










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Medicina Tropical
versión On-line ISSN 1561-3054
Rev Cubana Med Trop vol.70 no.3 Ciudad de la Habana set.-dic. 2018
ARTÍCULO ORIGINAL
Resistencia a insecticidas en cepas de Aedes aegypti (Diptera: Culicidae) de tres distritos de la Región Pacífico Central de Costa Rica
Insecticide resistance in Aedes aegypti (Diptera: Culicidae) strains from three districts of the Central Pacific Region of Costa Rica
Ólger Calderón-Arguedas, Karina Vargas, Adriana Troyo
Centro de Investigación en Enfermedades Tropicales. Departamento de Parasitología. Facultad de Microbiología. Universidad de Costa Rica (UCR). San José, Costa Rica.
RESUMEN
Introducción: Aedes aegypti es vector de virus dengue, chikungunya y Zika en Costa Rica. Su alta incidencia y carencia de vacunas hacen del control vectorial, incluyendo el químico, la única alternativa para disminuir su transmisión. El uso reiterativo de insecticidas propicia su resistencia.
Objetivo: Determinar la resistencia y mecanismos de detoxificación enzimática a temefós e insecticidas piretroides en cepas de Ae. aegypti de tres distritos de la Región Pacífico Central de Costa Rica.
Métodos: La resistencia a temefós, deltametrina y cipermetrina fue determinada en tres cepas de Barranca, Jacó y Quepos mediante bioensayos en larvas. Para cada insecticida se calculó la concentración letal 50 % (CL50) y un factor de resistencia 50 % (FR50), empleando la cepa Rockefeller como control. Ante la ocurrencia de resistencia, se repitieron los bioensayos utilizando butóxido de piperonilo, S,S,S, tributilfosforotritioato y ácido etacrínico que inhiben monooxigenasas, esterasas y glutatión S transferasa, respectivamente.
Resultados: Ninguna cepa mostró resistencia a temefós. Las cepas Barranca y Jacó fueron resistentes a deltametrina (FR50= 7,38; 28,23, respectivamente). La cepa Jacó mostró, adicionalmente, resistencia a cipermetrina (FR50= 7,70). La cepa Quepos no mostró resistencia a ningún piretroide. Solo la cepa Barranca mostró disminución de resistencia a deltametrina asociada al butóxido de piperonilo denotando vinculación con monooxigenasas (FR50: 10,10). Para los otros casos de resistencia, ninguno de los sinergistas disminuyó la resistencia.
Conclusiones: Las larvas de Ae. aegypti de las localidades evaluadas no mostraron resistencia a temefós. Respecto a piretroides se evidenció la aparición de resistencia que posiblemente no es mediada por detoxificación enzimática.
Palabras clave: resistencia; insecticidas; Aedes aegypti; arbovirus; enfermedades de transmisión vectorial; Costa Rica.
ABSTRACT
Introduction: Aedes aegypti is the vector of dengue, chikungunya, and Zika viruses in Costa Rica. The high incidence and the lack of vaccines make vector control, including chemical control, the only measure to prevent transmission. The repetitive use of insecticides may induce resistance.
Objective: To determine resistance and enzymatic detoxifying mechanisms to temephos and pyrethroids insecticides in strains of Ae. aegypti from three districts of the Central Pacific Region of Costa Rica.
Methods: Resistance to temephos, deltamethrin, and cypermethrin was determined in three strains of Barranca, Jacó, and Quepos by larval bioassays. In each test, the lethal concentration 50 % (LC50) and a factor of resistance 50 % (FR50) were calculated, using the Rockefeller strain as control. When resistance was observed, the bioassays were repeated using piperonyl butoxide, S,S,S, tributylphosphorotritioate, and ethacrynic acid, that inhibit monoxygenases, esterases, and glutathione S- transfererase, respectively.
