










My SciELO
 Custom services
Custom servicesServices on Demand
Journal

Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Cubana de Medicina Tropical
Print version ISSN 0375-0760On-line version ISSN 1561-3054
Rev Cubana Med Trop vol.69 no.1 Ciudad de la Habana Jan.-Apr. 2017
ARTÍCULO ORIGINAL
Clustered distribution of virus infected cells in the liver of patients with chronic hepatitis C
Distribución agrupada de las células infectadas por el virus en el hígado de pacientes con hepatitis C crónica
Viviana Falcón,I Nelson Acosta-Rivero,II Sirenia González,III Emilio Acosta Medina,IV Rocio Garateix Suárez,I José A. Silva,I Liz Álvarez-Lajonchere,I Gillian Martínez-Donato,I Carmen M. Rosales,V Santiago Dueñas-Carrera,I Juan KouríIII
I Center for Genetic Engineering and Biotechnology (CIGB). Havana, Cuba.
II Centre for Protein Studies. Faculty of Biology. University of Havana. Havana, Cuba.
III CINVESTAV-IPN. México, DF, México.
IV Centre for Advanced Study in Cuba. Havana, Cuba.
V Latin American School of Medicine. Havana, Cuba.
ABSTRACT
Introduction: detection of hepatitis C virus RNA and proteins in the liver can not only contribute to the knowledge of the mechanisms of HCV replication and pathogenesis but also complement diagnostics and therapeutic studies.
Objective: to determine the presence of hepatitis C virus in the liver of patients with chronic hepatitis C that were immunized with a therapeutic vaccine candidate.
Methods: detection of hepatitis C virus RNA and proteins were studied by in situ hybridization and immunofluorescence assays in liver biopsies from 14 hepatitis C virus -infected individuals.
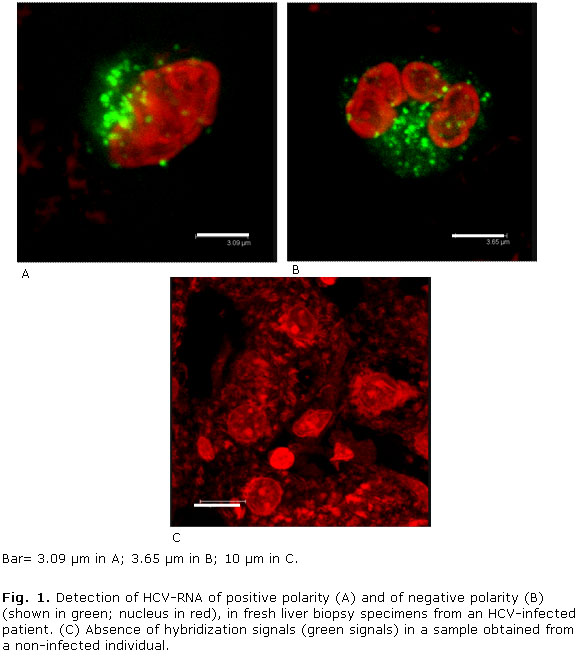
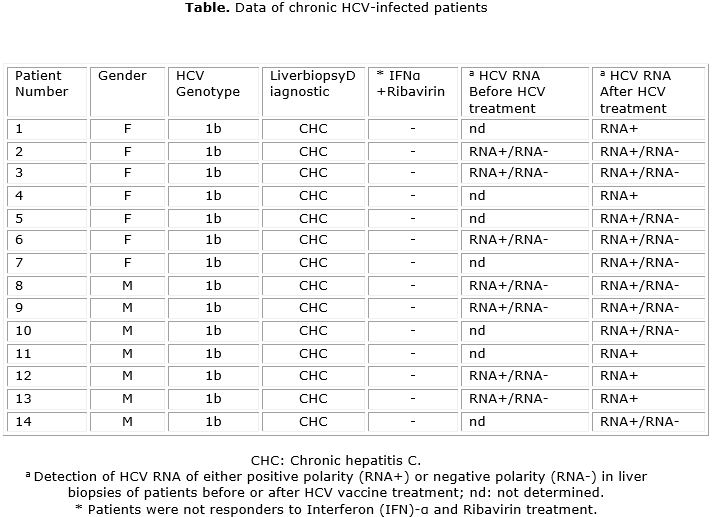
Results: hybridization signals for hepatitis C virus-RNA of both positive and negative polarity were detected in the cytoplasm of hepatocytes from most of the samples analyzed. However, the HCV structural antigens could not be detected in any of the samples. The hepatitis C virus RNA was observed on granular structures in the cytoplasm of hepatocytes. This staining pattern is similar to that described for stress granules and/or lipid droplets which are involved in hepatitis C virus replication. The proportion of cells showing positive reactions for hepatitis C virus RNA of negative polarity (which is a marker of ongoing viral replication) ranged from 4.47 % to 15.94 %.
