulante.
Objetivo: estudiar algunas propiedades biológicas de cepas aisladas en diferentes momentos de la epidemia de La Habana, 2001-2002.
Métodos: se estudiaron 9 cepas de dengue virus-3, se evaluó el efecto citopatogénico y el crecimiento viral en las líneas celulares C6/36 HT y Vero, tamaño de las placas virales, sensibilidad a la temperatura, neurovirulencia en ratones lactantes y la influencia del pH en la unión del virus a la célula, así como en el medio de multiplicación de este.
Resultados: las cepas de dengue virus-3 resultaron más citopatogénicas en células Vero, sin embargo, en C6/36 HT se obtuvieron títulos superiores. El conjunto de cepas mostró reducción del título viral y del tamaño de placa al aumentar la temperatura y fueron poco neurovirulentas. En cuanto a la unión del virus a la célula, a pH básico se observaron los mejores títulos, mientras que a pH ácido solo se observó el crecimiento de algunas cepas aisladas al final de la epidemia. El medio de multiplicación del virus a pH 6,5-7 favoreció el crecimiento de las cepas del inicio de la epidemia, mientras que las cepas del final tuvieron títulos superiores a pH 7-8.
Conclusiones: se pudo comprobar la existencia de cambios fenotípicos entre cepas de diferentes momentos de la epidemia, que pudieran estar asociados con diferencias en cuanto a su adecuación viral o en su potencial virulento. No obstante, algunas de las propiedades biológicas estudiadas sugieren que las cepas de dengue virus-3 son menos virulentas que las cepas cubanas de dengue virus-2 de 1997.
Palabras clave: dengue, cepas, propiedades biológicas, severidad.
ABSTRACT
Introduction: during dengue epidemics in Cuba, an increase in clinical severity with the epidemics progression in time, particularly in secondary infections, have been frequently observed. It is considered that this increase could be related with genetic changes in the circulating virus.
Objective: to study some biological attributes related to strains isolated at different points of time during the dengue epidemic occurred in Havana city, 2001-2002.
Methods: nine DENV-3 strains were studied. Cytopathogenic effect, viral growth in C6/36 HT and Vero cell lines, viral plaque sizes, temperature sensitivity, neurovirulence in newborn mice and pH influence in the binding of the virus and the cell as well as in the multiplication medium were evaluated.
Results: DENV-3 strains were more cytopathogenic in Vero Cells. However, higher titres were obtained in C6/36 HT cells. All the strains showed reduction of viral titres and plaque size with temperature increasing and low neurovirulence. Basic pH favoured virus-cell binding whereas acid pH was only permissive for some strains isolated at the end of the epidemic. On the other hand, at pH 6.5-7, the viral multiplication medium favoured the growth of strains isolated at the beginning of the epidemic whereas the growth of those isolated at the endof the epidemic was noticeable at pH 7-8.
Conclusions: this study proved the phenotypical changes among strains isolated at different points of time in the epidemic. They might be related to differences in viral fitness or in virulent potential. Nevertheless, some of the studied biological properties suggest that dengue virus-3 strains are less virulent than the Cuban dengue virus 2 strains isolated in 1997.
Key words: dengue, strain, biological attributes, severity.