Results: None of the strains were resistant to temephos. Resistance to deltamethrin (FR50= 7.38 and FR50= 28.23, respectively) was determined in the strains from Barranca and Jacó, while resistance to cypermethrin was detected only in Jacó (FR50= 7.70). The Quepos strain was not resistant to any pyrethroid. Only the Barranca strain showed a decrease in the resistance to deltamethrin when piperonyl butoxide was used, linking the resistance to monooxygenase enzymes (FR50: 10.10). For the other cases, none of the synergists decreased the resistance.
Conclusions: Larvae of Ae. aegypti from the localities evaluated were not resistant to temephos. With respect to pyrethroids, results show an emergence of resistance that may not be mediated by enzymatic detoxification.
Keywords: resistance; insecticides; Aedes aegypti; arbovirus; vector-borne diseases; Costa Rica.
INTRODUCCIÓN
Aedes aegypti es el principal vector arbovirus en ambientes urbanos.1 En Costa Rica su eliminación tuvo lugar en los años 60 y ocurrió gracias a las campañas basadas en el uso de DDT que enarboló la Organización Mundial de la Salud a finales de la década de 1950.2 Desde entonces hubo diversas reintroducciones, principalmente en las áreas portuarias, hasta que, para inicios de la década de 1990, se informó sobre la distribución del vector a lo largo de todo el país.3 En 1993 se documentaron los primeros casos de infección con virus Dengue (DENV) en la Región Pacífico Central y desde entonces el dengue constituye la principal enfermedad de transmisión vectorial de Costa Rica.3 Solo para el año 2013, se reportaron 49 993 casos de la virosis.4 La introducción de los virus chikungunya (CHIKV) en el año 2014 y Zika (ZIKV) en el 2015,5,6 que utilizan a Ae. aegypti para su propagación, ha subrayado la necesidad de implementar campañas efectivas para el control del vector.
Aunque el control ecológico, basado en la eliminación de criaderos, ha sido considerado como la alternativa más importante para impactar las poblaciones vectoriales, el control químico sigue siendo una alternativa viable para los sistemas de salud, sobre todo en condiciones de epidemia.7 En Costa Rica, durante la última década se han utilizado insecticidas piretroides como deltametrina y cipermetrina y organofosforados como temefós en el control del Ae. aegypti.8,9 Una de las tendencias que ha sido recurrente en los diferentes países donde se emplean estos químicos es la aparición de poblaciones de mosquitos resistentes.10 La generación de resistencia se produce por el uso indiscriminado de estos productos o por los errores en su aplicación, en los cuales tiene lugar la utilización de concentraciones sub letales o métodos de aplicación incorrectos.11 Estas condiciones representan presiones selectivas que favorecen la aparición de individuos resistentes.
Como parte de las labores de vigilancia de todo sistema de salud, y con la perspectiva de poder efectuar un uso racional de los insecticidas, es prioritario analizar el estado de resistencia de las cepas locales de Ae. aegypti. El propósito del presente estudio es determinar el estado de resistencia y mecanismos de detoxificación enzimática correspondientes a temefós e insecticidas piretroides en cepas de Ae. aegypti provenientes de tres localidades de la Región Pacífico Central de Costa Rica.
MÉTODOS
Cepas de Ae. aegypti. Las cepas de Ae. aegypti se establecieron a partir de larvas colectadas en un mínimo de 20 criaderos ubicados dentro de un radio de 2 km con respecto a cada centro poblacional. Las cepas y datos de colecta se muestran a continuación.
Cepa Barranca, procedente del distrito Barranca (9°58′N, 84°50′O), cantón Puntarenas, provincia Puntarenas, establecida en febrero del 2015.
Cepa Jacó, procedente del distrito Jacó (9°42′35″N, 84°36′52″O), cantón Garabito, provincia Puntarenas, establecida en agosto del 2015.
Cepa Quepos, procedente del distrito Quepos (09°25′52″N, 84°10′00″O), cantón Quepos, provincia Puntarenas, establecida en agosto del 2016.
La cepa Rockefeller, suministrada por el Instituto de Medicina Tropical "Pedro Kourí", La Habana, Cuba (IPK), fue utilizada como cepa de referencia susceptible.8
Insecticidas
Temefós: Fosforotritionato de o, o, o, o´-tetrametil-o, o´-tio-di-p-fenileno (IUPAC). Chem Sevice. West Chester Pennsylvania. Grado Analítico. 98,1 % de pureza.
