










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Medicina Tropical
versión On-line ISSN 1561-3054
Rev Cubana Med Trop vol.67 no.2 Ciudad de la Habana mayo.-ago. 2015
ARTÍCULO ORIGINAL
Caracterización bioquímica y biológica del veneno de la serpiente Neotropical Macagua (Bothrops Colombiensis) de la región de Barlovento, estado Miranda, Venezuela
Biological and biochemical characterization of the Neotropical Macagua (Bothrops colombiensis) snake venom from Barlovento region, Miranda state, Venezuela
Dra. Elda E. Sánchez,I Dra. María E. Girón,II Dra. Belsy Guerrero,III Dr. Néstor L. Uzcátegui,II Dr. Alexis Rodríguez-AcostaII
I National Natural Toxins Research Center, Texas A&M University-Kingsville. USA.
II Laboratorio de Inmunoquímica y Ultraestructura Instituto Anatómico de la Universidad Central de Venezuela. Caracas.
III Laboratorio de Fisiopatología, Centro de Medicina Experimental, Instituto Venezolano de Investigaciones Científicas, Caracas 1020A.
RESUMEN
Introducción: el veneno de B. colombiensis no es solamente un elemento tóxico; en su composición existen múltiples componentes, que tienen un gran potencial terapéutico, principalmente en el tratamiento de patologías de la trombosis y la coagulación.
Objetivos: estudiar una mezcla de venenos de Bothrops colombiensis de una ubicacion geográfica de Venezuela, a fin de hacer un barrido de sus actividades hemostáticas, que permitirá posteriormente purificar y caracterizar moléculas con actividad antitrombótica y anticoagulantes, entre otras, con potencial terapéutico.
Métodos: el veneno a estudiar, es una mezcla de ellos obtenidos de serpientes provenientes de la Región de Barlovento, estado Miranda, Venezuela. Se caracterizó bioquímicamente por cromatografias de exclusión molecular, cromatografía de fase reversa C18 y por electroforesis a través de SDS–PAGE; y biológicamente por medio de actividades relacionadas con la hemostasia. Se analizaron los perfiles en relación a las actividades fibrinolítica, proteolítica sobre polvo azul y cadena β de insulina, procoagulante, hemorrágica y letal.
Resultados: la actividad hemorrágica, definida como la Dosis Hemorrágica Mínima fue de 8,7 mg/kg. La letalidad, definida como la Dosis Letal cincuenta fue 8,7 mg/kg. El veneno presentó actividad procoagulante y fibrinolítica. Las fracciones mostraron actividad fibrinolítica y proteolítica sobre polvo azul de ocultamiento y sobre la cadena β de insulina.
Conclusiones: las características biológicas de los componentes de este veneno le confieren un enorme potencial terapéutico, ya que contiene una alta actividad fibrinolítica y anticoagulante. Estos compuestos una vez purificados y caracterizados podrían explorarse como coadyuvantes en procesos trombolíticos, dado que disuelven coágulos de fibrina y degradan fibrinógeno, evitando episodios de retrombosis.
Palabras clave: actividad fibrinolítica; Bothrops colombiensis; hemostasia; hemorragia; veneno de serpientes.
ABSTRACT
Introduction: this paper is a screening of multiple toxic activities, of which some will be potentially useful for the management of coagulation pathologies.
Objetives: a pool of Bothrops colombiensis venoms from a specific geographical location was studied, in order to carry out a hemostatic activities screening, allowing then to purify and characterise molecules with antithrombotic and anticoagulant activity, among others, which could have therapeutic potential.
Methods: the venom was chromatographically by molecular exclusion and reverse phase C18 and SDS -PAGE characterized; its hemostatic activity was also established. Snakes were from the region of Barlovento, Miranda state, Venezuela. Profiles of fibrinolytic, proteolytic, procoagulant, hemorrhagic and lethal activities were analyzed. Hemorrhagic activity was 8.7 mg/kg. The LD50 was 8.7 mg/kg. The venom showed strongly procoagulant activity. Both, crude venom as fractions showed high fibrinolytic activity. The majority of the eluted fractions showed significant proteolytic activity in azure blue powder and on β chain of insulin.
Conclusions: the biological characteristics of the components of this venom confer enormous therapeutic potential because they contain a high fibrinolytic and anticoagulant activity. Most of these proteinases, once purified and characterized, could be explored as thrombolytic agents given that dissolves fibrin clots or prevent their formation.
Keywords: fibrinolytic activity; Bothrops colombiensis; hemostasis; hemorrhage; snake venom.
INTRODUCCIÓN
El veneno de serpientes del género Bothrops es una mezcla de componentes bioactivos de enzimas proteolíticas, péptidos y proteínas no enzimáticas que interactúan con múltiples componentes del sistema hemostático y la matriz extracelular.1-3 El género Bothrops ocasiona la gran mayoría de las mordeduras de serpiente en América Central y América del Sur y es responsable del 85 % de los envenenamientos por serpientes en Venezuela.4 Los envenenamientos se caracterizan por efectos locales, tales como edemas, hemorragias y necrosis, lo que puede llevar a daños permanentes por discapacidad de los miembros afectados. Tambien son generalmente graves las manifestaciones sistémicas, tales como hemorragia, coagulopatías de consumo, insuficiencia renal aguda y choque.5,6
Bothrops colombiensis es una serpiente muy agresiva y altamente venenosa, que causa accidentes graves, siendo la segunda serpiente más grande de Venezuela y que puede secretar cantidades considerables de veneno.7 En este, se han descrito toxinas variadas, incluyendo proteasas tales como serino y metaloproteinasas, fosfolipasa A2 y L-aminoacido-oxidasas.8 Se han purificado y secuenciado algunas de ellas, por ejemplo fibrinogenasas, que pueden actuar como serino o metaloproteinasas, las cuales muestran una variedad de actividades fisiopatológicas relacionadas con la hemostasis.1 Dentro de las metaloproteinasas se incluyen enzimas responsables de la escisión de las proteínas del tejido, tales como laminina, nidogen, fibronectina, colágeno tipo IV y proteoglicanos presentes en la membrana basal endotelial.9,10
Pero el veneno no es solamente un elemento tóxico; en su composición existen múltiples componentes, que tienen un gran potencial terapéutico, principalmente en el tratamiento de patologías de la trombosis y la coagulación. En el presente trabajo, se hace un barrido de múltiples actividades tóxicas y/o algunas potencialmente útiles para el manejo de patologías de la de la hemostasis, centrándose en la descripción de fracciones del veneno de B. colombiensis, con actividades procoagulante, hemorrágica, proteolítica, fibrinolítica y su acción sobre la función plaquetaria, analizando sus propiedades fisicoquímicas y biológicas, por lo tanto, permitirá tener información acerca de nuevas moléculas a caracterizarse en un futuro próximo, las cuales probablemente servirán para tratamientos antitrombóticos y suplementarios, así como la mejora de inmunógenos para producir mejores antídotos. Así, esta investigación preliminar, consentirá identificar muchas actividades importantes, para luego regresar e investigar minuciosamente, algunas de estas proteínas que tienen un futuro prometedor en la investigación biomédica.