INTRODUCCIÓN
En las Américas, los esfuerzos para erradicar el Aedes aegypti en las décadas de 1940 y 1950 restringieron las epidemias de dengue principalmente a la cuenca del Caribe. Cuba escapó a las infecciones por un genotipo de dengue virus (DENV) serotipo 3 endémico en Asia desde las décadas de 1960 y 1970.1 Tras la reintroducción en 1994 del serotipo 3 en Centroamérica, hubo una expansión notable al Caribe y Sudamérica, este virus aún se mantiene circulando de forma endémica.2-4 La presencia de un genotipo nuevo, luego de casi 2 décadas sin la circulación del DENV-3, se ha asociado con un incremento de los casos de dengue hemorrágico en nuestra región.5-8 Numerosos autores han confirmado que las cepas de DENV-3 aisladas desde 1994 hasta la fecha, pertenecen al genotipo III, el cual agrupaba originalmente, cepas aisladas durante epidemias severas ocurridas durante los años ochenta en la India y Sri Lanka. Los estudios filogenéticos demuestran el origen asiático de la cepa de DENV-3 introducida en 1994, constatándose una evolución in situ de esta cepa en las Américas.5-8
A pesar del fuerte programa de control y erradicación de Aedes aegypti que Cuba ha desarrollado durante 3 décadas, el país no ha estado exento de la introducción de los virus del dengue; aunque la vigilancia activa y las acciones de control establecidas han permitido detectar tempranamente y eliminar los brotes para evitar la endemicidad.9,10
Luego del control de la epidemia de Santiago de Cuba en 1997, no se reportaron más casos de dengue en la isla hasta 2000. Aunque desde 1994, se había demostrado la entrada del DENV-3 en Latinoamérica, no es hasta 2000 que se reportan los primeros casos de DENV-3 en Cuba.10 En este período se comprobó un pequeño brote que afectó 3 áreas de salud en La Habana, con un total de 138 casos, todos de fiebre dengue (FD), donde además se aisló el DENV-4.10
Sin bien, el brote de 2000 fue eliminado de inmediato, meses más tarde se reportó en la misma ciudad un brote de mayor envergadura, en el cual se identificó al DENV-3 como agente etiológico. Se extendió rápidamente a toda la ciudad de La Habana con el reporte de 12 889 casos confirmados de la enfermedad, con 78 casos de FHD/SCD y 3 fallecidos, todos adultos.11 Además, se produjeron algunas transmisiones locales en 5 provincias del país para un total de 14 443 casos confirmados, 81 casos de FHD/SCD y 3 fallecidos. En este período se desarrolló una estrategia de lucha antivectorial, lo cual posibilitó que la epidemia pudiera controlarse en 8 meses y no se extendiera al resto del país.11
Tanto en la epidemia de 1981 como en la de 1997, la enfermedad severa se observó en individuos con evidencias serológicas de infección por DENV-1 en 1977. En ambas epidemias, la proporción de casos de FHD/casos de FD, así como la proporción de fallecidos/casos de FHD, se incrementó de modo notable con la progresión de estas.12,13 Ese fenómeno también se observó durante la epidemia de DENV-3 del período 2001-2002, constatándose un incremento en su severidad con la progresión en el tiempo (Guzmán MG, Instituto de Medicina Tropical !Pedro Kourí", 2002, datos no publicados). Teniendo en cuenta estos elementos; así como los hallazgos en la caracterización molecular y biológica de cepas de DENV-2 de la epidemia de 1997, se consideró de interés estudiar algunas propiedades biológicas de cepas de DENV-3 aisladas en diferentes momentos de la epidemia ocurrida en el período 2001-2002.
MÉTODOS
Cepas virales: para el estudio se utilizaron 9 cepas de DENV-3 aisladas durante la epidemia de La Habana, 2001-2002, en diferentes momentos (inicio, medio y final). Todos los aislamientos virales fueron obtenidos a partir de suero de pacientes en fase aguda, con excepción de la cepa 580/01 aislada a partir de una muestra de tejido (bazo) obtenida durante la autopsia de un caso fatal. Las cepas utilizadas se multiplicaron en la línea celular C6/36 HT (Aedes albopictus, cepa celular del clono C6/36, ATCC CRL 1660). Una vez realizada la cosecha viral se prepararon lotes de todas las cepas, que fueron conservados a -70 °C y se les determinó el título viral (tabla 1).