Deltametrina: (1R,3R)-3-(2,2-dibromovinil)-2,2-dimetilciclopropano carboxilato de (S)-ciano3-fenoxibenzilo (IUPAC), Chem Sevice, West Chester Pennsylvania, grado analítico, 99,5 % de pureza.
Cipermetrina: (1RS)-cis,trans-3-(2,2-diclorovinil)-2,2-dimetilciclopropano carboxilato de (RS)-ciano-3-Fenoxibencileno (IUPAC). Sigma Aldrich Laborchemikalien. Grado técnico. 94,3 % de pureza.
Sinergistas
Butóxido de piperonilo (BP): Chem Sevice. West Chester Pennsylvania. Grado analítico. 98,2 % de pureza.
S,S,S Tributilfosforotritionato (DEF): Chem Sevice. West Chester Pennsylvania. Grado analítico. 97,5 % de pureza.
Ácido etacrínico (AE): Ácido 2-(2,3-dicloro-4-(2-metilideno-1oxibutil) fenoxi) acético. Abcam Biochemicals. Grado analítico. 99,0 % de pureza.
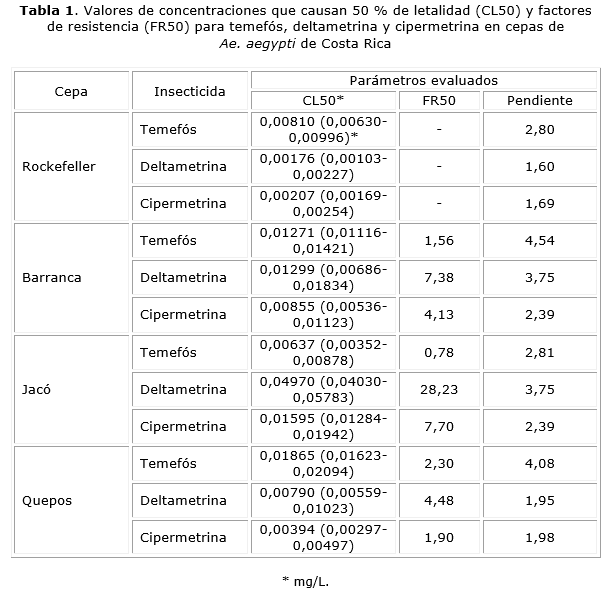
Bioensayos en larvas. Los bioensayos se efectuaron de acuerdo con metodologías descritas, en las cuales se utilizaron larvas de tercero y cuarto estadio provenientes las cepas de las Ae. aegypti en estudio.12 Se evaluaron, por quintuplicado, cinco concentraciones de los insecticidas que produjeron entre un 2 % y un 100 % de mortalidad en las larvas. Para cada réplica se utilizaron 20 larvas. Los insecticidas fueron disueltos en alcohol absoluto y 1,0 mL de cada solución madre fue diluida en 249,0 mL de agua libre de cloro para alcanzar la concentración de trabajo con la cual fueron expuestas las larvas. Los controles se trabajaron utilizando 1,0 mL de alcohol absoluto en lugar del insecticida. Los experimentos fueron efectuados en condiciones controladas a 27,8 °C ± 0,5 °C °C, 95 % de humedad relativa y un fotoperiodo de 12 h. La mortalidad se evaluó luego de 24 h, mediante la cuantificación de larvas muertas o moribundas. Posteriormente se efectuó un análisis probit-log, con el que se calculó la concentración letal 50 % (CL50). El valor de significancia utilizado como factor de heterogeneidad fue de 0,05. Para la ejecución de dicho análisis se empleó el programa SPSS v.11.5 para Windows®. La cepa control de susceptibilidad (cepa Rockefeller) se procesó de la misma forma descrita. Para cada cepa de campo se calculó un factor de resistencia 50 % (FR50) relativo a cada insecticida, que se calculó dividiendo la CL50 de dicha cepa entre la CL50 de la cepa Rockefeller. De acuerdo con los criterios de Mazarri y Georghiou, las cepas con FR50 inferiores a 5 se consideraron susceptibles, cepas con FR50 entre 5 y 10 mostraron resistencia incipiente y cepas con FR50 superiores a 10 fueron tipificadas como resistentes.13
