










My SciELO
 Custom services
Custom servicesServices on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Cubana de Medicina Tropical
Print version ISSN 0375-0760On-line version ISSN 1561-3054
Rev Cubana Med Trop vol.69 no.2 Ciudad de la Habana May.-Aug. 2017
ARTÍCULO DE REVISIÓN
Péptidos antimicrobianos: potencialidades terapéuticas
Antimicrobial peptides: their therapeutic potential
Melaine González García,I Javier San Juan Galán,II Fidel Ernesto Morales VicenteIII Anselmo J. Otero GonzálezI
I Centro de Estudios de Proteínas. Facultad de Biología. Universidad de La Habana. La Habana, Cuba.
II Laboratorio Nacional de Referencia de Micología. Instituto de Medicina Tropical "Pedro Kourí". La Habana, Cuba.
III Departamento de Química General. Facultad de Química. Universidad de La Habana. La Habana, Cuba.
RESUMEN
El aumento en la incidencia de las enfermedades infecciosas en los últimos años se ha favorecido por diferentes causas. Entre estas se destacan las inmunodeficiencias adquiridas (sida, trasplantes de órganos, quimioterapia oncológica), la migración de personas que trae consigo la posibilidad de importar enfermedades hacia poblaciones susceptibles, así como el excesivo empleo de antibióticos. Debido a esta situación se ha incrementado la búsqueda de nuevos candidatos terapéuticos para el desarrollo de terapias más efectivas. En este sentido los péptidos antimicrobianos constituyen una opción promisoria, pues presentan un amplio espectro de actividad frente a varios microorganismos patógenos. Además, se encuentran ampliamente distribuidos en la naturaleza, desde organismos unicelulares hasta mamíferos. Algunos péptidos antimicrobianos ya están siendo evaluados en estudios clínicos aunque muchos de ellos no han tenido resultados favorables in vivo debido a su poca estabilidad metabólica y toxicidad, entre otros. Con el fin de optimizar estas propiedades de los péptidos antimicrobianos se han trazado diferentes estrategias como la modificación química de su estructura y la conjugación con nanopartículas magnéticas. Es por eso que este artículo tiene el objetivo de revisar las potenciales aplicaciones terapéuticas de estas moléculas, teniendo en cuenta la información publicada al respecto en MedLine, Web of Science y Scopus en los últimos años.
Palabras clave: péptidos antimicrobianos; susceptibilidad; agentes infecciosos; antibióticos; nanoconjugados; análogos químicos.
ABSTRACT
The growing incidence of infectious disease in recent years may be attributed to several causes, among them acquired immunodeficiencies (AIDS, organ transplant, oncological chemotherapy), human migration and the consequent import of diseases into susceptible populations, and the excessive use of antibiotics. This situation has fostered the search for new therapeutic candidates for the development of more effective treatments. Antimicrobial peptides are a promising alternative in this respect, due to their broad spectrum of activity against several pathogenic microorganisms. Moreover, they are widely distributed in nature, from unicellular organisms to mammals. Some antimicrobial peptides are already being evaluated in clinical studies, though many of them have not produced any favorable results in vivo due to their low metabolic stability and their toxicity, among other factors. Several strategies have been developed to overcome the above mentioned drawbacks, among them conjugation of microbial peptides with magnetic nanoparticles and chemical modification of their structure. The present study is aimed at reviewing the potential therapeutic applications of these molecules based on information published in MedLine, the Web of Science and Scopus in recent years.
Key words: antimicrobial peptides; susceptibility; infectious agents; antibiotics; nanoconjugates; chemical analogs.
INTRODUCCIÓN
Los péptidos antimicrobianos (PAMs) se descubrieron hace más de 90 años. Uno de los primeros reportados es la gramicidina, pues su descubrimiento ocurrió en 1939 y se demostró su efectividad para proteger frente a la infección por neumococos. En 1941, otro PAM, la tirocidina, fue descubierto y se encontró que presentaba actividad frente a bacterias grampositivas y gramnegativas. En el mismo año también se aisló a partir de la planta Triticuma estivum el PAM purotionina, que resultó ser efectivo contra hongos y bacterias patógenas.1
A pesar de que los primeros PAMs se descubrieron hace varios años, las investigaciones en este tema no tomaron relevancia hasta la década del ochenta cuando se hallaron las cecropinas y magaininas. Desde entonces se han aislado más de 5 000 PAMs de diversas fuentes y otros se han sintetizado mediante métodos químicos.1
En la actualidad estas moléculas han revolucionado la manera de tratar las enfermedades en humanos destacándose las causadas por agentes infecciosos, pues en las últimas décadas su tratamiento se ha complejizado debido a diferentes causas como la emergencia o re-emergencia de agentes infecciosos y al desarrollo de resistencia a los antibióticos convencionales.2 Otros factores como el incremento de la población inmunodeprimida natural (sida, desnutrición, ancianidad) o artificialmente (trasplantes, cáncer, enfermedades autoinmunes y alérgicas)3 también contribuyen a la aparición de dichas enfermedades.