Estudio del efecto citopático (ECP) en las líneas celulares C6/36 HT y Vero: se emplearon tubos plásticos sembrados con células de mosquito C6/36 HT (Aedes albopistus) y Vero (riñón de mono verde africano, ATCC CCL-81) que presentaban monocapa confluente. Se inocularon con las cepas en estudio a una multiplicidad de infección (m) de 0.01. Luego de 1 h de adsorción a 33 °C en el caso de las células C6/36 HT y a 37 °C para las células Vero, se eliminó el inóculo, se lavó la monocapa con el medio correspondiente (medio esencial mínimo Eagle, MEM, para los cultivos de C6/36 HT y medio 199 para los de Vero) y, finalmente, se les adicionó 1 mL de medio de mantenimiento que contenía los medios indicados antes y suero de ternera fetal inactivado 2 %. Los tubos de C6/36 HT se incubaron durante 6 d a 33 °C; tiempo durante el cual se observó a diario el ECP. En el caso de los tubos sembrados con la línea celular Vero el período de incubación fue de 9 d a 37 °C.
Cinética de crecimiento viral en las líneas celulares C6/36 HT y Vero: se emplearon tubos plásticos sembrados con células C6/36 HT y Vero con monocapa celular confluente. Se inocularon con las cepas en estudio a m= 0,01. Luego de 1 h de adsorción a 33 °C en el caso de C6/36 HT y a 37 °C para Vero se eliminó el inóculo, se lavó la monocapa con 1 mL del medio correspondiente y por último se les adicionó 1 mL de medio de mantenimiento. Se tomó a diario del primero al sexto día, un tubo de cada una de las cepas inoculadas en la línea celular C6/36 HT y del primero al noveno día en el caso de la línea celular Vero, se recogió el sobrenadante y se conservó a - 70 ºC. A continuación, este sobrenadante se descongeló para ser titulado por triplicado según el método descrito antes, a partir de diluciones seriadas de base 10 (10-1-10-4). Los resultados se graficaron enfrentando la media de los logaritmos de los títulos virales (log10 UFP/mL) contra los días de cosecha, que representan además la desviación estándar.
Titulación viral: para la titulación viral se siguió el método descrito por Álvarez y otros, utilizando células BHK-21 (riñón de hámster recién nacido, clono 13, ATCC CCL 10).14
Sensibilidad a la temperatura y tamaño de placas: los títulos virales se establecieron a 37 y 39 °C, se determinó el porcentaje de reducción del título viral con el aumento de la temperatura para cada cepa viral.14 Las placas virales fueron evaluadas según su tamaño. Se consideraron pequeñas todas aquellas con un diámetro menor o igual que 1 mm y grandes aquellas con un diámetro mayor que 1 mm. También se tuvo en cuenta la forma de las placas obtenidas clasificándolas en redondeadas, estrelladas o heterogéneas en el caso que una misma cepa viral tuviera placas redondeadas y estrelladas.
Determinación de la neurovirulencia en ratones lactantes: las cepas en estudio fueron diluidas en medio 199 e inoculadas por vía intracerebral en ratones lactantes de 24 h de nacidos, a razón de 20 UFP/animal; se inocularon 2 familias de 8 a 10 animales cada una, para cada cepa. Los ratones se observaron a diario durante 14 d en busca de cualquier signo de enfermedad o muerte. Las muertes ocurridas en las primeras 24 a 48 h no fueron consideradas a los efectos del estudio. Se determinó el porcentaje de supervivencia en los días posinfección para cada cepa viral.
Influencia del pH en la multiplicación del virus: todas las cepas incluidas en el estudio se inocularon en células BHK-21 utilizando diluciones seriadas en base 10 (10-1-10-4), aplicándose el método descrito para la titulación.14 Las cepas se titularon en 4 ensayos independientes, se evaluó la influencia de los valores de pH 6,5; 7; 7,5 y 8. El valor deseado se obtuvo ajustando el pH del medio de cultivo con bicarbonato de sodio 1 %.