Conclusions: results from this work suggest that hepatitis C virus infection occurs in groups of neighbouring hepatocytes that are distributed sporadically in the liver. This is consistent with the model of clustered spatial distribution of hepatitis C virus infected cells and cell to-cell spread of hepatitis C virus in the liver, and suggests that hepatitis C virus replication was constrained in these patients.
Keywords: HCV; viral replication; liver biopsies; in situ hybridization.
RESUMEN
Introducción: la detección de proteínas y ARN del virus de la hepatitis C en el hígado no sólo puede contribuir al esclarecimiento de los mecanismos de replicación y patogénesis del VHC, sino también a complementar los estudios diagnósticos y terapéuticos.
Objetivo: determinar la presencia de virus de hepatitis C en el hígado de pacientes con hepatitis C crónica que fueron inmunizados con un candidato vacunal terapéutico.
Métodos: se determinó la presencia de proteínas y ARN del virus de la hepatitis C mediante hibridación in situ e inmunofluorescencia en biopsias de hígado de 14 pacientes infectados con el virus.
Resultados: se detectaron señales de hibridación de ARN del virus de la hepatitis C de polaridad tanto positiva como negativa en el citoplasma de hepatocitos de la mayoría de las muestras analizadas. Sin embargo, los antígenos estructurales del VHC no pudieron detectarse en ninguna de las muestras. El ARN del virus de la hepatitis C se observó en estructuras granulares del citoplasma de hepatocitos. Ese patrón de tinción es similar al descrito para los gránulos de estrés y/o gotas de lípidos que participan en la replicación del virus de la hepatitis C. La proporción de células con reacciones positivas para ARN del virus de la hepatitis C de polaridad negativa (que es un marcador de la ocurrencia de replicación viral) osciló entre 4,47 % y 15,94 %.
Conclusiones: Los resultados del estudio sugieren que la infección por el virus de la hepatitis C tiene lugar en grupos de hepatocitos adyacentes que se distribuyen de forma esporádica en el hígado, lo que concuerda con el modelo de distribución espacial agrupada de las células infectadas con el virus de la hepatitis C y la propagación de célula a célula del virus de la hepatitis C en el hígado, y además sugiere que la replicación del virus era limitada en esos pacientes.
Palabras clave: VHC; replicación viral; biopsias de hígado; hibridación in situ.