En este sentido, los PAMs son una alternativa prometedora porque presentan propiedades terapéuticamente atractivas. Son considerados antibióticos naturales presentes en todas las formas de vida, desde organismos unicelulares hasta mamíferos.4 Destacan por presentar un amplio espectro de actividad frente a hongos, bacterias, virus y protozoos,5 por lo que tienen una importante acción protectora en organismos que carecen de sistema inmune mientras que en los vertebrados forman parte de la inmunidad innata.4 Además, no son afectados por las mutaciones que provocan resistencia a los antibióticos clásicos.6 Es por eso que este artículo tiene el objetivo de revisar las potenciales aplicaciones terapéuticas de estas moléculas, teniendo en cuenta la información publicada al respecto en MedLine, Web of Science y Scopus en los últimos años.
CARACTERÍSTICAS GENERALES DE LOS PAMs
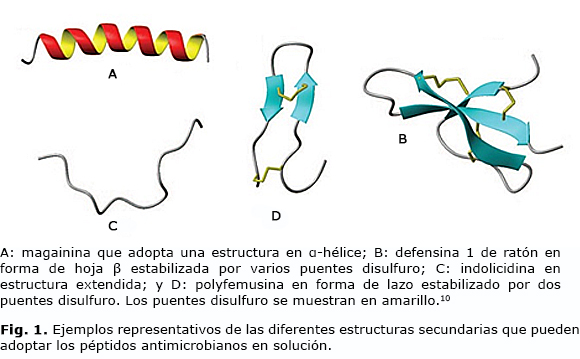
Los PAMs son moléculas de bajo peso constituidas por 10-100 aminoácidos, especialmente residuos de arginina, lo que hace que en su mayoría estén cargados positivamente. Generalmente son hidrofóbicos y anfipáticos a pH fisiológico. Son muy diversos, por lo que se han publicado numerosas clasificaciones teniendo en cuenta el organismo productor, célula blanco, mecanismo de acción, etc. Una de las más empleadas es la que se basa en su estructura secundaria en solución, por la cual los PAMs han sido divididos en 4 grupos: α-hélice, hoja β estabilizados por dos o tres puentes de disulfuro, estructuras extendidas con uno o más residuos predominantes (como triptófano y prolina) y en forma de lazo debido a la presencia de puentes simples de disulfuro (Fig. 1).7
Estas moléculas se encuentran codificadas en el material genético por familias multigénicas que incluyen a las catelicidinas, defensinas, cecropinas, dermaseptinas y pleurocidinas.8 Sufren diferentes modificaciones postraduccionales, las cuales pueden alterar su actividad de manera significativa. Estas incluyen la amidación del carboxilo terminal, halogenación, procesamiento proteolítico, isomerización aminoacídica, formación de puentes disulfuro y en menor medida glicosilación; por lo que se pueden formar múltiples isoformas de un mismo péptido como es el caso de las hepcidinas.9
MECANISMO DE ACCIÓN DE LOS PAMs
Los PAMs pueden presentar dos mecanismos de acción diferente. Algunos tienen acción directa sobre los microorganismos mientras que otros ejercen su actividad indirectamente a través de la activación del sistema inmune innato.11
Los PAMs que actúan directamente sobre los microorganismos pueden dividirse teniendo en cuenta el blanco celular. Una de estas dianas es la membrana citoplasmática, pues tienen gran afinidad por esta estructura en las células microbianas. Esto se debe fundamentalmente a las diferencias que existen entre las células microbianas y las del hospedero como la existencia de componentes aniónicos (lipopoplisacáridos, LPS, en bacterias gramnegativas, ácido teicoico y lipoteicoico en grampositivas o 1,3-β glucano y fosfomananos en las superficies fúngicas), composición y distribución fosfolipídica (fosfatidil glicerol y cardiolipina en bacterias y fosfatidilinositol en hongos) y diferencias en el potencial transmembrana (-130 a -150 mV en células de mamíferos y -90 a -110 en células microbianas).7 Ejemplo de estos PAMs lo constituyen la familia de las catelicidinas que se caracterizan por lisar la célula mediante perturbaciones directas en la membrana.12
Otra estructura que constituye un blanco para la acción antimicrobiana de estas moléculas es la pared celular. Inicialmente este mecanismo fue descrito para PAMs procarióticos como la nisina A pero después se estableció igualmente para algunos eucarióticos como la plectasina. Se conoce que la acción de la nisina A sobre la pared celular es a través de la formación de complejos con el lípido II, el cual es un precursor de esta estructura y, por tanto, inhibe su síntesis.13 Otros PAMs presentan blancos intracelulares, los cuales ejercen su actividad antimicrobiana a través de la inhibición de procesos esenciales para la célula como la síntesis de proteínas y la replicación del ADN. Se ha demostrado que los péptidos ricos en prolina como drosocina y apidaecina tienen la capacidad de translocarse hasta el citosol e inhibir la acción de la DnaK.14