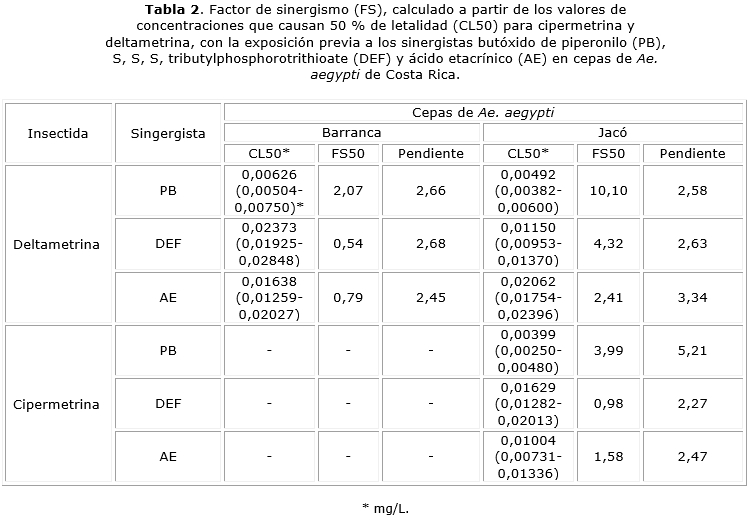
En los casos en que se evidenció algún tipo de resistencia y con el fin de perfilar el mecanismo de detoxificación correspondiente, se realizó una repetición de los ensayos sometiendo las larvas a concentraciones sub letales de los sinergistas por un periodo de 4 h previo a su exposición a los insecticidas. Las concentraciones empleadas fueron 5 mg/L para el PB, el cual es un inhibidor de la actividad citrocromo P450 monoxigenasa; 0,008 mg/L para el DEF, cuyo efecto es inhibitorio sobre actividades esterasa; y 5,0 mg/L para el AE, el cual es un inhibidor de la glutatión S-transferasa.8,14 Posteriormente se calculó un factor de sinergismo (FS50) en el que se relacionó la CL50 de cada insecticida en el ensayo inicial con su CL50 calculada luego de la exposición de las larvas al sinergista.
RESULTADOS
Con respecto a temefós, todas las cepas evaluadas mostraron FR 50 inferiores a 5 por lo que se determinó que no hubo resistencia a este insecticida (tabla 1). En relación con los piretroides, las cepas Barranca y Jacó mostraron resistencia a deltamentrina. En el caso de la cepa Barranca, el FR50 fue de 7,38 lo que denota una resistencia incipiente a este insecticida. Con respecto a la cepa Jacó, el FR50 fue de 28,23 evidenciándose una alta resistencia a dicho insecticida (tabla 1). Al aplicar el sinergista PB a la cepa de Jacó, la resistencia a la deltametrina disminuyó 10,10 veces (tabla 2), lo cual indicó que la resistencia estuvo dado por el sistema enzimático del citocromo P450 monooxigenasa. En el caso de la cepa Barranca, no se pudo determinar ningún efecto detoxificante relacionado con los sinergistas utilizados. Con respecto a cipermetrina, solo la cepa Jacó mostró una situación de resistencia incipiente a este insecticida que se evidenció por el cálculo de un FR50 de 7,70 (tabla 1). El tratamiento con sinergistas no generó disminución en el estado de resistencia a la cipermetrina en la cepa Jacó (tabla 2).