Influencia del pH del medio de inoculación en la unión del virus a la célula: tubos plásticos (Greiner) sembrados con células C6/36 HT con monocapa celular confluente se inocularon con las cepas en estudio a m= 0,01. Al medio utilizado en la dilución de las cepas se le ajustó el pH con bicarbonato de sodio a pH 6; 6,5; 7 y 7,5. Luego de 1 h de adsorción a 33 °C se eliminó el inóculo, se lavó la monocapa con 1 mL de medio de mantenimiento sin suero fetal bovino inactivado (SFBI) y por último se les adicionó 1 mL de medio de mantenimiento. Los tubos se incubaron durante 2 d a 33 °C. Se tomó un tubo de cada una de las cepas inoculadas a 24 y 48 h, se recogió el sobrenadante y se conservó a - 70 ºC. A continuación, este sobrenadante fue descongelado para ser titulado por triplicado según el método descrito por Álvarez y otros.14
Análisis estadístico. Los resultados de los experimentos para determinar la influencia del pH en la multiplicación de los DENV así como en la unión de este a la célula fueron analizados mediante el programa Statistica; en el primer caso se hizo un Anova/Manova mientras que en el segundo caso se realizó una prueba de Student.
RESULTADOS
ECP en células C6/36 HT: el ECP observado consistió en la formación de sincitios redondeados, con bordes bien definidos y alta refringencia, que degeneraban hacia la desaparición de la estructura de la monocapa. Las cepas aisladas entre el primero y el tercer mes de la epidemia tuvieron un desarrollo más lento del ECP, mientras que las cepas del cuarto al séptimo mes mostraron una intensidad mayor del ECP desde más temprano. La formación de sincitios en estas últimas, fue apreciable en 50 % de la monocapa celular, a partir del cuarto día posinoculación, mientras que las cepas de los primeros meses de la epidemia, mostraban solo 25 % de la monocapa afectada. De manera general, se observa la presencia de 2 patrones de comportamiento. Las cepas del inicio pudieron ser agrupadas por mostrar un ECP de progresión lenta, por su parte, las cepas del final muestran una progresión más rápida.
ECP en células Vero: las cepas 492/01 y 21/02 mostraron ECP a partir del cuarto día post-inoculación, el resto muestra este ECP a partir del quinto día, con excepción de la cepa 73/01 aislada en el primer mes de la epidemia, en la que comienza el ECP a partir del sexto día. Más de la mitad de las cepas mostraron 50 % de destrucción de la monocapa al séptimo día posinoculación y al noveno día de observación la totalidad de los cultivos inoculados alcanza 50 % de ECP.
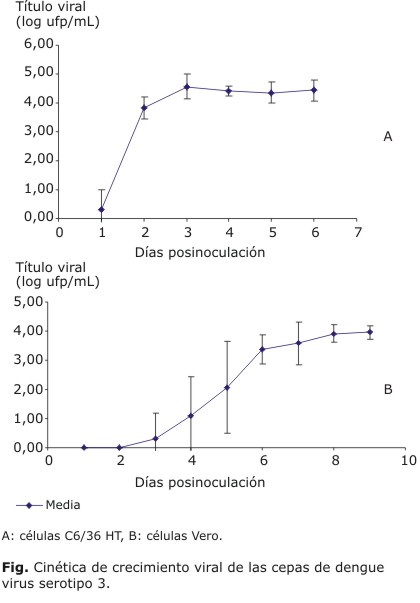
Cinética de crecimiento en células C6/36 HT y Vero: en la figura A, se muestra la curva obtenida a partir de la media y la desviación estándar del título infectivo de las cepas de DENV-3 según el día de cosecha. El título infectivo de las cepas aumentó con los días de cosecha hasta el tercer día, a partir de entonces se mantiene prácticamente similar para todas las cepas como muestra la curva de crecimiento. Los resultados de este experimento muestran un comportamiento homogéneo durante el crecimiento de las cepas de DENV-3 en células C6/36 HT. En la figura B, puede observarse que los títulos infectivos de las cepas multiplicadas en células Vero resultaron menores que los observados en la cinética desarrollada en la línea celular C6/36 HT. Solo la cepa 580/01 aislada de un fallecido por FHD elevó su título a partir del tercer día, el resto comenzó a multiplicarse entre el cuarto y el quinto día de la cosecha, la mayor parte de ellas en el quinto día. El título máximo se observó de forma general al octavo día para todas las cepas. La curva de crecimiento obtenida con las medias de los títulos virales de todas las cepas por días de cosecha, muestra una elevada desviación estándar hasta el sexto día, lo cual indica gran variabilidad entre estas en cuanto a su multiplicación en células Vero, en contraposición con la homogeneidad observada en la cinética desarrollada en células C6/36 HT.