Por su parte, los PAMs que tienen una acción indirecta sobre los microorganismos interactúan con el sistema inmune innato a través de diferentes rutas. De manera general, modulan la respuesta proinflamatoria y antiinflamatoria a través de la alteración de rutas de señalización, reclutan directa o indirectamente células efectoras como fagocitos, promueven la maduración de células dendríticas y la diferenciación de macrófagos, así como la apoptosis.15
POTENCIALIDADES TERAPÉUTICAS
Los PAMs pudieran ser empleados como agentes terapéuticos en el tratamiento de las enfermedades infecciosas y/o complementar la terapia con los antibióticos convencionales, pues tienen sinergismo con estos.16 En estudios realizados por Nuding y otros17 se demostró que los péptidos HBD1 - HBD3, HNP1, HD5, y LL-37 al combinarse con los antibióticos tigeciclina, moxifloxacina y el meropen tienen efecto sinérgico, lo cual favorece la actividad antimicrobiana contra cepas toxigénicas y no toxigénicas de Clostridium difficile. También la bacitracina ha mostrado este comportamiento en combinación con antibióticos de uso tópico como la neomicina y la polimixina B.18
También algunos PAMs como el LL-37 pueden neutralizar las endotoxinas bacterianas como el LPS evitando la secreción de citocinas proinflamatorias que pueden causar destrucción del tejido, shock e incluso la muerte.18 Esto demuestra que pudieran ser empleados como inmunomoduladores en enfermedades infecciosas y no infecciosas.
En cuanto a la actividad antiviral se conoce que las defensinas tienen actividad tanto contra virus envueltos como desnudos, aunque con mecanismos de acción diferentes. Algunas actúan directamente sobre las envolturas virales, glicoproteínas y cápsides mientras que otras se unen a los receptores celulares impidiendo su interacción con el virus o inhiben la replicación viral.19 Por ejemplo, las α-defensinas HNP-1 a la -3 y HD-5 han mostrado en estudios in vitro que pueden bloquear la infección por papilomavirus.20 También se ha descrito que las catelicidinas tienen efecto inhibitorio en la replicación in vitro de los lentivirus.21
Se ha demostrado recientemente que el péptido Ib-AMP-4 (derivado de la planta Impatiens balsamina) tiene una potente actividad frente a diferentes bacterias grampositivas y gramnegativas. En estudios realizados por Fan y otros22 se demostró que al combinarse con vancomicina inhibe el crecimiento de Enterococcus faecalis resistente a vancomicina (VRE). También se ha demostrado la eficacia de otros PAMs como el LZ1. Este péptido inhibe el crecimiento de Propionibacterium acnes así como de Staphylococcus epidermidis y Staphylococcus aureus. Además, se ha comprobado que inhibe la secreción de citoquinas proinfalamatorias como la interleuquina 1-β (IL 1-β).23
Se conoce que los PAMs antifúngicos pueden tener actividad fungicida o fungistática, y que pueden presentar o no actividad antibacteriana al mismo tiempo que ejercen su acción contra los hongos. Uno de los PAMs antifúngicos descrito es Cm-p5. En estudios realizados por Lopez-Abarrategui y otros24 se demostró que tiene actividad fungistática in vitro frente a Candida albicans con una concentración mínima inhibitoria de 10 µg/mL. Otros como NaD, presentan una potente actividad frente a varios hongos patógenos como Fusarium oxysporum, Botrytis cinerea, Aspergillus niger, Cryptococcus sp., así como a las levaduras Saccharomyces cerevisiae y C. albicans.25 La acción antifúngica de este péptido está mediada por la formación de gránulos en el citoplasma y la muerte celular.26