INTRODUCTION
Hepatitis C virus (HCV) infection is a major health problem (130-170 million people are estimated to be chronically infected) and is considered as a leading cause of liver disease worldwide.1 Most of infected persons develop chronic disease (70-80 %) that can progress to cirrhosis, hepatocellular carcinoma (HCC) and liver failure.1 In addition, it is predicted that the prevalence of HCV-related cirrhosis and HCC will considerably increase in the next decade.2 With the recent development of directly acting antiviral agents (DAAs) for HCV, significant improvements in sustained virological response rates have been obtained for patients infected with HCV.3 However, as less than 5 % of the world's HCV-infected population are estimated to be aware that they are infected; most individuals with HCV infection will remain at risk for transmitting the virus to others.4 Thus, development of vaccines to prevent both persistent HCV infection and reinfection after cure of chronic hepatitis C with costly DAAs, is an area of real unmet clinical need.4-6
HCV was first identified in 1989 using molecular biology techniques from sera of experimentally infected chimpanzees.7 HCV is an enveloped positive-strand RNA virus classified as the type member of the genus Hepacivirus in the family Flaviviridae.8 The HCV genome is a 9.6 kb single-stranded positive sense RNA that contains a single open reading frame (ORF) encoding a polyprotein of 3010-3030 amino acids (aa) residues.9 The ORF is flanked by 5´- and 3´-terminal nontranslated regions (NTRs important for viral RNA translation and replication. Upon its synthesis, the HCV polyprotein is co- and post-translationally processed by cellular and viral proteases into at least 10 mature cleavage products, including the structural (core, E1 and E2) and the non-structural (NS) proteins (p7, NS2, NS3, NS4A, NS4B, NS5A and NS5B). Core, E1, E2, p7 and NS2 are primarily involved in HCV assembly while NS3 to NS5B are primarily involved in viral RNA replication.9
A major advance in HCV research occurred in 2005 with the discovery that the highly replicative HCV JFH1 strain (genotype 2a) was capable of producing infectious virions from select hepatoma cell lines in culture.10-12 Despite recent advances in the knowledge of HCV replication and morphogenesis in cell culture systems little is known about HCV life cycle and the mechanisms of viral pathogenesis in the liver of virus infected persons. Although detection of HCV components in the liver of infected patients have been hampered by the low levels and complexity of HCV morphogenesis, we have previously provided evidences suggestive of HCV replication and assembly in vivo. Thus, detection of HCV RNA and proteins in the liver can not only contribute to the knowledge of the mechanisms of HCV replication and pathogenesis but also complement diagnostics and therapeutic studies. The goal of this study was to determine the presence of HCV in the liver of patients with chronic hepatitis C that were immunized with a therapeutic vaccine candidate.
METHODS
PATIENTS AND SAMPLES
Patients infected with HCV genotype 1b have been previously described.13 They had a diagnosed chronic hepatitis by liver biopsy and had no other documented cause of liver disease. Patients were non responders to previous IFN-α plus ribavirin treatment. Liver needle biopsy samples were taken at the time of routine diagnostic biopsy from all patients before and after treatment with the therapeutic HCV vaccine candidate (CIGB-230) (composed of a recombinant HCV Core and a plasmid expressing HCV Core, E1 and E2) as shown before.13 Biopsies were immediately snap-frozen and stored at -80 °C until further processing. A liver needle biopsy sample was taken from a HCV-uninfected healthy donor liver for transplantation purpose as a negative control. All procedures were conducted in accordance with the national ethics guidelines and the Helsinki Declaration of 1975, as revised in 1983. Fresh frozen liver biopsies samples from 7 patients were analysed before HCV treatment and biopsies from 14 patients were investigated after HCV treatment (table).
ANTIBODIES AND PRIMERS
The following mouse monoclonal antibodies (mAbs) were used for immunodetection studies: anti-core SS-HepC.1 mAb recognizing aa 5 to 35 of core, anti-E1 SS-HepC.2mAb recognizing aa 190 to 219 of E1 and anti-E2 SS-HepC.3 mAb recognizing aa 643 to 671 of E2. These mAbs have been described elsewhere (Falcon, 2003). The following synthetic probes corresponding to the highly conserved 5' noncoding region of HCV have been shown before (14): To detect the HCV-RNA of positive strand the biotin-labelled antisense probe (HCV-1) (5'- biotin-GTTTATCCAAGAAAGGACCC-3', position 188-207) was used. To detect the HCV-RNA of negative strand the biotin-labelled sense probe (HCV-2) (5'-biotin-TTCACGCAGAAAGCGTCTAG-3', position 63-82) was used.14
IN SITU HYBRIDIZATION (ISH) ASSAY
Samples were immediately fixed with 4 % paraformaldehyde in PBS at4 ºC and then mounted on gelatine-coated glass slides and stored for 2 days at -200 ºC. The specificity of the ISH assay was confirmed in sample obtained from a HCV-uninfected individual as previously described.14
IMMUNOFLUORESCENCE STAINING
Samples were immediately fixed with 4 % paraformaldehyde in PBS at4 ºC and then mounted on gelatine-coated glass slides and stored for 2 days at -200 ºC. Later on, mounted samples were hydrated for 10 min in PBS and incubated with 0.2 % Triton X in PBS during 10 min. To block non-specific antibody reaction, best results were obtained by incubating the sections with 0.2 % bovine serum albumin (BSA) (free of IgG) (Sigma Chemical Co. St. Louis, Mo. USA), for 10 min at RT. After two washes in PBS-T, samples were incubated overnight at 4 ºC with either the anti-core mAb or the anti-E1 mAb or the anti-E2 mAb (dilutions 1:20 inPBS-T). Incubations were followed by washes with PBS-T. The second incubations were accomplished with FITC-conjugated anti-mouse IgG (dilutions 1:60 in PBS-T, Vector laboratories, Inc., Burlingame, CA., USA) for 1 hour at RT. After three washes with PBS-T the sections from all samples were counter stained with propidium iodide (dilution 1:1000, Vector laboratories, Inc. Burlingame CA., USA), followed by extensive washing in PBS-T. Immunostained samples were cover slipped in Vectashield mounting medium (Vector Laboratories, Inc. Burlingame, CA., USA), sealed with nail polish and viewed on a confocal laser scanning microscope. Negative controls were performed by substituting the primary antibodies with normal mouse serum.