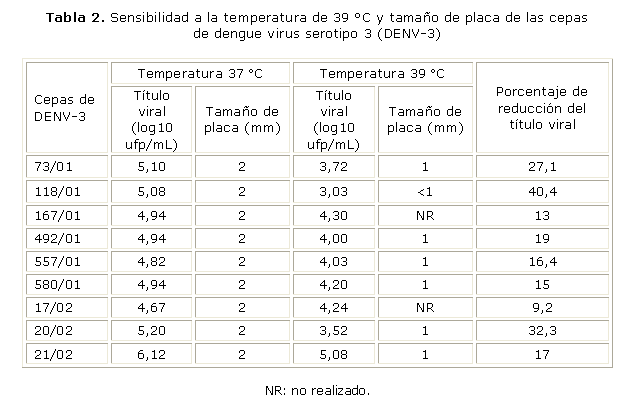
Sensibilidad a la temperatura: se observó una reducción del título viral que osciló entre 9 y 27 %, con excepción de las cepas 118/01 y 20/02, que mostraron reducciones de 40,4 y 32,3 %, respectivamente (tabla 2). Las placas observadas a 37 °C fueron de 2 mm y se apreció una reducción de la mitad de su diámetro al aumentar la temperatura. Las cepas 167/01 y 17/02 tuvieron dificultad para el desarrollo de las placas a 39 °C, las cuales no se observaron de manera nítida, por lo que no se pudo realizar su medición.
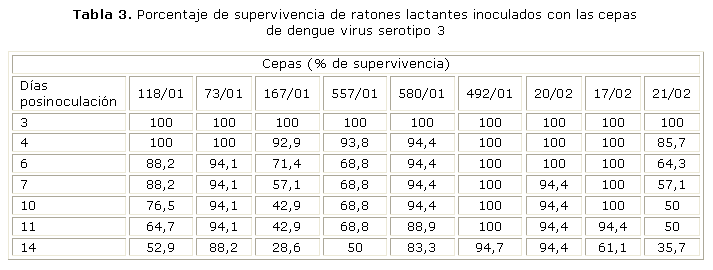
Neurovirulencia en ratones lactantes: las cepas de DENV-3 resultaron poco neurovirulentas. Pudo observarse que las familias de ratones inoculados con las cepas 492/01 y 20/02 mostraron el mayor porcentaje de supervivencia mientras que las cepas 167/01 y 21/02 fueron las más neurovirulentas. Esta observación no tiene una posible asociación ni con el cuadro clínico ni con el momento de aislamiento, porque puede observarse un comportamiento similar en las cepas del inicio y final, siendo todas aisladas de casos de FD (tabla 3).
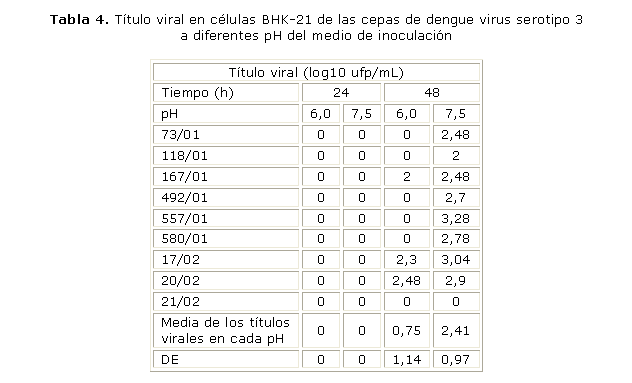
Influencia del pH del medio de inoculación en la unión del virus a la célula: en la tabla 4 puede observarse que ninguna de las cepas de DENV-3 mostró título a las 24 h posinoculación, a ninguno de los valores de pH estudiados. A las 48 h se observaron diferencias muy significativas entre los títulos infectivos obtenidos a los diferentes pH (p= 0,004). Los mejores títulos se alcanzaron a pH 7,5 y los más altos se encontraron para las cepas del final de la epidemia. Al incrementarse el valor del pH aumentó el número de cepas en las que se detectó título viral y solo las cepas del final fueron capaces de multiplicarse a un pH de unión ligeramente ácido (6,0). El título de las cepas del inicio de la epidemia solo se hizo notable cuando el pH de unión resultó algo alcalino (7,5).