En cuanto a la actividad antiparasitaria se pueden mencionar como PAMs representativos los provenientes de insectos como la defensina A, la cual tiene actividad contra Plasmodium, Leishmania y Trypanosoma.27 También se ha descrito que el péptido recombinante HNP-1 (rHNP-1) tiene una potente actividad contra Leishmania durante todo su ciclo de vida.28
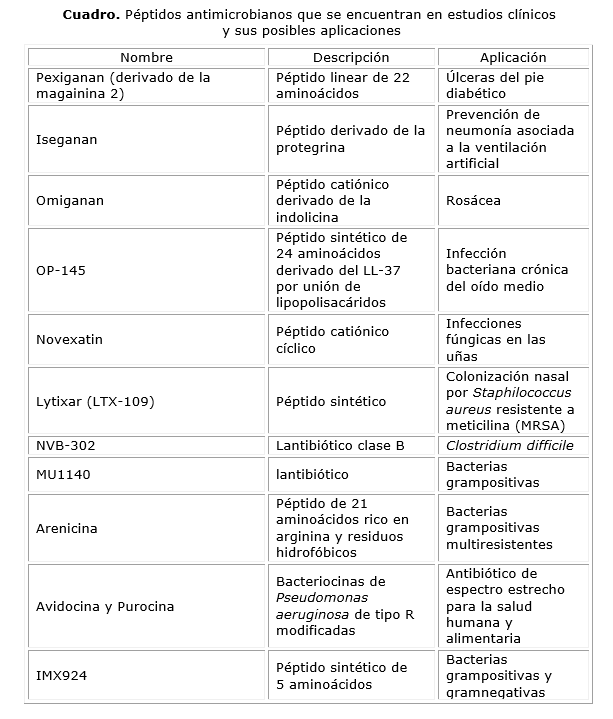
Algunos PAMs ya se encuentran en estudios clínicos (cuadro) como Pexiganan, Iseganan y Omiganan. El primero de estos es un derivado de 22 aminoácidos de la magainina 2 que muestra equivalencia con la fluoroquinona oral para el tratamiento de las úlceras en los pies de pacientes diabéticos. Comenzó a ser probado en 1999 para el tratamiento de las úlceras del pie diabético e intentó comercializarse con una formulación en crema bajo el nombre de Locilex, pero no fue aprobado por la Administración de Comida y Drogas debido a su baja eficacia. Iseganan se ha analizado para uso tópico y como tratamiento profiláctico de la neumonía asociada a la ventilación artificial; sin embargo, los resultados de estas investigaciones han sido desfavorables debido a la toxicidad del péptido. Omiganan (Fig. 2), análogo de la indolicina, ha sido capaz de reducir la colonización de los catéteres.29
Los PAMs que tienen acción indirecta sobre los microorganismos también se han evaluado en estudios clínicos. En estudios in vitro se ha demostrado la efectividad de derivados de la lactoferrina humana como hLF1-11 y formas recombinantes de la misma como la Talactoferrin-α frente a bacterias resistentes a los antibióticos de uso clínico y a cepas de C. albicans resistentes a fluconazol.29 Por su parte, la empresa canadiense Inimex Pharmaceuticals ha desarrollado PAMs con este mecanismo de acción y los datos preliminares muestran que disminuyen considerablemente la carga de S. aureus in vivo en un modelo de ratón.30
DESVENTAJAS ACTUALES Y ESTRATEGIAS DE OPTIMIZACIÓN
A pesar de las múltiples propiedades de los PAMS, no se han obtenido in vivo resultados satisfactorios, por lo que muchos de ellos no se emplean en tratamientos médicos. Estos péptidos presentan como desventajas la baja estabilidad metabólica y absorción oral, excreción rápida a través de riñones e hígado, elevada toxicidad e inmunogenicidad y altos costos de producción.31
Por ejemplo, se tienen evidencias de que el LL-37 provoca la liberación de histamina e incrementa la infiltración de neutrófilos provocando reacciones alérgicas e inflamación. Por otra parte, los PAMs son altamente susceptibles a sufrir degradación por enzimas secretadas por diferentes microorganismos. Ejemplo de esto es la degradación que sufre el LL-37 por la metaloproteinasa aureolisina de S. aureus. Además, otras proteasas propias del hospedero como las enzimas digestivas tripsina y quimotripsina también clivan a estos péptidos.32
Estas propiedades mencionadas anteriormente pueden ser mejoradas siguiendo disímiles estrategias. Algunas de ellas incluyen la inducción endógena de los PAMs propios mediante estimulantes administrados por vía sistémica o tópica, con lo cual se evitaría una respuesta inmune no deseada. Con respecto a esto se ha reportado que la inducción del LL-37 por la administración de fenilbutirato pudiera ser efectiva para el tratamiento de la shigellosis.33
La toxicidad es otra de las características de los PAMs que limita su empleo como agentes terapéuticos, es por ello que se sugiere el enmascaramiento de estas moléculas mediante diferentes formulaciones como las liposomales.29 Se ha demostrado que estas formulaciones potencian la actividad de los péptidos frente a microorganismos patógenos a la vez que disminuyen su toxicidad y susceptibilidad proteolítica. Esta estrategia fue evaluada por Yang y otros34 empleando el péptido WLBU-2, y se demostró que los liposomas modificados con este péptido presentaron actividad in vitro frente a cepas de MRSA y Pseudomonas aeruginosa.