LASER CONFOCAL SCANNING IMAGING AND SERIAL SECTION COLLECTION
Samples were viewed with a 60x (NA 1.4) objective on a Nikon microscope with attached laser confocal scanning system MRC 600 (BioRad, Watfod).Ten to twelve fields were imaged from each sample. Four to fifteen serial optical z-sections (0.2-0.5 µm thick) were collected from each observed field using the dual channel imaging with filter (554 nm) for propidium iodide excitation and the filter (494 nm) forthe fluorescein channel. Each series of confocal optical sections was scanned through a total of 25 µm. The resulting optical sections were fully projected onto two dimensional planes using the imaging processing system of the microscope (Camos package). The proportion of hepatocytes showing positive labelling was calculated using the ImageJ 1.38 software (Website: http://rsb.info.nih.gov/ij/). Association of treatment and labelling of HCV-RNA of negative strand was analysed with a 2 x 2 contingency table using the Fisher exact test (GraphPad Prismv5.0).Comparison between proportions of HCV RNA detection before and after vaccination was performed using the z-test (StatGraphics Centurion v15.1.02).
RESULTS
DETECTION OF HCV IN LIVER BIOPSY SAMPLES
All HCV infected patients analyzed before HCV treatment showed labelling for both positive and negative strands of the HCV genome that were localized in the cytoplasm of hepatocytes from liver biopsies (Fig. 1 A, B; Fig. 2 A, B and table). However, no hybridization signals were observed in a liver biopsy specimen from a non-infected individual (Fig. 1, C). Note that HCV RNA-specific signals localize to granules-like structures (Fig. 1 and 2). Detection of negative HCV-RNA strand has been related with ongoing viral replication. In this study, the proportion of cells showing positive reactions for HCV RNA of negative polarity ranged from 4.47 % to 15.94 % (mean ± SD 9.52 % ± 5.1). Nonetheless, reaction products suggestive of HCV antigens could not be detected on any of the samples (Not shown).

On the other hand, all HCV infected patients analyzed after HCV treatment showed hybridization labelling for positive strands of the HCV genome while the HCV RNA of negative polarity was detected in liver biopsies from 9 out 14 HCV-infected patients (64.28 %) (table). Nevertheless, there was neither association between detection of HCV RNA of negative polarity and HCV treatment (p= 0.1235 Fisher's exact test) nor difference between proportions of HCV RNA detection before and after vaccination (p= 0.07 Z test for comparison between proportions). Interestingly, hepatocytes containing such reaction products were found in groups of adjacent cells distributed sporadically or focally (Fig. 1 and 2).