MÉTODOS
Venenos
Se empleó una mezcla de venenos crudos, extraído de 15 ejemplares adultos de serpientes Macagua (Bothrops colombiensis) mantenidas en cautiverio en el serpentario del Instituto de Medicina Tropical (IMT) de la Universidad Central de Venezuela. Los venenos fueron extraídos, refrigerados a 4ºC, centrifugados para remover impurezas, liofilizados y congelados a –70 ºC hasta su uso.
Animales experimentales
Conejos
Para las pruebas de actividad hemorrágica en piel, se emplearon conejos blancos de la raza New Zealand (Oryctolagus cuniculus), con un peso promedio de 2 Kg, provenientes del Centro Nacional de Investigaciones de Toxinas Naturales, Texas A&M University-Kingville, Kingsville, Texas, U.S.A. Todas las especies empleadas en los modelos experimentales, fueron mantenidas bajo las condiciones establecidas por las normas de cuidado y uso de los animales de laboratorio, publicadas por el Instituto Nacional de Salud de los Estados Unidos de América.11
Ratones
Se utilizaron ratones machos (BALB/c) de 18-20 g de peso provenientes del Instituto Nacional de Higiene "Rafael Rangel", Caracas, Venezuela y del Centro Nacional de Investigaciones de Toxinas Naturales, Texas A&M University-Kingville, Kingsville, Texas, U.S.A. La colonia de ratones se mantuvo en cajas plásticas (Tecniplast, Italia), alojando 6 ratones en cada jaula, en una sala mantenida a 23° C en un ciclo de luz/oscuridad de 12/12-hr.
Declaración ética
Un personal profesional debidamente entrenado, estuvo a cargo de todos los eventos experimentales en el uso de animales vivos. Se utilizaron las regulaciones pertinentes, así como las pautas institucionales, según protocolos aprobados por el “National Natural Toxins Research Center”, de la A&M University-Kingsville, Texas, Estados Unidos y del Instituto de Anatomía de la Universidad Central de Venezuela y de normas obtenidas de las directrices para el cuidado y uso de animales de laboratorio, publicado por el “National Institute of Health, USA. 11
Ensayo de letalidad
La Dosis Letal cincuenta (DL50) se determinó mediante el método de Spearman & Karber 12, mediante la inyección intraperitoneal de un volumen de 0,2 mL de diluciones seriadas (desde 5.0 hasta 10 mg/kg) de veneno de B. colombiensis en ratones de 18-20 g de peso. Fueron utilizados cinco ratones por dosis, para un total de cinco dosis a ser probadas. Los animales se mantuvieron en observación por un período de 48 h. Los animales del grupo control fueron inyectados con una solución salina isotónica. Una vez finalizado los experimentos, los ratones fueron usados para la alimentación de las serpientes en cautiverio.
Curvas de Titulación Electroforéticas (TE)
Se hicieron curvas de titulación electroforética (TE) para determinar el contenido proteínico y el punto isoeléctrico (pI) de las proteínas del veneno. Las muestras se liofilizaron y se reconstituyeron en agua deionizada a 3 mg/mL. Se estableció un gradiente de pH de 3-9, en geles de IEF 3-9 (PhastGels de Amersham Biosciences, Piscataway, NJ, U.S.A.). Luego, el gel fue rotado 90˚ y 3 µL de la muestra fue aplicada en el centro del gel. Las proteínas separadas electroforeticamente fueron teñidas con plata según los manuales de Amersham Pharmacia PhastSystem™.
Fraccionamiento del veneno
Cromatografía de exclusión molecular
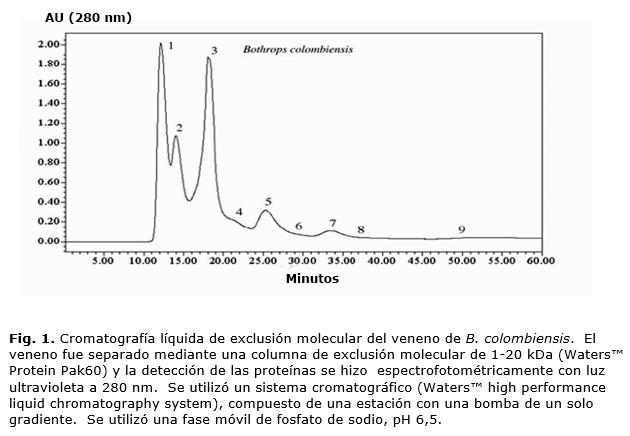
Para el fraccionamiento inicial del veneno se empleó una cromatografía de exclusión molecular sobre una columna Waters™Protein Pak 60 (Waters™, Corp., Milford, MA, U.S.A), el equilibrio de la columna y la elución de las proteínas se realizó con una fase móvil de fosfato de sodio 0,02 M, pH 6,5 a una velocidad de flujo de 0,5 mL/min. Para ello, se utilizó un sistema cromatográfico isocrático (Waters™ 5500 HPLC System), compuesto de una estación de trabajo con una bomba de gradientes, conformada por una bomba de un pistón. Dicho sistema emplea una válvula de inyección manual (Waters™U6K) y un detector ultravioleta para las proteínas (Waters™ 484 Tunable Detector).