Otra alternativa para optimizar la actividad de los PAMs es la obtención de nuevos péptidos a través de la síntesis de derivados cíclicos que contienen combinaciones de aminoácidos D y L, y la síntesis de PAMs por mimetismo molecular utilizando peptoides, que difieren de los péptidos en que la cadena lateral está unida a la estructura amídica nitrogenada en lugar de al carbono. Estas modificaciones hacen a los PAMs resistentes a las proteasas.35
En la actualidad también se emplea el diseño de péptidos sintéticos mediante modificaciones químicas de PAMs existentes en la naturaleza. Esta es una estrategia muy empleada en la actualidad por varios investigadores, pues permite el diseño de análogos de péptidos naturales. A partir de estos estudios se puede determinar la relación estructura-actividad de los péptidos con el objetivo de potenciar la actividad antimicrobiana de los PAMs naturales y de la misma manera disminuir la toxicidad frente a las células de mamíferos.36 Entre las modificaciones covalentes que pueden realizarse se encuentran ciclación, polimerización, síntesis de péptidos multiantigénicos y conjugación con proteínas portadoras, introducción de N-sustituciones o residuos peptoides, variación de los extremos N y C, peguilación, cambio de aminoácidos, fosforilación, glicosilación, lipidación o síntesis de híbridos péptido-esteroidales. Estos cambios en la estructura peptídica le confieren a los péptidos mayor selectividad y resistencia a las proteasas y aumentan la rigidez conformacional, según el caso.37
La lipidación de péptidos se realiza por incorporación de ácidos grasos, isoprenos y glicofosfolípidos a diferentes posiciones del péptido. Esto modifica la flexibilidad en la posición ocupada por el lípido,38 aumenta la hidrofobicidad y contribuye a la asociación con la membrana celular.39 Además, se conoce que los lipopéptidos presentan carácter anfifílico y se emplean como biosurfactantes y antibióticos.40 Muchas de estas moléculas también juegan un papel importante en los procesos de inducción de apoptosis y se reconocen por los receptores tipo Toll, componentes esenciales de la respuesta inmune.41 Dentro de este grupo de nuevos agentes antimicrobianos encontramos al MSI-843 (Oct-OOLLOOLOOL-NH2), lipopéptido obtenido a partir del acoplamiento del ácido octanoico al extremo N-terminal de la ornitina. Esta modificación incrementó la hidrofobicidad del derivado, el cual mostró actividad antimicrobiana tanto frente a bacterias como a hongos a bajas concentraciones.42
Por otra parte, la conjugación de cadenas peptídicas bioactivas con esteroides se ha convertido en los últimos años en una metodología exitosa para reproducir el efecto biológico de determinadas proteínas y péptidos naturales. Se conoce que la actividad biológica de polipéptidos y proteínas se debe a la disposición espacial que presentan determinadas secuencias de aminoácidos a lo largo de la cadena peptídica, factor que determina el tipo y alcance de las interacciones no-covalentes que tienen lugar durante dicho efecto biológico, por lo que al unir la secuencia peptídica responsable del efecto biológico a esqueletos rígidos se puede favorecer el proceso de reconocimiento molecular.43 El empleo del esqueleto esteroidal ha sido la estrategia más reciente, debido a que tiene la capacidad de imponer una determinada preorganización conformacional a las cadenas peptídicas, ya sea a través de la formación de estructuras rígidas macrocíclicas, como acíclicas, en las que las cadenas peptídicas quedan alineadas en disposición convergente.44
Banerjee y otros45 obtuvieron péptido-esteroides por introducción de análogos del tripéptido RGD (secuencia reconocida por varias integrinas) a esteroides reconocidos por membranas celulares, con elevada actividad antagonista de las ανβ3 integrinas, enzimas responsables del proceso de adhesión celular. Otros PAMs modificados químicamente incluyen el KSL con potente actividad bactericida y fungicida, y el péptido Bac8c derivado de la bactenecina, el cual presenta una potente actividad frente a un amplio grupo de bacterias gramnegativas y grampositivas así como de levaduras.46
Algunos de los péptidos que recientemente han sido modificados químicamente son el KABT-AMP y la uperina 3.6, los cuales presentan una estructura en α-hélice anfipática. En estudios realizados por Lum y otros36 se sintetizaron cuatro derivados (KU1, KU2, KU3 y KU4) de los mismos para potenciar su acción antifúngica, los cuales fueron obtenidos mediante la selección y fusión de fragmentos de estos PAMs. Estos péptidos híbridos mostraron una potente actividad frente a C. albicans en comparación con sus péptidos parentales y un efecto sinérgico en combinación con antifúngicos convencionales. Además, tuvieron baja actividad hemolítica pero significante actividad citotóxica en dos líneas celulares epiteliales humanas.