DISCUSSION
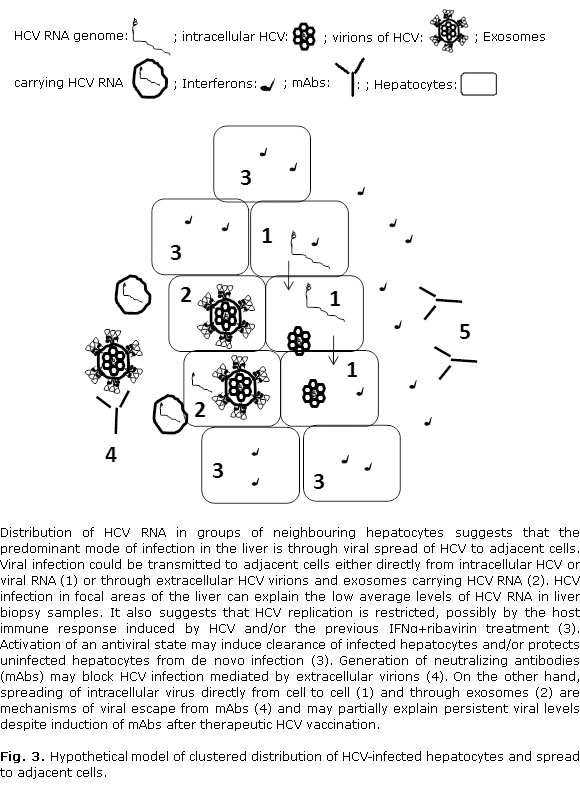
Detection of HCV RNA and proteins in the liver of virus infected individuals is essential to elucidate the mechanisms of HCV replication and pathogenesis at the cellular level. It is also important to complement diagnostics and therapy studies. HCV replication proceeds via synthesis of a complementary negative-strand RNA using the genome as a template and the subsequent synthesis of genomic positive strand RNA from this minus-strand RNA intermediate.15 Therefore, detection of negative HCV-RNA strand can be regarded as a marker of ongoing viral replication. We have previously shown that HCV-RNA of both positive and negative polarity can be detected in patients negative for viral RNA in serum, thus showing the relevance of this approach in therapeutic studies.14 The proportion of viral infected or replicating hepatocytes has been shown to range from 1 to 54 % depending on the serum HCV detection and viral load.14,16-18 Thus, the proportion of HCV-replicating hepatocytes has been estimated to be around 10 % in the liver of patients negative for serum HCV-RNA.14 This is similar to results from this study showing positive reactions for HCV RNA of negative polarity in 9.52 % of the hepatocytes analyzed, suggesting that HCV replication were restricted in these patients, possibly by the host immune response and/or the previous IFNa+ribavirin treatment (Fig. 3).
Hybridization signals for both positive and negative strands of HCV RNA were detected in all liver biopsies taken before treatment. On the other hand, after the therapeutic HCV vaccination, HCV RNAs of positive and negative polarity were labelled in 100 % and 64.28 % of individuals, respectively. These results are in agreement with previous quantitative analysis of the relative amount of the minus-strand of HCV-RNA describing that it can be from 10 to 100 times lower than that of the plus one in liver tissue of patients with chronic hepatitis.19 On the other hand, a diminished proportion of patients showing immunolabelling of the HCV RNA of negative polarity after vaccination might suggest a reduction in HCV replication rates but not in viral clearance because the HCV RNA of positive polarity was detected in all samples. In addition, these results are consistent with preceding investigation describing that HCV titers in sera were not affected by HCV vaccination.13 Moreover, no association was shown between HCV RNA labelling and HCV treatment. Nevertheless, one limitation of this study is the small number of patients, which increases the variance and may compromise the certainty of the findings.