Influencia del pH del medio en la multiplicación del virus: se evidenció que las cepas del inicio de la epidemia mostraron títulos virales ligeramente más superiores a pH 6,5-7, mientras que las del final de la epidemia mostraban títulos virales superiores a pH 7-8. No obstante, las diferencias encontradas no resultaron significativas (p= 0,7668).
DISCUSIÓN
Durante 3 epidemias de dengue ocurridas en Cuba (1981, 1997 y 2001), se ha observado un incremento notable de la severidad con el transcurso de estas, el cual se ha notificado mes por mes, apreciándose el aumento de la proporción de casos graves así como de fallecidos hacia el final de la epidemia. Ese fenómeno es relevante, si se tiene en cuenta que durante esta última fase es ostensible la efectividad de las medidas para controlar el vector, los médicos están conscientes de la situación epidemiológica, por lo que existe una política de ingreso temprano y un procedimiento clínico adecuado de los pacientes y, por lo tanto, se observa una disminución del número de casos. Dado que este incremento de severidad es reiterativo, se ha planteado que pudiese estar relacionado con cambios genéticos en el virus circulante que propicien una mayor adecuación de este hacia el final de las epidemias, puesto que no sería plausible plantear que los individuos más susceptibles se acumulan justo en esa etapa.
Los resultados obtenidos al estudiar las propiedades biológicas del grupo de cepas de DENV-3 de la epidemia cubana ocurrida en el período 2001-2002, indican que existe una variabilidad en el comportamiento de los virus aislados de diferentes momentos de una misma epidemia. Los estudios de Bielefeldt-Ohmann y otros15 muestran que el nivel de virus que se une a las células varía ampliamente al comparar diferentes líneas celulares y de manera notable entre cepas virales, incluso del mismo serotipo.
Asimismo, pudo comprobarse la existencia de diferencias entre las cepas de DENV-3 de este estudio y las cepas de DENV-2 de la epidemia de Santiago de Cuba, 1997 estudiadas previamente.16 Las cepas de DENV-2 resultaron más citopatógénicas en células C6/36 HT que las de DENV-3, sin embargo, estas últimas fueron más citopatogénicas en células Vero que las de DENV-2. Por otra parte, se encontró una concordancia entre los experimentos de estudio del ECP y la cinética de crecimiento viral. De esta forma las cepas menos citopatogénicas mostraron un crecimiento viral más tardío. En el caso de las cepas de DENV-2 caracterizadas por producir un mayor ECP en células C6/36 HT, se observó una caída de los títulos virales hacia el final de la cinética de crecimiento. Esta observación no pudo realizarse para DENV-3 porque la aparición más tardía del ECP, así como su poca intensidad, favoreció el tiempo de vida de las células durante la multiplicación viral. De esta manera los títulos se mantuvieron prácticamente constantes desde el tercer día posinoculación hasta el último día de la cinética; de haberse extendido el tiempo de la cinética de crecimiento, hubiese sido posible observar una disminución del título viral con el deterioro celular. Sin embargo, el hecho de que para las cepas de DENV-2 con la misma duración de la cinética se haya observado una caída del título infectivo entre el quinto y sexto día de cosecha, apoya el resultado obtenido en el estudio del ECP, donde se observa que las cepas de DENV-3 son menos citopatogénicas que las de DENV-2 en células C6/36 HT.