Cromatografía de fase reversa C18
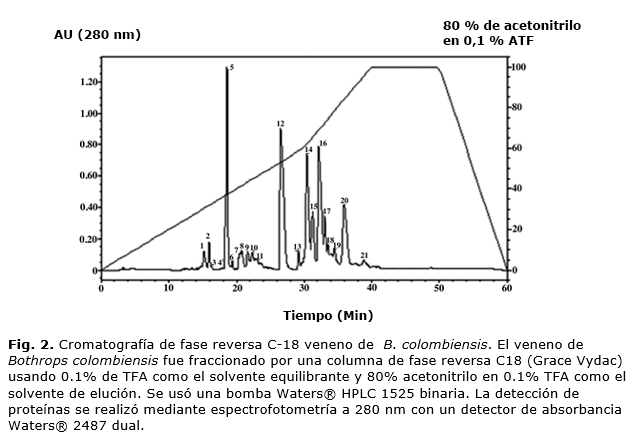
Las fracciones del veneno de B. colombiensis obtenidas por exclusión molecular, que contenían actividad sobre la agregación y retracción plaquetaria fueron recromatografiadas por una columna de fase reversa C-18 (Grace Vydac”C18, 50 x 4.5 mm, Hesperia, CA, U.S.A.). La columna se equilibró con una solución de ácido trifluoroacético (ATF) al 0,1 % y la elución se realizó con un gradiente de 0 a 80 % acetonitrilo en ATF 0,1 % durante 60 min, a una velocidad de flujo de 1,0 mL/min. Las proteínas se detectaron a 280 nm.
Electroforesis en geles de poliacrilamida
Se llevaron a cabo electroforesis en geles de dodecil sulfato de sodio y poliacrilamida (SDS-PAGE) (12 %), en condiciones reductoras, siguiendo el método de Laemmli.13 Se usaron marcadores de masa molecular (Bio-Rad) en paralelo y los geles se tiñeron con azul de Coomassie R-250. La masa molecular se determinó en un Multi-Analyst TM / PC versión 1.1 del software (Bio-Rad).
Diálisis y concentración de proteínas
Las fracciones del veneno de B. colombiensis obtenidas de las cromatografías fueron desalinizadas a través de una columna Pharmacia G25 HiTrap y concentradas en tubos de 15 micrones por liofilización al vacío a -40˚C (Freezone 6 de Labconco, MO, U.S.A.).
Análisis hemorrágico
Para determinar la actividad hemorrágica del veneno de B. colombiensis y de las fracciones, se utilizó la técnica modificada de Omori-Satoh et al. 14 Un total de 100 µL de cada fracción cromatográfica se inyectó intracutáneamente (i.c.) en la espalda del conejo. El animal se sacrificó a las 18 h y se extrajo la piel. La actividad hemorrágica específica fue determinada dividiendo el tamaño del punto hemorrágico (milímetros), por la cantidad de proteína inyectada (µg). La actividad hemorrágica se presenta como la Dosis Hemorrágica Mínima (DHM), que se define como la cantidad de veneno que causa un halo de hemorragia de 10 mm. Como control positivo se determinó la DMH del veneno crudo del C. atrox que fue de 2,5 µg.
Actividad Fibrinolítica
Para medir la actividad fibrinolítica, de las fracciones de B. colombiensis se utilizó el método modificado de Bajwa et al. 15 Se agregaron, a cada pozo de una placa de 24 pozos, 300 µL de una solución de fibrinógeno y 12 µL de una solución de trombina (2 UI/mL, en 0,025 M CaCl2); luego se agitó suavemente y se dejó formar el coágulo de fibrina a temperatura ambiente; después, la placa se incubó a 37˚C por 3 h. Para el ensayo se agregó a cada pozo 20 µL de cada fracción y se incubó a 37˚C por 15 h. Luego se agregó en cada pozo 700 µL de ácido tricloroacético al 10 %, para detener la reacción, decantándose después de 10 min. La actividad fibrinolítica específica se calculó dividiendo el área de la fibrina eliminada (milímetros) por la cantidad de muestra de proteína (µg) colocada en cada pozo.
Actividad proteolítica sobre Polvo de Azure
Se utilizó el método modificado de Rinderknecht et al.16 para probar la actividad proteolítica. Se mezclaron 8 mg de polvo azure con 2 mL de 0,02 M Tris-HCl, pH 8,0, luego se agregaron 100 µL de fracción cromatográfica, se incubó a 37˚C por 1 h y se agitó a intervalos de 5 min. Después de la incubación, la solución se centrifugó (centrífuga Beckman Avanti™ 3) a 420 x g por 5 min. El sobrenadante se transfirió a un vial y se midió la absorbancia a 595 nm. La actividad específica se calculó dividiendo la absorbancia por la cantidad de proteína usada (mg).
Análisis de gelatinasa
Se utilizó el método modificado de Huang y Pérez,17 para probar la actividad de las fracciones del veneno sobre gelatina. Veinte microlitros de cada fracción del veneno de B. colombiensis, se colocaron en una película de radiografía (Kodak X-OMAT) con capa de gelatina. La hidrólisis de la gelatina de la película de radiografía, se determinó lavando la película radiográfica con agua, después de una incubación a 37˚C por 4 h, en una incubadora con ambiente húmedo.
Actividad Proteolítica Sobre la Cadena β de Insulina
La actividad proteolítica de cada fracción cromatografica sobre la cadena β de insulina, se determinó por una electroforesis capilar (P/ACE 5500 de Beckman). Se incubaron 10 µL de la fracción del veneno (0,06 mg/mL) con 10 µL de una solución de cadena β de insulina (0,5 mg/mL) y 10 µL de 0,1M tampón borato de sodio pH 8,3 a temperatura ambiente por 1 h. Luego, la mezcla se separó durante 10 min a 20 kV, usando 0,1M tampón borato de sodio, pH 8,3, a través de un tubo capilar de 75 µm (800 x100 de abertura). Las proteínas fueron detectadas a 214 nm en un equipo UV de absorbancia de P/ACE.
Obtención de la muestra de sangre
La muestra de la sangre fue recogida usando un sistema que fluye por gravedad y causa poco trauma. Se recogieron dieciocho mililitros de sangre humana, en un tubo de 50 mL que contenía 2 mL de citrato de sodio al 1 %. Después de recogida la sangre, el tubo fue invertido suavemente dos veces para asegurar la citratación total de la sangre. Esta se dispensó en muestras de 2 mL antes de su uso.