Results from this work indicate that HCV infection and replication take place in groups of neighbouring hepatocytes that are distributed sporadically in the liver (Fig. 3). This is in accord with previous studies showing that detection of HCV RNA in hepatocytes occurs in focal areas of the liver.16-18 Based on the clustered spatial distribution of HCV infected cells, it has been proposed that the predominant mode of infection in the liver is by cell to-cell spread of HCV.16-18 Viral infection could be transmitted to adjacent cells either directly from intracellular HCV or viral RNA20,21 or through extracellular HCV virions and exosomes carrying HCV RNA (Fig. 3).22 Spreading of intracellular virus directly from cell to cell is a mechanism of viral escape from neutralizing antibodies (mAbs) and may partially explain persistent viral levels despite induction of mAbs after therapeutic HCV vaccination (Fig. 3).13
Focal distribution of infected hepatocytes can also explain the low average levels of HCV RNA in biopsy samples. Although previous reports have shown that activation of innate immune responses and the endogenous IFN system do not affect the proportion of infected hepatocytes and the serum viral levels,18 other studies have suggested that IFNβ expression correlates with viral but not interferon stimulated genes (ISGs) levels.17 Therefore, it has been proposed that activation of an antiviral state may induce clearance of infected hepatocytes and/or protects uninfected hepatocytes from de novo infection suggesting that HCV replication in hepatocytes may be transient.17 Interestingly, increased mRNA detection of the ISGIFI27 has been reported in both HCV-infected hepatocytes and adjacent cells of patients infected with HCV suggesting that the stimulus driving the ISG mRNA expression originated from HCV infected hepatocytes.18 In addition, HCV RNA and ISG mRNA levels have been shown to be comparable across all sites from an individual liver in end-stage liver disease.24 Nevertheless, HCV persist in the liver of most infected individuals and those exhibiting high levels of hepatic ISGs are less responsive to pegylated IFN-α and ribavirin treatment.23-26 HCV-infected patients studied in this work were non responders to IFN-α and ribavirin treatment. As a result, it is possible that the IFN-mediated innate immune response was activated in these patients contributing to control of viral infection in the liver (Fig. 3).
HCV RNA was detected on granular structures in the cytoplasm of hepatocytes which are similar to those described in the HCV cell culture system.27,28 HCV has been shown to replicate its genome in a cytoplasm microenvironment, called the membranous web ´MW´.29 On the other hand, components of the HCV replication complex have been detected in various cytoplasmic structures showing punctate spots or speckle-like arrangement.27,28 These subcellular organelles have been shown to be stress granules (SGs) and/or lipid droplets (LDs). Interestingly, at early stages of HCV infection host factors such as DEAD Box Helicase 3, X-Linked (DDX3X) and I-Kappa-B Kinase-Alpha (IKKα) and the HCV 3'unstranlated region (HCV 3'UTR) localize to SGs.27 HCV NS proteins also localize with SGs suggesting the involvement of SGs in HCV replication.30,31 In fact, several SGs proteins including GTPase-activating protein (SH3 domain)-binding protein 1 (G3BP1), TIA-1, TIAR, ataxin-2 (ATX2), and poly(A)-binding protein 1 (PABP1), have been shown to be required for HCV RNA replication, assembly and secretion.30-32 Then, when HCV core is present in sufficient levels on LDs, DDX3X and SGs may redistribute from endoplasmic reticulum (ER) to LDs contributing to the viral life cycle.27 Further studies will be necessary to elucidate the nature of the cytoplasmic granular structures containing HCV RNA that were observed in hepatocytes and their role in HCV viral life cycle and pathogenesis.
We could not detect the HCV structural proteins in the samples analysed. This is in contrast with our previous immunoelectron and immunofluorescence microscopy studies showing that core and envelope proteins localize mainly to ER membranes in hepatocytes of HCV-infected patients.14,33 Nonetheless, detection of HCV proteins in human liver has only been sporadically reported and involved samples with very high viral load. It has been suggested that the focal distribution of HCV infection, the low average levels of HCV RNA and the very low expression of viral proteins in the liver may hamper detection of HCV antigens.16,17,34
CONCLUSIONS
Hybridization signals using probes specific for HCV-RNA of both positive and negative polaritywere detected in the cytoplasm of hepatocytes from most of the samples analyzed. No association was shown between labelling of the negative strand of HCV RNA and the HCV treatment. In addition, HCV RNA was observed on granular structures which are similar to subcellular organelles described as stress granules and/or lipid droplets and that have been shown to be required for replication, assembly and secretion of HCV in the cell culture system. Nevertheless, reaction products suggestive of HCV structural antigens could not be detected in any of the samples. HCV RNA was shown in groups of neighbouring hepatocytes that were distributed sporadically in the liver. This is consistent with the model of clustered spatial distribution of HCV infected cells and cell to-cell spread of HCV in the liver. It also suggests that HCV replication were constrained in these patients, possibly by the host immune response.