Con respecto a este tema, en Cuba se han realizado varias investigaciones con el objetivo de introducir modificaciones químicas en péptidos sintéticos y naturales para el desarrollo de candidatos terapéuticos y vacunales contra diferentes enfermedades. Ejemplo de esto es la polimerización de un péptido cíclico sintético derivado del lazo 4 de la proteína PorA de Neisseria meningitidis serogrupo B para aumentar su inmunogenicidad, lo cual permitiría su empleo como candidato vacunal. En este estudio se comprobó que esta estrategia mejora la respuesta inmune in vitro en varias líneas de ratones con respecto al péptido cíclico sin polimerizar.31
La nanotecnología también ha representado un gran avance. Los nanosistemas incrementan la estabilidad, eficacia y biodistribución de los PAMs47 además de que presentan un amplio espectro de actividad, pues son capaces de reconocer con elevada especificidad y selectividad a componentes bacterianos como el lipopolisacárido y glicopéptidos, ácidos nucleicos, así como a la bacteria completa.7 Su tamaño es suficientemente pequeño como para que permanezcan en el sistema circulatorio y pasen a través del sistema de capilares de los órganos y tejidos sin provocar la embolia de los vasos, y su elevada magnetización permite controlar su movimiento por la sangre e inmovilizarlos cerca del tejido patológico mediante un campo magnético.48 Estas son características favorables en las aplicaciones biomédicas in vivo, lo cual sugiere que la fusión de PAMs y nanopartículas magnéticas (NPMs) puede combinar las mejores propiedades de ambos, generando nanopartículas antimicrobianas con mejor actividad. Ejemplo de esto es la conjugación del péptido Cm-p5 con NPMs de ferrita de manganeso recubiertas con ácido cítrico. Con respecto a esto, Lopez-Abarrategui y otros48 evaluaron la actividad antifúngica del conjugado y se comprobó que presenta una mejor actividad in vitro frente a C. albicans en comparación con cada uno de sus componentes por separado. También registraron el mayor valor de CMI en 100 µg/mL a la vez que no observaron toxicidad frente a la línea celular de macrófagos RAW 264.7 a las concentraciones a las cuales tenía actividad antimicrobiana.
CONCLUSIONES
Los PAMs han sido estudiados con énfasis en los últimos años debido a los diversos efectos biológicos que presentan, pues pueden ser empleados por separado como antimicrobianos o en combinación con antibióticos debido a su efecto sinérgico, también como inmunomoduladores y/o compuestos neutralizadores de endotoxinas. Sin embargo, in vivo no han demostrado una buena actividad antimicrobiana, por lo que se siguen diversas alternativas para optimizar su acción con el objetivo de emplearlos como agentes terapéuticos en el futuro.
Conflictos de interés
Los autores no declaran conflictos de interés.
REFERENCIAS BIBLIOGRÁFICAS
1. Wang G. Computational Peptidology, Methods in Molecular Biology. In: Zhou P, Huang J (eds.). Improved Methods for Classification, Prediction, and Design of Antimicrobial Peptides. New York: Springer Science+Business Media; 2015. p. 44-66.
2. Arias CA, Murray BE. Antibiotic-Resistant Bugs in the 21st Century - A Clinical Super Challenge. N Engl J Med. 2009;360(5):439-43.
3. Keller M, Blench M, Tolentino H, Freifeld CC, Mandl KD, Mawudeku A, et al. Use of unstructured event-based reports for global infectious disease surveillance. Emerg Infect Dis. 2009;5(5):689-95.
4. Otero-Gonzalez AJ, Magalhaes BS, Garcia-Villarino M, Lopez-Abarrategui C, Sousa DA, Dias SC, et al. Antimicrobial peptides from marine invertebrates as a new frontier for microbial infection control. FASEB J. 2010;24(5):1320-34.
5. Guani-Guerra E, Santos-Mendoza T, Lugo-Reyes SO, Teran LM. Antimicrobial peptides: general overview and clinical implications in human health and disease. Clin Immunol. 2010;135(1):1-11.
6. Sanchez ML. Mecanismos de acción de péptidos antimicrobianos y mecanismos de resistencia de los patógenos. Bioquímica y Patología Clínica. 2016;80(1):36-43.
7. Lopez-Abarrategui C, Figueroa-Espi V, Reyes-Acosta O, Reguera E, Otero-Gonzalez AJ. Magnetic nanoparticles: new players in antimicrobial peptide therapeutics. Curr Protein Pept Sci. 2013;14(7):595-606.
8. Patrzykat A, Douglas SE. Antimicrobial peptides: cooperative approaches to protection. Protein Pept Lett. 2005;12(1):19-25.