Obtención de Plasma
Se obtuvo una muestra de la sangre humana de donantes sin alteraciones en la hemostasia, que fue anticoagulada con citrato de sodio al 3,8 %en una proporción 1:9. El plasma fresco fue obtenido por centrifugación a 2000 g, durante 15 min, a 4 ° C.
Actividad procoagulante
Para determinar la actividad coagulante del veneno de B. colombiensis, se siguió el método de Austen and Rhymes,18 utilizando como sustrato fibrinógeno bovino para evaluar actividad semejante a trombina o plasma fresco congelado para determinar actividad procoagulante (activadora de factores II, IX, o X). Brevemente, 100 µL de plasma citratado o solución 0,3 % de fibrinógeno en tampón Tris-HCl 0,05 M, pH 7,4 (tamón de coagulación) se incubaron por 3 min a 37 °C, después se agregaron 100 µL de tampón de coagulación y 100 µL de solución de trombina (0,5 a 15 UI/mL) y 100 µL de veneno crudo o fracción (100 µg/mL); se mezcló y se registró el tiempo de coagulación a 37 ° C. Como referencia, la solución de trombina se ajustó a 2,5 UI/mL, preparada con tampón de coagulación, que debería inducir la coagulación entre 18 y 22 seg.
La actividad similar a la trombina presente en el veneno crudo se reportó en UI/mL, al extrapolar los resultados en una curva de calibración hecha con un patrón de trombina.
Inhibición de la retracción plaquetaria
Se usó un ensayo de cristales activados (Kit-gbAC, Sienco®, Inc.) para la detección del coágulo, la formación de la fibrina y la función de la plaqueta en un Sonoclot® y Analizador de la Función de la Plaqueta (Sienco®, Inc. Wheat Ridge, CO, U.S.A.). Se incubó la sangre humana citratada a 37˚C, por al menos 5 min antes de su uso. La sangre incubada fue colocada en una cubeta y se le añadió 13 µL de CaCl2 0,25M, dándole una vuelta de 180º. Fueron agregados 13 µL de veneno de B. colombiensis o de las fracción cromatográfica, luego 300 µL de sangre humana citratada e incubada a 37˚C, evitando la formación de burbujas de aire. El ensamblaje principal del analizador se cerró 10 seg después de que se presionó la tecla de partida. La adquisición de datos fue analizada mediante un "Signature Viewer™" proporcionado por Sienco, Inc. en una computadora IMac.
Análisis de la inhibición de agregación plaquetaria
Se evaluó la agregación plaquetaria inducida por ADP usando un metodo turbidimétrico con un Chronolog™ (Havertown, PA, U.S.A.) Para determinar la inhibición de agregación plaquetaria inducida por las fracciones cromatográficas, 450 µL de sangre humana citratada fueron incubados a 37˚C por 5 min con 450 µL de solución salina 0,15M. Fueron incubados diez microlitros de la muestra del veneno de B. colombiensis, con la muestra de la sangre por 2 minutos. Se insertó un electrodo en la muestra de la sangre, y en un minuto y medio, se agregó 20 µL ADP 1mM a la muestra, para promover la agregación plaquetaria. Los datos fueron analizados mediante "Aggro/Link™ V.5.1.4" proporcionado por Chrono-Log™, Crop, en una computadora Dell.
RESULTADOS
Ensayo de letalidad
La DL50 calculada del veneno de B. colombiensis fue 8.7 mg/kg.
Cromatografía de exclusión molecular del veneno de B. colombiensis
El perfil cromatográfico obtenido del veneno de B. colombiensis muestra 9 fracciones (Fig. 1), de las cuales todas excepto F1 hidrolizaron la cadena β de la insulina. Las fracciones 1-4 mostraron actividad hemorrágica, las fracciones 2-5 y 7, 8 presentaron actividad fibrinolítica. Todas las fracciones del veneno tuvieron actividad procoagulante cuando fueron probadas en el analizador de Sonoclot. Las fracciones 1-6 inhibieron la agregacion plaquetaria inducida por ADP. La actividad fibrinolítica estaba presente en las fracciones 2-5 y 7-8. Las fracciones 5, 7 y 8 fueron re-diluidas para semicuantificar la actividad fibrinolítica, observándose que la fracción 5 fue la única que mantuvo actividad a una dilución > a 1/128.
Cromatografía de fase reversa C18
Cuando el veneno de B. colombiensis fue aplicado sobre la columna de fase reversa C-18 se obtuvieron 21 fracciones (Fig. 2). La agregación plaquetaria fue inhibida por las fracciones 3-6 en un 100 %, y por las fracciones 18 y 19 en un 69 y 85 %, respectivamente. La retracción plaquetaria fue inhibida por la fracción 14 en un 100 %. Las fracciones 14 y 15 presentaron actividad procoagulante. Ninguna de las fracciones presentó actividad proteolítica sobre polvo de azure.
Curva de titulación electroforético (TE)
En la curva de titulación electroforética de veneno crudo de B. colombiensis se observo un patrón de bandas que evidencia variaciones en la carga superficial de las proteínas, tituladas a diferentes pHs, lo que demostró que el veneno esta conformado por una mezcla de moléculas complejas, que contiene muchas proteínas ácidas (datos no mostrados). Aunque el TE se utiliza principalmente para determinar las condiciones óptimas para la separación de moléculas mediante cromatografía de intercambio iónico, también es utilizado para indicar su pureza.
Electroforesis en geles de poliacrilamida (PAGE-SDS)
El patrón electroforético en presencia de SDS, obtenido del veneno de B. colombiensis en condiciones reductoras sobre un gel al 12 %, mostró una gama de bandas entre ~ 16 y 22 kDas; entre ~ 27 y 32 kdas; alrededor de 38 kDa y entre 46 y 70 kDas.
Actividad Hemorrágica
La DMH del veneno crudo de B. colombiensis empleado en este estudio fue de 5,32 µg. Las fracciones 1, 2, 3 y 4 presentaron actividad hemorrágica (Tabla 1).
Actividad fibrinolítica
El veneno crudo de Bothrops colombiensis presentó una actividad fibrinolítica en placas de fibrina de 30,6 (mm²/µg). Esta actividad fibrinolitica se detecto en las fracciones 2-5 y 7-8 (Tabla 1).