BIBLIOGRAPHIC REFERENCES
1. Mohd Hanafiah K, Groeger J, Flaxman AD, Wiersma ST. Global epidemiology of hepatitis C virus infection: new estimates of age-specific antibody to HCV seroprevalence. Hepatology. 2013;57:1333-42.
2. Kanwal F, Hoang T, Kramer JR, Asch SM, Goetz MB, Zeringue A, et al. Increasing prevalence of HCC and cirrhosis in patients with chronic hepatitis C virus infection. Gastroenterology. 2011;140:1182-8.
3. Welzel TM, Dultz G, Zeuzem S. Interferon-free antiviral combination therapies without nucleosidic polymerase inhibitors. Journal of Hepatology. 2014;61:S98-S107.
4. Cox AL. Global control of hepatitis C virus. Science. 2015;349(6250):790-1.
5. Swadling L, Klenerman P, Barnes E. Ever closer to a prophylactic vaccine for HCV. Expert Opin Biol Ther. 2013;13(8):1109-24.
6. Walker CM, Grakoui A. Hepatitis C virus: why do we need a vaccine to prevent a curable persistent infection? Current Opinion in Immunology. 2015;35:137-43.
7. Choo QL, Kuo G, Weiner AJ, Overby LR, Bradley DW, Houghton M. Isolation of a cDNA clone derived from a blood-borne non-A, non-B viral hepatitis genome. Science. 1989;244(4902):359-62.
8. Simmonds P. The origin of hepatitis C virus. Curr Top Microbiol Immunol. 2013;369:1-15.
9. Moradpour D, Penin F. Hepatitis C virus proteins: from structure to function. Curr Top Microbiol Immunol. 2013;369:113-42.
10. Lindenbach BD, Evans MJ, Syder AJ, Wolk B, Tellinghuisen TL, Liu CC, et al. Complete replication of hepatitis C virus in cell culture. Science. 2005;309:623-6.
11. Wakita T, Pietschmann T, Kato T, Date T, Miyamoto M, Zhao Z, et al. Production of infectious hepatitis C virus in tissue culture from a cloned viral genome. Nat Med. 2005;11:791-6.
12. Zhong J, Gastaminza P, Cheng G, Kapadia S, Kato T, Burton DR, et al. Robust hepatitis C virus infection in vitro. Proc Natl Acad Sci U S A. 2005;102:9294-9.
13. Alvarez-Lajonchere L, Shoukry NH, Grá B, Amador-Cañizares Y, Helle F, Bédard N, et al. Immunogenicity of CIGB-230, a therapeutic DNA vaccine preparation, in HCV chronically infected individuals in a Phase I clinical trial. J Viral Hepat. 2009;16:156-67.
14. Falcón V, Acosta-Rivero N, Shibayama M, Luna-Munoz J, Miranda-Sanchez M, de la Rosa MC, et al. Evidences of hepatitis C virus replication in hepatocytes and peripheral blood monocuclear cells from patients negative for viral RNA in serum. Am J Infect Dis. 2005;1(1):34-42.
15. Lohmann V. Hepatitis C Virus RNA Replication. Curr Top Microbiol Immunol. 2013;369:167-98.
16. Liang Y, Shilagard T, Xiao SY, Snyder N, Lau D, Cicalese L, et al. Visualizing hepatitis C virus infections in human liver by two-photon microscopy. Gastroenterology. 2009;137:1448-58.
17. Stiffler JD, Nguyen M, Sohn JA, Liu C, Kaplan D, Seeger C. Focal distribution of hepatitis C virus RNA in infected livers. PLoS One. 2009;4:e6661.
18. Wieland S, Makowska Z, Campana B, Calabrese D, Dill MT, Chung J, et al. Simultaneous detection of hepatitis C virus and interferon stimulated gene expression in infected human liver. Hepatology. 2014;59(6):2121-30.
19. Fong TL, Shindo M, Feinstone SM, Hoofnagle JH, Di Bisceglie AM. Detection of replicative intermediates of hepatitis C viral RNA in liver and serum of patients with chronic hepatitis C. J Clin Invest. 1991;88:1058-60.