DISCUSIÓN
Las cepas de Ae. aegypti que fueron evaluadas en el presente estudio no mostraron resistencia a temefós. A pesar de que este insecticida es la principal alternativa para el control larvario del vector, las políticas implementadas en Costa Rica, dirigidas a la modificación ecológica mediante eliminación de criaderos, han evitado su uso masivo. Con esto se disminuye la presión selectiva que representa la aplicación repetitiva e indiscriminada de este insecticida. Este resultado coincide con lo encontrado recientemente en estudios realizados en localidades del Caribe de Costa Rica, donde también se encontró una condición de susceptibilidad a temefós.9,15 En otros países del mundo, la resistencia a temefós representa un problema significativo que reduce el repertorio de controladores para las poblaciones larvales de Ae. aegypti.16
En relación con la situación de los piretroides, tanto la cepa procedente de Barranca como la de Jacó mostraron algún grado de resistencia a estos químicos. La cepa de Barranca solo mostró resistencia incipiente a deltametrina, siendo el valor de su FR50 7,38. Esta condición no pudo ser reducida mediante la utilización de PB, ni de los otros sinergistas, lo que denota la posibilidad de que otros mecanismos de resistencia estén involucrados. En este sentido, en otros países se ha podido demostrar que mecanismos como Kdr asociados con mutaciones en los genes de las proteínas que conforman los canales de sodio dependientes de voltaje, se encuentran vinculados con la aparición de este tipo de resistencia.17,18 La cepa de Jacó presentó resistencia tanto a deltametrina como a cipermetrina con FR50 de 28,23 y 7,70, respectivamente. Estos resultados sugieren la posibilidad de que esta cepa pueda exhibir una condición de resistencia cruzada a insecticidas de esta familia.11 Para deltametrina, la disminución de la resistencia se pudo demostrar mediante el pretratamiento de las larvas con PB, lo que vincula en parte la ocurrencia de resistencia con el sistema citocromo P450 monooxigenasa que es el mecanismo más importante en la generación de resistencia a piretroides.19 Llama la atención el alto nivel de resistencia a deltametrina que se observó en esta cepa. Esta condición había sido descrita anteriormente por Bisset y otros en una cepa de este mismo distrito.8 Esta fue la razón por la cual se motivó la sustitución de deltametrina por cipermetrina como una política de control a nivel nacional. A más de media década de esta determinación, las cepas de este sitio siguen mostrando resistencia manifiesta a deltametrina. Dado el hecho de que estos insecticidas están químicamente relacionados, esta podría ser una de las razones por las cuales el proceso de reversión de la resistencia aún no ha tenido lugar completamente.
En el caso de la resistencia incipiente a cipermetrina que se observó en la cepa de Jacó, y al igual a lo que tuvo lugar con deltametrina en la cepa de Barranca, la reversión de la resistencia no ocurrió mediante el pretratamiento con PB. Ante el aumento en los reportes de resistencia a piretroides por parte de insectos plaga, las empresas manufactureras de insecticidas han empleado la adición de PB a sus formulados como una solución. El PB tiene dos formas de acción que conllevan la inhibición de enzimas vinculadas con el sistema citocromo P450 monooxigenasa y el incremento en la penetración cuticular del insecticida, dado su efecto disolvente para el insecticida y surfactante para la epicutícula.18 Es posible que la adición de este químico pueda representar, como suele ser lo usual en los sistemas biológicos, una presión selectiva que promueva la neutralización o reversión de estos mecanismos disminuyendo su efectividad.
Por otro lado, ha sido demostrado, en modelos experimentales, que el uso sostenido de PB puede generar cáncer hepatocelular en ratas.20 Estos procesos neoplásicos se han acompañado además de manifestaciones clínicas como trombocitemia esencial, hemorragias en estómago y ciego, anemia, lesiones degenerativas en alveolos y nefrotoxicidad.20 Además, la aplicación de insecticidas como cipermetrina en conjunto con PB parece provocar mayor toxicidad en ratas que la reportada para el insecticida solo, con afectación en el metabolismo de proteínas, degeneración de hígado y alteraciones en la función de hígado y riñones, principalmente en machos.21,22
De manera similar, estudios en conejos han demostrado que la exposición prolongada a cipermetrina y PB generan genotoxicidad e inflamación renal y hepática, con un mayor efecto observado cuando se administran simultáneamente.23 De esta forma, resulta urgente la racionalización en la toma de decisiones sobre el uso de químicos en control de vectores. Con esto se espera disminuir la ocurrencia de procesos de desarrollo de resistencia en vectores y evitar efectos tóxicos o degenerativos en el ser humano. El hecho de que la cepa de Quepos no mostrara resistencia a los insecticidas evaluados, subraya la necesidad de que las evaluaciones sobre resistencia se hagan de forma puntual, pues características del vector como su desplazamiento espacial limitado pueden generar tendencias y estados de resistencia diferenciados de un centro urbano a otro.