Actividad proteolítica sobre polvo de azure
La actividad proteolítica sobre polvo de azure en el veneno crudo de B. colombiensis fue de 7.9 U/mg/proteína. Las fracciones 1-5 y 7 presentaron esta actividad (Tabla 1).
Determinación de la actividad proteolítica sobre la cadena β oxidada de la insulina de las fracciones de veneno de Bothrops colombiensis, utilizando electroforesis capilar
Todas las fracciones del veneno de B. colombiensis tenían actividad proteolítica y rompían la cadena β de insulina oxidada, con la excepción de la fracción 1, que no lo hizo (Tabla 1).
Actividad gelatinasa
El veneno crudo mostró una débil actividad gelatinolítica. Las fracciones de 1, 2 y 3 presentaron una ligera actividad proteolítica sobre gelatina (datos no mostrados). Las fracciones 4 a 12 no presentaron esta actividad.
Actividad procoagulante
La actividad procoagulante del veneno de B. colombiensis, en plasma fresco fue 11,0 ± 0,7 UI T/µg; sobre fibrinógeno humano purificado fue de 14,0 ± 0,9 UI T/µg.
Activación del coágulo
Todas las fracciones y el veneno crudo de B. colombiensis tenían actividad procoagulante, cuando fueron ensayadas en el analizador Sonoclot y activaron la coagulación antes de los 60 segundos (Tabla 1 y Fig. 3).
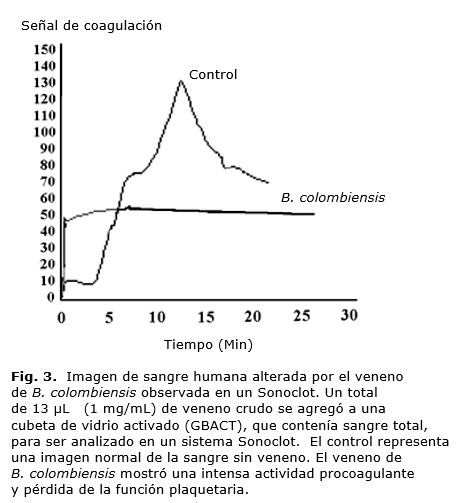
Inhibición de la agregación de plaquetas
Las fracciones 3, 4, 5, 6, 18 y 19 obtenidas por la cromatografía de fase reversa C18 inhibieron la agregación plaquetaria (Tabla 2).
DISCUSIÓN
La función principal de los venenos de serpiente es digestiva e interviene también en la captura de sus presas. En este último caso, provocando una muerte rápida, ocasionada por falla respiratoria o circulatoria, donde intervienen varias neurotoxinas, cardiotoxinas, enzimas con actividad sobre el sistema de la coagulación y fibrinolitico, y otras sustancias que actúan sinérgicamente, impidiendo que la presa escape.19,20 Los venenos contienen sustancias diseñadas para afectar procesos vitales, tales como la función de nervios y de músculos, la actividad cardíaca, la circulación sanguínea y la permeabilidad de las membranas.21, Los venenos de serpiente y otras secreciones tóxicas contienen una gran cantidad de sustancias farmacológicas, altamente activas y con un modo específico de acción. Puesto que tales compuestos son de gran valor en la investigación de procesos fisiológicos importantes, la investigación sobre venenos no es una actividad esotérica. La variación en la composición del veneno en el seno de una misma especie constituye un mecanismo biológico que dota a la serpiente de gran flexibilidad para adaptarse a ambientes cambiantes y poder así garantizar su supervivencia: de la adaptación a un ecosistema particular depende el éxito depredador de la especie.22
Las principales manifestaciones del envenenamiento bothrópico sistémico están representadas por las alteraciones en la hemostasia. Estas junto a las manifestaciones clínicas podrían ser provocadas por la variabilidad individual de los venenos. Las diferencias en la composición molecular y actividad farmacológica de los venenos de serpientes, se presentan no solo entre especies diferentes sino también dentro de la misma especie pero con edades o sexos diferentes.23
En esta investigación se evaluó las actividades fibrinolítica, coagulante, hemorrágica local y letal, además sobre las células plaquetarias, de los venenos de serpientes B. colombiensis provenientes de la localidad geográfica de Barlovento, estado Miranda, Venezuela.
En el envenenamiento bothrópico la coagulopatía observada se ha asociado a enzimas que actúan sobre el fibrinógeno con actividad similar a la trombina, adicional a enzimas procoagulantes capaces de generar trombina a partir de la activación de protrombina, factor X y/o factor IX. Esta actividad de trombina directa o que se genera, induce la activación de la hemostasia, en especial del sistema de la coagulación con consumo de plaquetas, fibrinógeno y de otros factores coagulantes, con generación de los productos de degradación del fibrinógeno/fibrina, en un proceso conocido como coagulopatía de consumo o Coagulación Intravascular Diseminada (CID), el cual es característico en pacientes víctimas de mordedura por vipéridos en Sur América.24-26
Hasta el presente, la actividad coagulante de B. colombiensis no ha sido caracterizada. Nuestros datos indican la presencia de una actividad procoagulante en este veneno. El método coagulante mostró que era más activo sobre fibrinógeno purificado, lo cual evidencia una elevada actividad semejante a trombina. Su acción, especialmente sobre plasma fresco como sustrato, evidencia que en este veneno existen activadores de protrombina y/ o del Factor X como ya ha sido reportado en otros venenos.27-30 En relación a las fracciones obtenidas por exclusión molecular, todas activaron la coagulación antes de los 60 seg, lo que las hace muy coagulantes. Los componentes presentes en la fracción de mayor masa molecular llevan a pensar en estudios futuros para aislar y caracterizar actividades procoagulantes las cuales deben estar asociadas con las coagulopatías evidentes en las manifestaciones clínicas presentes en los envenenamientos por B. colombiensis.4,6,30
Como se describió anteriormente, en los venenos de la familia Viperidae se han descrito enzimas fibrinolíticas que causan lisis de coágulos de manera directa o por acción de activadores del plasminógeno.31,32 Sin embargo, existen pocos estudios sobre la actividad fibrinolítica del veneno de B. colombiensis.2,3 Con miras a futuras aplicaciones clínicas de enzimas fibrinolíticas en terapia trombolítica, iniciamos el estudio con esta este veneno, el cual presenta una elevada actividad fibrinolítica con baja actividad procoagulante y hemorrágica. Esta actividad fibrinóltica se ha atribuido a metaloproteinasas 2,3 tal como se ha reportado en veneno de B.jararaca,25 actividad que debe estar asociada al cuadro hemorrágico inducido por la degradación de fibrinógeno/fibrina, así como de proteínas de matriz extracelular relacionadas con el proceso hemostático, cicatrización y modelaje de los vasos sanguíneos33 como laminina, fibronectina, entre otras.