20. Fafi-Kremer S, Fofana I, Soulier E, Carolla P, Meuleman P, Leroux-Roels G, et al. Viral entry and escape from antibody-mediated neutralization influence hepatitis C virus reinfection in liver transplantation. J Exp Med. 2010;207(9):2019-31.
21. Timpe JM, Stamataki Z, Jennings A, Hu K, Farquhar MJ, Harris HJ, et al. Hepatitis C virus cell-cell transmission in hepatoma cells in the presence of neutralizing antibodies. Hepatol. 2008;47(1):17-24.
22. Liu Z, Zhang X, Yu Q, He JJ. Exosome-associated hepatitis C virus in cell cultures and patient plasma. Biochem Biophys Res Commun. 2014;455:218-22.
23. Pfeffer LM, Li K, Fleckenstein JF, Marion TN, Diament J, Yang CH, et al. An interferon response gene signature is associated with the therapeutic response of hepatitis C patients. PLoS ONE2014;9(8): e104202.
24. Asselah T, Bieche I, Narguet S, Sabbagh A, Laurendeau I, Ripault MP, et al. Liver gene expression signature to predict response to pegylated interferon plus ribavirin combination therapy in patients with chronic hepatitis C. Gut. 2008;57(4):516-24.
25. Chen L, Borozan I, Feld J, Sun J, Tannis LL, Coltescu C, et al. Hepatic gene expression discriminates responders and nonresponders in treatment of chronic hepatitis C viral infection. Gastroenterol. 2005;198:1437-44.
26. Sarasin-Filipowicz M, Oakeley EJ, Duong FHT, Christen V, Terracciano L, Filipowicz W, et al. Interferon signaling and treatment outcome in chronic hepatitis C. PNAS. 2008;105:7034-9.
27. Pène V, Li Q, Sodroski C, Hsu C-S, Liang TJ. Dynamic Interaction of Stress Granule, DDX3X and IKK-α Mediates Multiple Functions in Hepatitis C Virus Infection. J Virol. 2015;89(10):5462-77.
28. Romero-Brey I, Merz A, Chiramel A, Lee J-Y, Chlanda P, Haselman U, et al. Three-dimensional architecture and biogenesis of membrane structures associated with hepatitis C virus replication. PLoS Pathog. 2012;8(12):e1003056.
29. Gosert R, Egger D, Lohmann V, Bartenschlager R, Blum HE, Bienz K, et al. Identification of the hepatitis C virus RNA replication complex in Huh-7 cells harboring subgenomic replicons. J Virol. 2003;77:5487-92.
30. Ariumi Y, Kuroki M, Kushima Y, Osugi K, Hijikata M, Maki M, et al. Hepatitis C virus hijacks P-body and stress granule components 515 around lipid droplets. Journal of Virology. 2011;85:6882-92.
31. Yi Z, Pan T, Wu X, Song W, Wang S, Xu Y, et al. Hepatitis C virus co-opts Ras-GTPase-activating protein-binding 687 protein 1 for its genome replication. Journal of Virology. 2011;85:6996-7004.
32. Garaigorta U, Heim MH, Boyd B, Wieland S, Chisari FV. Hepatitis C virus (HCV) induces formation of stress granules whose proteins regulate 525 HCV RNA replication and virus assembly and egress. Journal of Virology. 2012;86:11043-56.
33. Falcón V, Acosta-Rivero N, Chinea G, Gavilondo J, de la Rosa MC, Menendez I, et al. Ultrastructural evidences of HCV infection in hepatocytes of chronically HCV-infected patients. Biochem Biophys Res Commun. 2003;305(4):1085-90.
34. Lau DT, Fish PM, Sinha M, Owen DM, Lemon SM, Gale M Jr. Interferon regulatory factor-3 activation, hepatic interferon-stimulated gene expression, and immune cell infiltration in hepatitis C virus patients. Hepatol. 2008;47:799-809.
Recibido: 24 de junio de 2016.
Aceptado: 10 de abril de 2017.
Viviana Falcón. Center for Genetic Engineering and Biotechnology (CIGB). P.O. Box 6162, Havana 10600, Cuba. E-mail: viviana.falcon@cigb.edu.cu


{kind=link}
{kind=link}
{kind=link}