Agradecimientos
Los autores expresan su agradecimiento al Dr. Juan Bisset Lazcano del Instituto de Medicina Tropical "Pedro Kourí" de La Habana, Cuba, por haber suministrado la cepa Rockefeller para los ensayos efectuados; al Dr. Carlos Venegas, Director Regional del Ministerio de Salud de Costa Rica (MS) en la Región Pacífico Central y al Dr. Rodrigo Marín, coordinador general del Programa de Control de Vectores (MS) por su apoyo logístico. También desean agradecer a Iván Coronado y a Ariela Rojas de la Sección de Entomología Médica en la Universidad de Costa Rica (UCR) por su colaboración operativa y a la Vicerrectoría de Investigación (UCR) por su apoyo financiero al proyecto VI 803-B5-096.
Conflicto de intereses
No existe conflicto de intereses.
REFERENCIAS BIBLIOGRÁFICAS
1. Nene V, Wortman JR, Lawson D, Haas B, Kodira C, Tu ZJ, et al. Genome sequence of Aedes aegypti, a major arbovirus vector. Science. 2007;316(5832):1718-23.
2. Morice-Trejos A, Marín-Rodríguez R, Ávila-Agüero ML. El dengue en Costa Rica: evolución histórica, situación actual y desafíos. En: Escuela de Salud Pública (UCR). Estado actual, retos y perspectivas. Costa Rica: Sistema Editorial y de difusión de la información. Universidad de Costa Rica; 2010.
3. Troyo A, Porcelain SL, Calderón-Arguedas O, Chadee DD, Beier JC. Dengue in Costa Rica: the gap in local scientific research. Rev Panam Salud Pública. 2006;20:350-60.
4. Ministerio de Salud de Costa Rica. Situación del dengue 2013 [citado 20 sept 2017]. Disponible en: https://www.ministeriodesalud.go.cr/index.php/vigilancia-de-la-salud/analisis-de-situacion-de-salud
5. Corrales Aguilar E, Troyo A, Calderón-Arguedas O. Chikungunya: un virus que nos acecha. Acta Méd Costarric. 2015;57:7-15.
6. Calderón-Arguedas O, Moreira-Soto R, Troyo A. Zika Virus (ZIKA: New Emerging Pathogen Transmitted by Aedes Mosquitoes (Diptera: Culicidae) in the Latin American Subcontinent. Vector Biol J. 2016;1(1). DOI: 10.4172/2473-4810.1000103
7. Manjarres-Suarez, A, Olivero-Verbel J. Chemical control of Aedes aegypti: a historical perspective. Rev Costarric Salud Pública. 2013;22:68-75.
8. Bisset JA, Marín R, Rodríguez MM, Severson DW, French L, Díaz M, et al. Insecticide resistance in two Aedes aegypti (Diptera: Culicidae) strains from Costa Rica. J Med Entomol. 2013;50:352-61.
9. Calderón-Arguedas O, Troyo A. Evaluación de la resistencia a insecticidas en cepas de Aedes aegypti (Diptera: Culicidae) de la Región Caribe de Costa Rica. Rev Cubana Med Trop. 2016;68(1).
10. Smith LB , Kasai S, Scott JG. Pyrethroid resistance in Aedes aegypti and Aedes albopictus: Important mosquito vectors of human diseases. Pesticide Biochemistry and Physiology 2016 [citado 20 sept 2017];133:1-12. Disponible en: https://doi.org/10.1016/j.pestbp.2016.03.005
11. Bisset JA. Uso correcto de insecticidas: control de la resistencia. Rev Cubana Med Trop. 2002;54:202-19.
12. Bisset J, Blanco S, Braga I, Coto H, Massuh H, Moncayo A, et al. Protocolo para determinar la susceptibilidad o resistencia a insecticidas de mosquitos de la especie Ae. aegypti. Puerto Iguazú: Fundación Mundo Sano/Red Latinoamericana de Control de Vectores (RELCOV); 2005.