9. Addo L, Ikuta K, Tanaka H, Toki Y,Hatayama M, Yamamoto M, et al. The three isoforms of hepcidin in human serum and their processing Int J Hematol. 2016;103(1):34-43.
10. Jenssen H, Hamill P, Hancock REW. Peptide Antimicrobial Agents. Clin Microbiol Rev. 2006;19(3):491-511.
11. Steinstraesser L, Kraneburg U, Jacobsen F, Al-Benna S. Host defense peptides and their antimicrobial-immunomodulatory duality. Immunobiology. 2011; 216(3):322-33.
12. Van der Weerden NL, Bleackley MR, Anderson MA. Properties and mechanisms of action of naturally occurring antifungal peptides. Cell Mol Life Sci. 2013;70(19):3545-70.
13. Scherer KM, Spille JH, Sahl H, Grein F, Kubitscheck U. The Lantibiotic Nisin Induces Lipid II Aggregation, Causing Membrane Instability and Vesicle Budding. Biophysical Journal.2015;108(5):1114-24.
14. Yi HY, Chowdhury M, Huang YD, Yu XQ. Insect antimicrobial peptides and their applications. Appl Microbiol Biotechnol. 2014;98(13):5807-22.
15. Mansour SC, Pena OM, Hancock RE. Host defense peptides: front-line immunomodulators. Trends Immunol. 2014;35(9):443-50.
16. De la Fuente NM, Villarreal J, Díaz MA, García AP. Evaluación de la actividad de los agentes antimicrobianos ante el desafío de la resistencia bacteriana. Revista Mexicana de Ciencias Farmacéuticas. 2016;46(2):7-16.
17. Nuding S, Frasch T, Schaller M, Stange EF, Zabel LT. Synergistic Effects of Antimicrobial Peptides and Antibiotics against Clostridium difficile. Antimicrob Agents Chemother. 2014;58(10):5719-25.
18. Zhang L, Falla TJ. Host defense peptides for use as potential therapeutics. Curr Opin Investig Drugs. 2009; 10(2):164-71.
19. Wilson SS, Mayim ME, Jason SG. Antiviral Mechanisms of Human Defensins. J Mol Bio. 2013;425(24):4965-80.
20. Wiens ME, Smith JG. Defensin HD5 Inhibits Human Papillomavirus 16 Infection via Capsid Stabilization and Redirection to the Lysosome. Mbio. 2017;8:1 e02304-16.
21. Steinstraesser L, Tippler B, Mertens J, Lamme E, Homann HH, Lehnhardt M, et al. Inhibition of early steps in the lentiviral replication cycle by cathelicidin host defense peptides. Retrovirology. 2005;2:2.
22. Fan X, Reichling J, Wing M. Antibacterial activity of the recombinant antimicrobial peptide Ib-AMP4 from Impatiens balsamina and its synergy with other antimicrobial agents against drug resistant bacteria. Pharmazie. 2013;68:628-30.
23. Zhang Z, Mu L, Tang J, Duan Z, Wang F, Wei L, et al. A small peptide with therapeutic potential for inflammatory acne vulgaris. PLoS One. 2013;8(8):e72923.
24. López-Abarrategui C, McBeth C, Mandal SM, Sun ZJ, Heffron G, Alba-Menéndez A, et al. Cm-p5: an antifungal hydrophilic peptide derived from the coastal mollusk Cenchritis muricatus (Gastropoda: Littorinidae). FASEB J. 2015;29:3315-25.
25. Hayes BM, Bleackley MR, Wiltshire JL, Anderson MA, Traven A, van der Weerden NL. Identification and mechanism of action of the plant defensin NaD1 as a new member of the antifungal drug arsenal against Candida albicans. Antimicrob Agents Chemother. 2013;57(8):3667-75.
26. Poon I, Baxter AA, Lay FT, Mills GD, Adda CG, Payne JA, et al. Phosphoinositide-mediated oligomerization of a defensin induces cell lysis. Elife. 2014;3:e01808.
27. Pretzel J, Mohring F, Rahlfs S, Becker K. Antiparasitic Peptides. Adv Biochem Eng Biotechnol. 2013;135:157-92.
28. Dabirian S, Taslimi Y, Zahedifard F, Gholami E, Doustdari F, Motamedirad M, et al. Human Neutrophil Peptide-1 (HNP-1): A New Anti-Leishmanial Drug Candidate. PLOS Neglected Tropical Diseases. 2013;7(10):e2491.
29. Fjell CD, Hiss JA, Hancock RE, Schneider G. Designing antimicrobial peptides: form follows function. Nature reviews Drug discovery. 2012;11(1):37-51.
30. Gordon YJ, Romanowski EG, McDermott AM. A review of antimicrobial peptides and their therapeutic potential as anti-infective drugs. CurrEye Res. 2005;30(7):505-15.