La mayoría de las fracciones cromatográficas obtenidas del veneno de B. colombiensis por exclusión molecular mostraron actividad fibrinolítica (Tabla 1); entre estas las fracciones 2, 3 y 4 presentaron la más alta actividad procoagulante, hemorrágica, y fibrinolítica e inhibición de la agregación plaquetaria, lo que evidencia que en estas fracciónes deben estar concentradas la mayor parte de metaloproteasas asociadas a estas actividades.3
Las bandas entre 31 a 66 kDa, pueden estar asociadas con enzimas tipo serino como la Bothrombina30 y Bothrojarasina34 que a su vez se asocian con las actividades coagulantes y fibrinolítica presentes en estos venenos, así como a metaloproteinasas asociadas con actividades fibrinolítica y tipo hemorraginas como la jararagina.35 Las bandas entre 20 y 50 kDa, podrían estar asociadas a metaloproteasas del tipo P-II o P-III.2,5,10
La hemorragia es uno de los principales efectos locales del envenenamiento por viperidios y de gran significancia en la patología desarrollada en estos pacientes.7 Entre los componentes presentes en venenos de serpientes asociados a esta actividad se han reportado principalmente metaloproteinasas, entre estas las conocidas hemorraginas.3, 5
En cuanto a la presencia de metaloproteinasas en el veneno de B. colombiensis, nuestros resultados coinciden con los presentados en el análisis proteómico de este veneno realizado por Calvete et al. (2009).22 Ellos determinaron que el 42,3 % del contenido total de proteínas del veneno corresponde a metaloproteinasas dependientes de Zinc+,2 porcentaje repartido entre metaloproteinasas tipo PIII (11 %), que en nuestro caso correspondería a proteinasas hemorrágicas y probablemente a activadores de protrombina tipo A y B, concentradas en las fracciones de alto peso molecular obtenidas de la columna de exclusión molecular y metaloproteinasas tipo PI (30,8 %), que correspondería a metaloproteasas hemorrágicas y mayoritariamente no hemorrágicas probablemente α-fibrinogenasas, en las de menos peso molecular. En contraste, el porcentaje de serino proteinasas determinado por Calvete et al.,22 corresponde solo a un 1 % del contenido total de proteínas del veneno. Al compararlo con las actividades asociadas a serino proteinasas determinadas por estos autores (actividad similar a trombina, calicreína y t-PA), podría sugerir que se trate de proteínas pobremente representadas en términos cuantitativos, pero muy activas y por consiguiente, muy importantes en el cuadro clínico. Se ha demostrado que en el envenenamiento por ejemplares de la familia Viperidae proteínas similares a calicreína son responsables del dolor local, incrementos en la permeabilidad capilar, hipotensión sistémica y la contracción del músculo liso.36
La hemorragia local debida a lesiones en el endotelio vascular es un efecto clínico importante en el envenenamiento por B. colombiensis.2 Esta actividad hemorrágica es manifestada por equimosis y petequias locales con sangramiento en el sitio de la mordedura. La dosis mínima hemorrágica (DHM) evaluada en B. colombiensis mostró una DHM 5,32 µg, aproximadamente 47 % mayor a la reportada por los veneno de B. venezuelensis (2.48 µg) y C.atrox (2.5 µg).
El veneno del B. colombiensis fraccionado en una columna de fase reversa C18 se probaron en sus actividades proteolíticas y en su capacidad de inhibir la agregación y la retracción plaquetaria, solamente las fracciones que tenían actividad significativa o contenían una cantidad significativa de proteína, fueron reportadas. Debido a los solventes usados para fraccionar el veneno (80 % de acetonitrilo en 0,1 % TFA), ninguna de las fracciones presentaron actividad proteolítica. Las proteínas más grandes, tales como las metaloproteinasas (e.g. hemorraginas) son susceptibles a los solventes orgánicos, que pueden desnaturalizarlas de una manera irreversible, perdiendo su actividad. Las fracciones 3-6, 18 y 19 inhibieron la agregación plaquetaria. La fracción 14 también inhibió la retracción plaquetaria en el analizador Sonoclot™ en un 100 % (datos no mostrados) alterando el fibrinógeno, que originó la pérdida de la función plaquetaria, necesaria para la retracción del coágulo. La fracción 14 también causó inhibición de la agregación plaquetaria. En este caso, pensamos que el fibrinógeno se rompe en la cadena α, perdiendo su potencial de unirse a las integrinas, en la superficie de las plaquetas. Este fenómeno contribuye al falso positivo de la inhibición de la agregación plaquetaria, que ocurre en el Chrono-Log™aggregometer. La ruptura del fibrinógeno en la muestra sanguínea, inhibe a las plaquetas de unirse unas a las otras. Por otra parte, la ruptura del fibrinógeno convirtió el fibrinógeno a fibrina, lo que causó la presencia de coágulos en la muestra. Los coágulos, causaron una señal drástica de agregación o unión de las plaquetas en el coágulo, que dió lugar a una inhibición de la señal de la agregación. Vale decir, que es importante examinar visualmente la muestra sanguínea, después de que se haya analizado en un agregómetro. Los coágulos de sangre en la muestra son una buena indicación de que la proteína del veneno es enzimática y no una desintegrina.37
Se han purificado y caracterizado, una gran cantidad de toxinas de los venenos de serpiente. Estos estudios han demostrado, que cada veneno contiene toxinas que actúan sobre cientos de proteínas. Estas toxinas, sin embargo, pertenecen a un número muy pequeño de superfamilias de proteínas. Por ejemplo, un solo veneno de serpiente puede contener tanto como 15 isoformas de fosfolipasas A2.38-40 Estas comparten semejanzas notables en sus estructuras primarias, secundarias, y terciarias. Sin embargo, se diferencian ocasionalmente en sus blancos biológicos y por lo tanto en sus efectos farmacológicos. De manera similar, otras proteínas, como las proteínas no-enzimáticas, también se presentan en variadas isoformas41y se pueden clasificar en familias. Se han caracterizado hasta ahora más de 1000 proteínas no-enzimáticas, y estas toxinas se han agrupado en familias bien conocidas: 1) toxinas de tres-dedos, 2) inhibidores de serino-proteinasas, 3) lectinas, 4) sarafatoxinas, 5) factores del crecimiento del nervio, 6) péptidos atriales natriuteticos, 7) bradykininas reparadoras de péptidos, 8) helveprinas/CRISP, y 9) desintegrinas. Los miembros en cada familia de estas toxinas, tienen una composición molecular similar, pero exhiben funciones múltiples.