13. Mazzari MB, Geroghiou GP. Characterization of resistance to organophosphate, carbamate, and pyrethroid insecticides in field populations of Aedes aegypti from Venezuela. J Am Mosq Cont Assoc. 1995;11:315-22.
14. Bisset JA, Rodríguez MM, Cáceres L. Niveles de resistencia a insecticidas y sus mecanismos en 2 cepas de Aedes aegypti de Panamá. Rev Cubana Med Trop. 2003;55:191-5.
15. Calderón-Arguedas O, Troyo A. Perfil de resistencia a insecticidas en una cepa de Aedes aegypti (Linnaeus) de la región Caribe de Costa Rica. Rev Cubana Med Trop. 2014;66:351-9.
16. Moyes CL, Vontas J, Martins AJ, Ng LC, Koou SY, Dusfour I, et al. Contemporary status of insecticide resistance in the major Aedes vectors of arboviruses infecting humans. PLOS Neglected Tropical Diseases. 2017 [citado 20 sept 2017];11(7):e0005625. Disponible en: https://doi.org/10.1371/journal.pntd.0005625
17. Harris AF, Rajatileka S, Ranson H. Pyrethroid Resistance in Aedes aegypti from Grand Cayman. Am J Trop Med Hyg. 2010;83:277-84. doi: 10.4269/ajtmh.2010.09-062.
18. Bingham G, Strode C, Tran L, Khoa PT, Jamet HP. Can piperonyl butoxide enhance the efficacy of pyrethroids against pyrethroid-resistant Aedes aegypti? Trop Med Int Health. 2011;16:492-500. doi: 10.1111/j.1365-3156.2010.02717.x.
19. Hardstone M, Leichter C, Harrington LC, Kasai S, Tomita T, Scott JG. Cytochrome P450 monooxygenase-mediated permethrin resistance confers limited and larval specific cross-resistance in the southern house mosquito, Culex pipiens quinquefasciatus. Pesticide Biochemistry and Physiology. 2007 [citado 20 sept 2017];89:175-84. Disponible en: https://doi.org/10.1016/j.pestbp.2007.06.006
20. Takahashi O, Oishi S, Fujitani T, Tanaka T, Yoneyama M. Chronic toxicity studies of piperonyl butoxide in F344 rats: induction of hepatocellular carcinoma. Fundam Appl Toxicol. 1994;22:293-303
21. Yavuz O, Aksoy A, Das YK, Gulbahar MY, Yarim GF, Cenesiz M, et al. Repeated-dose 14-day dermal toxicity of different combinations of some synthetic pyrethroid insecticides, piperonyl butoxide, and tetramethrin in rats. Cutaneous and Ocular Toxicology. 2010;29:16-25.
22. Yavuz O, Aksoy A, Das YK, Gulbahar MY, Guvenc D, Atmaca E, et al. Subacute oral toxicity of combinations of selected synthetic pyrethroids, piperonyl butoxide, and tetramethrin in rats. Toxicol Ind Health. 2015;31:289-97.
23. Vardavas AI, Stivaktakis PD, Tzatzarakis MN, Fragkiadaki P, Vasilaki F, Tzardi M, et al. Long-term exposure to cypermethrin and piperonyl butoxide cause liver and kidney inflammation and induce genotoxicity in New Zealand white male rabbits. Food Chem Toxicol. 2016;94:250-9. doi: 10.1016/j.fct.2016.06.016.
Recibido: 3 de octubre de 2017.
Aprobado: 8 de agosto de 2018.
Ólger Calderón-Arguedas. Facultad de Microbiología. Universidad de Costa Rica. Oficina 208. Código Postal 2060. San José, Costa Rica. Correo electrónico: olger.calderon@ucr.ac.cr


{kind=link}
{kind=link}