En este estudio se han encontrado algunos patrones de comportamiento que dividen las cepas de inicio y final en 2 grupos, como es la tendencia de las cepas del final de las epidemias a ser más citopatogénicas y que el pH ácido o básico favorezca la unión a la célula de un grupo de cepas y otras no. Además, se encontraron comportamientos atípicos para algunas cepas en particular que pudieran tener implicaciones en la patogénesis.
Si se analiza la sensibilidad a la temperatura para las cepas de DENV-3, en todas se reduce tanto el tamaño de las placas como el título viral al aumentar la temperatura a 39 °C. No obstante, pudo observarse que algunas cepas son mucho más sensibles a la temperatura que el resto, como es el caso de la 118/01 (del inicio de la epidemia), en la que se reduce el título viral en 40 % y las cepas 167/01 y 17/02 que presentaron dificultad para el desarrollo de placas a 39 °C.
Otro de los marcadores fenotípicos de virulencia de las cepas es la neurovirulencia en ratones lactantes; en este sentido, al estudiar las cepas de DENV-3 se encontró que esta era baja, con un comportamiento homogéneo. Todo lo anterior podría estar relacionado con el menor potencial virulento de las cepas de DENV-3, eso concuerda con que la proporción de casos de FHD/SCD respecto al total de casos, que fue menor para la epidemia de DENV-3 (0.56) que para la epidemia de DENV-2 (6,8).9,11
Con relación al incremento de severidad observado durante la epidemia de La Habana, 2001-2002, sería de gran interés estudiar la secuencia nucleotídica de cepas de diferentes momentos de la epidemia y correlacionar estos datos con los resultados del presente trabajo en cuanto a cambios fenotípicos de las cepas del final de la epidemia. Los hallazgos al estudiar las propiedades biológicas de cepas de DENV-3 muestran que de la misma forma que ocurrió para DENV-2 el virus sufre cambios con la progresión de la epidemia.16 Es de señalar que el estudio de la secuencia nucleotídica de las cepas de DENV-2 estudiadas previamente reveló la existencia de mutaciones en los genes no estructurales frente a una conservación total de los genes estructurales, así como de las regiones no codificantes.17,18 En particular el gen de la envoltura de 20 cepas de diferentes momentos de la epidemia de 1997, no tuvo variación alguna;19 sin embargo, la secuencia del gen de la envoltura de 2 de las cepas de DENV-3 analizadas en este estudio, reveló diferencias nucleotídicas no sinónimas.8 Es relevante señalar que una de las cepas correspondía a un caso fatal (580/01) y la otra a un caso de FD (21/02). Este trabajo constituye un acercamiento inicial en la caracterización de los virus aislados progresivamente en epidemias cubanas de dengue; su continuidad, sin duda, aportará nuevas evidencias sobre el papel del virus en la patogénesis de esta enfermedad.
REFERENCIAS BIBLIOGRÁFICAS
1. Halstead SB. Dengue in the Americas and Southeast Asia: do they differ? Rev Panam Salud Publica. 2006;20(6):407-15.
2. Guzmán MG. Dengue type 3 infection-Nicaragua and Panama October-November, 1994. MMWR. 1995;44:21-4.
3. Guzmán MG, Rosario D, Mune M, Álvarez M, Rodríguez R, Kourí G. Genetic relatedness of the dengue 3 virus isolated in the outbreak of dengue hemorrhagic fever in Nicaragua, 1994. Rev Cubana Med Trop. 1996;48(2):114-7.
4. Guzmán MG, Vázquez S, Martínez E, Álvarez M, Rodríguez R, Kourí G, et al. Dengue in Nicaragua, 1994: reintroduction of serotype 3 in the Americas. Bol Oficina Sanit Panam. 1996;121(2):102-10.
5. Messer WB, Gubler DJ, Harris E, Sivananthan K, de Silva AM. Emergence and global spread of a dengue serotype 3, subtype III virus. Emerg Infect Dis. 2003;9(7):800-9.