31. Garay HE. Síntesis de péptidos modificados químicamente con posibles aplicaciones farmacéuticas. Tesis presentada en opción al grado científico de Doctor en Ciencias Químicas. La Habana, Cuba: Universidad de La Habana; 2012.
32. Kim H, Jang JH, Kim SC, Cho JH. De novo generation of short antimicrobial peptides with enhanced stability and cell specificity. J Antimicrob Chemother Res Pract. 2014;69(1):121-32.
33. Sarker P, Ahmed S, Tiash S, Sultana R, Stromberg R, Andersson J, et al. Phenyl butyrate Counteracts Shigella Mediated Downregulation of Cathelicidin in Rabbit Lung and Intestinal Epithelia: A Potential Therapeutic Strategy. PLoS ONE.2011;6(6):e20637.
34. Yang K, Gitter B, Ruger R, Wieland GD, Chen M, Liu X, et al. Antimicrobial peptide-modified liposomes for bacteria targeted delivery of temoporfin in photodynamic antimicrobial chemotherapy. Photochem Photobiol Sci. 2011;10(10):1593-601.
35. Chongsiriwatana NP, Patch JA, Czyzewski AM, Dohm MT, Ivankin A, Gidalevitz D, et al. Peptoids that mimic the structure, function, and mechanism of helical antimicrobial peptides. Proc Natl Acad Sci USA. 2008;105(8):2794-9.
36. Lum KY, Tay ST, Le CF, Lee VS, Sabri NH, Velayuthan RD, et al. Activity of Novel Synthetic Peptides against Candida albicans. Sci Rep. 2015;5:9657.
37. Morales FE. Síntesis en fase sólida de péptidos N-sustituidos con lípidos y esteroides. Tesis en opción al grado científico de Master en Ciencias Químicas. La Habana, Cuba: Universidad de La Habana; 2014.
38. Wittmann M, Linne U, Pohlmann V, Marahiel MA. Role of DptE and DptF in the lipidation reaction of daptomycin. FEBS Journal. 2008;275:5343-54.
39. Nadolski MJ, Linder ME. Protein lipidation. FEBS Journal.2007; 274:5202-5210.
40. Kaiser A, Gaidzik N, Becker T, Menge C, Groh K, Cai H, et al. Synthetic Vaccines Consisting of Tumor-Associated MUC1 Glycopeptides and a Lipopeptide Ligand of the Toll-like Receptor 2. Angew Chem Int Ed. 2010;49:3688-92.
41. Hamley I, Kirkham S, Dehsorkhi A, Castelletto V, Rezac M. Ruokolainen J. Toll-like receptor agonist lipopeptides self-assemble into distinct nanostructures. Chem. Commun. 2014;50: 15948-51.
42. Thennarasu S, Lee DK, Tan A, Prasad U, Ramamoorthy A. Antimicrobial activity and membrane selective interactions of a synthetic lipopeptide MSI-843. Biochim Biophys Acta. 2005;1711(1):49-58.
43. Rivera DG, León F, Concepción O, Morales FE, Wessjohann LA. A Multiple Multicomponent Approach to Chimeric Peptide-Peptoid Podands. Chem Eur J. 2013;19(20):6417-28.
44. Rivera DG, Wessjohann LA. Architectural Chemistry: Synthesis of Topologically Diverse Macromulticycles by Sequential Multiple Multicomponent Macrocyclizations. J Am Chem Soc Rev. 2009;131:3721-32.
45. Banerjee A, Sergienko E, Vasile S, Gupta V, Vouri K, Wipf P. Triple Hybrids of Steroids, Spiroketals and Oligopeptides as New Biomolecular Chimeras. Org Lett. 2009;11(1):65-8.
46. Lee W, Lee DG. Fungicidal mechanisms of the antimicrobial peptide Bac8c. Biochim Biophys Acta. 2015;1848(2):673-9.
47. Brandelli A. Nanostructures as promising tools for delivery of antimicrobial peptides. Mini Rev Med Che. 2012;12:731-41.
48. Lopez-Abarrategui C, Figueroa-Espi V, Lugo-Alvarez MB, Pereira CD, Garay H, Barbosa JARG, et al. The intrinsic antimicrobial activity of citric acid-coated manganese ferrite nanoparticles is enhanced after conjugation with the antifungal peptide Cm-p5. International Journal of Nanomedicine. 2016;11:3849-57.
Recibido: 28 de febrero de 2017.
Aceptado: 17 de julio de 2017.
Anselmo J. Otero-González . Centro de Estudios de Proteínas. Calle 25 y J, Vedado, municipio Plaza de la Revolución, CP 10400. La Habana, Cuba. Correo electrónico: aotero@fbio.uh.cu


{kind=link}
{kind=link}
{kind=link}