Muchos de los primeros esfuerzos de la investigación de venenos, fueron dirigidos hacia el aislamiento y la caracterización de cualquier proteína que se encontrara en abundancia o de los componentes más tóxicos. Con el advenimiento de técnicas más sofisticadas de purificación, ha habido estudios de los nuevos e interesantes componentes de proteína que se encuentran en cantidades más pequeñas. El trabajo hecho en esta publicación conducirá, en el futuro próximo, a las proteínas no-enzimáticas del veneno, las llamadas desintegrinas, donde ya hemos publicado una llamada Colombistatin,37 sobre más de 90 desintegrinas que han sido purificadas de los venenos de las familias Atractaspididae, Elapidae, Viperidae y Colubridae.42
Nuestros resultados destacan la importancia de complementar los estudios de proteómica de los venenos con la exhaustiva evaluación de las actividades biológicas de sus componentes.
En conclusión, las acciones tóxicas de los venenos de serpientes son versátiles ya que sus diferentes componentes tienen actividades que varían, pudiendo también ser más activas en relación con otras toxinas al actuar sinérgicamente incrementando en consecuencia sus acciones finales. La variabilidad de los venenos ocurre en diversos ámbitos incluyendo variaciones inter e intra especie.43,44 Adicionalmente, los componentes de los venenos pueden ser alterados por la localización geográfica y el hábitat de las serpientes.2,22,44
AGRADECIMIENTOS
El financiamiento para esta investigación fue proporcionado por las subvenciones del CDCH de la UCV (PG: 09-8760-2013/1. FONACIT (PG-2005000400), Instituto Venezolano de Investigaciones Científicas, Caracas, Venezuela, NCRR/BMRG, Viper Grant #s 8P40OD01960-10 y 3P40OD01096-10S1 (NNTRC, Texas A&M University-Kingsville, Dr. Sánchez) y la donación de Robert A. Welch Foundation Department # AC-0006 (TAMUK-Departamento de Química).Los autores declaran que no hay conflicto de interés con respecto a la publicación de este documento.
REFERENCIAS BIBLIOGRÁFICAS
1. Kamiguti AS, Cardoso JL. Haemostatic changes caused by the venoms of South American snakes. Toxicon 1989;27:955-63.
2. Girón ME, Salazar AM, Aguilar I, Pérez JC, Sánchez EE, Arocha-Piñango CL, et al. Hemorrhagic coagulant and fibrino(geno)lytic activities of crude venom and fractions from mapanare (Bothrops colombiensis) snakes. Comp Biochem Physiol C Toxicol Pharmacol. 2008;147:113-21.
3. Girón ME, Guerrero B, Salazar AM, Sánchez EE, Alvarez M, Rodríguez-Acosta A. Functional characterization of fibrinolytic metalloproteinases (colombienases) isolated from Bothrops colombiensis venom. Toxicon 2013;74:116-26.
4. Rengifo C, Rodríguez-Acosta A. Serpientes Veneno y Tratamiento Médico en Venezuela. Universidad Central de Venezuela, Caracas: Fondo Editorial de la Facultad de Medicina;2004.
5. Markland FS. Snake venoms and the hemostatic system. Toxicon 1998;36:1749-800.
6. Jorge MT, Ribeiro LA. Envenoming by the South American pit viper Bothrops neuwiedi Wagler. Ann Trop Med Parasitol. 2000;94:731-34.
7. Rodríguez-Acosta A, Uzcátegui W, Azuaje R, Aguilar I, Girón ME. A clinical and epidemiological analysis of accidental bites by snakes of the genus Bothrops in Venezuela. Rev Cubana Med Trop. 2000;52:90-4.
8. Loria GD, Rucavado A, Kamiguti AS, Theakston RD, Fox JW, Alape A, Gutiérrez JM. Characterization of 'basparin A' a prothrombin-activating metalloproteinase from the venom of the snake Bothrops asper that inhibits platelet aggregation and induces defibrination and thrombosis. Arch Biochem Biophys. 2003;418:13-24.
9. Kamiguti AS, Hay CR, Theakston RD, Zuzel M. Insights into the mechanism of haemorrhage caused by snake venom Metalloproteinases. Toxicon 1996;34:627-42.
10. Bjarnason JB, Fox JW. Hemorrhagic metalloproteinases from snake venoms. Pharmacol Ther. 1994;62:325-72.
11. NIH. Principles of Laboratory Animal Care. National Institute of Health of United States, Maryland. 1985.
12. Spearman-Kärber F. Alternative Methods of Analysis for Quantal Responses. In: Statistical Method in Biological Assay Finney DJ Charles Griffin & Co Ltd. Londres. 1978.
13. Laemmli U. Cleavage of structural proteins during the assembly of the head of bacteriophage T-4. Nature 1970;227:608-9.
14. Omori-Satoh T, Sadahiro S, Ohsaka A, Murata R. Purification and characterization of an antihemorrhagic factor in the serum of Trimeresurus flavoviridis a crotalid . Biochim Biophys Acta. 1972;285:414-26.
15. Bajwa SS, Markland FS, Russel FE. Fibrinolytic enzyme(s) in western diamondback rattlesnake (Crotalus atrox) venom. Toxicon 1980;18:185-90.
16. Rinderknecht H, Geokas MC, Silverman P, Haverback BJ. A new ultrasensitive method for the determination of proteolytic activity. Clin Chim Acta. 1968;21:197-03.
17. Huang S, Perez JC. Comparative study on hemorrhagic and proteolytic activities of snake venoms. Toxicon 1980;18:421-26.