6. Uzcategui NY, Comach G, Camacho D, Salcedo M, Cabello de Quintana M, Jiménez M, et al. Molecular epidemiology of dengue virus type 3 in Venezuela. J Gen Virol. 2003;84(Pt 6):1569-75.
7. Lanciotti RS, Lewis JG, Gubler DJ, Trent DW. Molecular evolution and epidemiology of dengue-3 viruses. J Gen Virol. 1994;75( Pt 1):65-75.
8. Rodríguez-Roche R, Álvarez M, Holmes EC, Bernardo L, Kourí G, Gould EA, et al. Dengue virus type 3, Cuba, 2000-2002. Emerg Infect Dis. 2005;11(5):773-4.
9. Kourí G, Guzmán MG, Valdes L, Carbonel I, del Rosario D, Vázquez S, et al. Reemergence of dengue in Cuba: a 1997 epidemic in Santiago de Cuba. Emerg Infect Dis. 1998;4(1):89-92.
10. Guzmán MG, Kourí G. Dengue: an update. Lancet Infect Dis. 2002;2(1):33-42.
11. Pelaez O, Guzmán MG, Kourí G, Perez R, San Martin JL, Vázquez S, et al. Dengue 3 epidemic, Havana, 2001. Emerg Infect Dis. 2004;10(4):719-22.
12. Guzmán MG, Kourí G, Valdes L, Bravo J, Vázquez S, Halstead SB. Enhanced severity of secondary dengue-2 infections: death rates in 1981 and 1997 Cuban outbreaks. Rev Panam Salud Publica. 2002;11(4):223-7.
13. Guzmán MG, Kourí G, Halstead SB. Do escape mutants explain rapid increases in dengue case-fatality rates within epidemics? Lancet. 2000;355(9218):1902-3.
14. Álvarez M, Rodríguez-Roche R, Bernardo L, Morier L, Guzmán G. Improved Dengue virus plaque formation on BHK21 and LLCMK2 cells: Evaluation of some factors. Dengue Bulletin; 2005. p. 29.
15. Bielefeldt-Ohmann H, Meyer M, Fitzpatrick DR, Mackenzie JS. Dengue virus binding to human leukocyte cell lines: receptor usage differs between cell types and virus strains. Virus Res. 2001;73(1):81-9.
16. Rodríguez-Roche R, López Matilla L, Álvarez Vera M, Morier Diaz L, Guzmán Tirado MG. Propiedades biológicas de cepas de virus dengue serotipo 2 aisladas durante la epidemia en Santiago de Cuba, 1997. Rev Cubana Med Trop. 2011;63(3):211-9.
17. Rodríguez-Roche R, Sanchez L, Burgher Y, Rosario D, Álvarez M, Kourí G, et al. Virus Role During Intraepidemic Increase in Dengue Disease Severity. Vector Borne Zoonotic Dis. 2011;11(7):675-81.
18. Rodríguez-Roche R, Álvarez M, Gritsun T, Halstead S, Kourí G, Gould EA, et al. Virus evolution during a severe dengue epidemic in Cuba, 1997. Virology. 2005;334(2):154-9.
19. Rodríguez-Roche R, Álvarez M, Gritsun T, Rosario D, Halstead S, Kourí G, et al. Dengue virus type 2 in Cuba, 1997: conservation of E gene sequence in isolates obtained at different times during the epidemic. Arch Virol. 2005;150(3):415-25.
Recibido: 2 de febrero de 2012.
Aprobado: 16 de febrero de 2012.
Rosmari Rodríguez-Roche. Departamento de Virología, Instituto de Medicina Tropical "Pedro Kourí". Autopista Novia del Mediodía, Km 6 ½, Lisa. La Habana, Cuba. AP 601 Marianao 13. CP17 100. Fax: 53-7-2046051 Correos electrónicos: rosmari@ipk.sld.cu; liudmila@cim.sld.cu; mayling@ipk.sld.cu; lupe@ipk.sld.cu


{kind=link}
{kind=link}
{kind=link}
{kind=link}