18. Austen D, Rhymes I. A Laboratory Manual of blood coagulation. Oxford, London: Osney Mead, Blackwell Scientific Publications;1975.
19. Swenson S, Markland FS jr . Snake venom fibrin(ogen)olytic enzymes. Toxicon 2005;45:1021-39.
20. Jorge MT, Ribeiro LA. The epidemiology and clinical picture of an accidental bite by the South American rattlesnake (Crotalus durissus). Rev Inst Med Trop Sao Paulo. 1992;34:347-54.
21. Jorge da Silva N Jr, Aird SD. Prey specificity, comparative lethality and compositional differences of coral snake venoms. Comp Biochem Physiol C Toxicol Pharmacol. 2001;128:425-56.
22. Calvete JJ, Borges A, Segura A, Flores-Díaz M, Alape-Girón A, Gutiérrez JM, et al. Snake venomics and antivenomics of Bothrops colombiensis a medically important pitviper of the Bothrops atrox-asper complex endemic to Venezuela: Contributing to its taxonomy and snakebite management . J Proteomics 2009;72: 227-40.
23. Rael ED, Lieb CS, Maddux N, Varela-Ramirez A, Perez J. Hemorrhagic and Mojave toxins in the venoms of the offspring of two Mojave rattlesnakes (Crotalus scutulatus scutulatus). Comp Biochem Physiol B. 1993;106:595-600.
24. Salazar AM, Rodríguez-Acosta A, Girón ME, Aguilar I, Guerrero B. A comparative analysis of the clotting and fibrinolytic activities of the mapanare (Bothrops atrox) snake venom from different geographical areas in Venezuela. Thromb Res. 2007;120:95-04.
25. Sugiki M, Maruyama M, Yoshida E, Mihara H, Kamiguti AS, Theakston DG. Enhancement of plasma fibrinolysis in vitro by jararhagin the main haemorrhagic metalloproteinase in Bothrops jararaca venom. Toxicon 1995;33:1605-617.
26. Hoffmann H, Bon C. Blood coagulation induced by the venom of Bothrops atrox, identification purification and properties of two factor X activators. Biochemistry. 1987;26:780-87.
27. Zingali RB, Jandrot-Perrus M, Guillin M, Bonn C. Bothrojaracin a new thrombin inhibitor isolated from Bothrops jararaca venom: characterization and mechanism of thrombin inhibition. Biochemistry. 1993;32:10794-802.
28. Sánchez EF, Santos CI, Magalhaes A, Diniz CR, Figueiredo S, Gilroy J, et al. Isolation of a proteinase with plasminogen-activating activity from Lachesis muta muta (bushmaster) snake venom. Arch Biochem Biophys. 2000;378:131-41.
29. Bello CA, Hermogenes AL, Magalhaes A, Veiga SS, Gremski LH, Richardson M, Sánchez EF. Isolation and biochemical characterization of a fibrinolytic proteinase from Bothrops leucurus (white-tailed jararaca) snake venom. Biochimie. 2006;88:189-200.
30. Kini RM. Anticoagulant proteins from snake venoms: structure function and mechanism. Biochem J. 2006;397:377-87.
31. Hermógenes AL, Richardson M, Magalhaes A, Yarleque A, Rodríguez E, Sánchez EF. Interaction of a plasminogen activator proteinase LV-PA with human alpha2-macroglobulin. Toxicon. 2006;47:490-94.
32. Sánchez EF, Swenson S. Proteases from South American snake venoms affecting fibrinolysis. Curr Pharmac Analysis. 2007;3:147-57.
33. Jiménez N, Escalante T, Gutiérrez JM, Rucavado A. Skin pathology induced by snake venom metalloproteinase: acute damage revascularization and re-epithelization in a mouse ear model. Invest Dermatol. 2008;128:2421-428.
34. Zingali RB, Ferreira MS, Assafim M, Frattani FS, Monteiro RQ. Bothrojaracin, a Bothrops jararaca snake venom-derived (pro)thrombin inhibitor, as an anti-thrombotic molecule. Pathophysiol Haemost Thromb. 2005;34:160-63.
35. Moura-da-Silva AM, Baldo C. Jararhagin, a hemorrhagic snake venom metalloproteinase from Bothrops jararaca. Toxicon. 2012;60:280-89.
36. Yukelson LYa, L'vov VM, Shkinev AV, Sultanalieva N. The kallikrein, kininase and related peptides activities in central Asian snake venoms. Agents Actions Suppl. 1992;38:430-40.
37. Sánchez EE, Rodriguez-Acosta A, Palomar R, Lucena SE, Bashir S, Soto JG, et al. Colombistatin: a disintegrin isolated from the venom of the South American snake (Bothrops colombiensis) that effectively inhibits platelet aggregation and SK-Mel-28 cell adhesion . Arch Toxicol. 2009;83:271-79.
38. Ogawa T, Oda N, Nakashima K, Sasaki H, Hattori M, Sakaki Y, et al. Unusually high conservation of untranslated sequences in cDNAs for Trimeresurus flavoviridis phospholipase A2 isozymes. Proc Natl Acad Sci USA. 1992;89:8557-561.
39. Takasaki C, Suzuki J, Tamiya N. Purification and properties of several phospholipases A2 from the venom of Australian king brown snake (Pseudechis australis). Toxicon. 1990;28:319-27.
40. Braganca BM, Sambray YM. Multiple forms of cobra venom phospholipase A. Nature. 1967;216:1210-1.
41. McCleary RJ, Kini RM.Non-enzymatic proteins from snake venoms: a gold mine of pharmacological tools and drug leads. Toxicon. 2013 Feb;62:56-74
42. Mclane MA, Sanchez EE, Wong A,Paquette-Straub C, Perez JC. Disintegrins. Curr Drug Targets Cardiovasc Haematol Disord. 2004;4:327-55.
43. Aguilar I, Guerrero B, Salazar AM, Girón ME, Pérez JC, Sánchez EE, et al. Individual venom variability in the South American rattlesnake Crotalus durissus cumanensis. Toxicon. 2007;50:214-24.
44. Sasa M. Diet and snake venom evolution: can local selection alone explain intraspecific venom variation? Toxicon. 1999;37:249-52.
Dr. Alexis Rodríguez-Acosta. Correo electrónico: rodriguezacosta1946@yahoo.es


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}