










 Serviços customizados
Serviços customizados Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Entre los animales capaces de atacar a los seres humanos y provocar graves lesiones se destacan las serpientes venenosas, que están ampliamente distribuidas por diversos continentes y son encontradas principalmente en el medio rural, lo que genera riesgos a la salud de las personas que allí habitan o trabajan.1 De acuerdo con los datos de la Organización Mundial de la Salud (OMS), anualmente ocurren miles de accidentes ofídicos en distintos grados (leve, moderado o grave) y con signos/síntomas clínicos peculiares.2 Específicamente en Brasil, según datos publicados por el Ministerio de Salud brasileño, el número de mordeduras de serpientes venenosas ha aumentado significativamente en todas las regiones del país y, según la misma fuente, entre 2012 y 2017 se han notificado 443 912 accidentes ofídicos con 1 815 óbitos.3
En vista de esto, dependiendo de la región anatómica afectada, la cantidad de veneno inoculada en el organismo de la víctima, y principalmente el tiempo transcurrido entre el accidente y el inicio del tratamiento, el cuadro clínico del accidentado puede desarrollar síntomas inmediatos y tardíos con repercusión local o sistémica, según el género de la serpiente agresora.4
Como el veneno de estos animales es formado por diversas enzimas con actividad catalítica dirigida a moléculas diana específicas, la víctima puede manifestar dolor, edema, equimosis e incluso mionecrosis en el sitio de la mordedura.5,6,7 Desde el punto de vista sistémico, es común el surgimiento de los trastornos de la hemostasia, hemorragia, miotoxicidad y neurotoxicidad, entre otros.6,7,8,9,10 Por consiguiente, las personas afectadas pueden ser totalmente sanadas, desarrollar secuelas permanentes, o incluso llegar a la muerte.4
Aunque el veneno agreda intensamente el organismo del paciente, el tratamiento más efectivo ya desarrollado, casi siempre disponible y aceptado por la OMS para el tratamiento del envenenamiento ofídico es la sueroterapia.11 Sin embargo, su acción en el sitio de la mordedura no es tan prometedora, ya que no bloquea totalmente los daños en el tejido de la región afectada por la baja concentración del suero en los tejidos más periféricos. Debido a eso y al difícil acceso a la sueroterapia en áreas más aisladas, la medicina popular siempre utilizó el potencial de las plantas como alternativa para el tratamiento del ofidismo.12
La fisiología de las plantas es bastante compleja y proporciona la síntesis de diversos metabolitos secundarios como forma de defensa contra depredadores.13 Como la flora siempre ha sido vista como fuente de materia prima para el tratamiento de diversas enfermedades, muchas investigaciones incluyen la purificación de moléculas con potencial antiofídico. Así, las investigaciones de Santhosh y otros14 relatan la existencia de diversos compuestos antiofídicos aislados de plantas y concluyen que estos metabolitos serían útiles como tratamiento auxiliar después del envenenamiento.
Teniendo en cuenta la gran importancia de los accidentes ofídicos para la salud pública, el objetivo de este trabajo fue evaluar si los extractos de Jatropha curcas (J.c), Myrsine guianensis (M.g) y Zanthoxylum monogynum (Z.m) son capaces de inhibir los efectos tóxicos de los venenos de Bothrops alternatus (B. alternatus) y Bothrops moojeni (B. moojeni).
MÉTODOS
Venenos y animales, identificación de las plantas y preparación de los extractos vegetales
Se adquirieron los venenos de las serpientes B. alternatus y B. moojeni a partir de especímenes mantenidos en el Serpentario Bioagents (Batatais, São Paulo, Brasil) y se almacenaron a 20 ºC hasta el momento de su uso. Los animales, ratones machos Balb/c de 20-25 g de peso, fueron proporcionados por el Centro de Bioterismo e Experimentação Animal (CBEA) de la Universidade Federal de Uberlândia (UFU), después de la apreciación y aprobación de todos los protocolos experimentales por la Comisión de Ética en el Uso de Animales (Protocolo CEUA/UFU: 008/12).
Se colectaron las hojas de M.g y J.c en el área rural de la ciudad Uberlândia, Minas Gerais, Brasil. Después del proceso de identificación de cada especie vegetal por el Instituto de Biología, Laboratório de Anatomia e Desenvolvimento Vegetal de la UFU, se depositó una exsiccata de cada especie. El método de Izidoro y otros,15 fue utilizado para procesar este material vegetal.
Brevemente, las hojas lavadas de cada planta, fueron sumergidas en agua desionizada y trituradas en licuadora durante 15 min. Se centrifugó la parte líquida a 30 000/g durante 20 min. Se congeló el sobrenadante en nitrógeno líquido, se liofilizó y se almacenó a -80 ºC. El extracto de Z.m fue donado por los profesores Dr. Jeferson C. do Nascimento y Dra. Vanderlúcia F. de Paula de la Universidade Estadual do Sudoeste da Bahia, Victoria da Conquista, Bahia, Brasil.
Dosificación de proteínas
Se determinó la concentración de proteínas por el método descrito por Bradford16 de acuerdo con una curva estándar de albúmina de suero bovino.
Inhibición in vitro de la actividad fosfolipasa (PLA 2 ) por los extractos de J.c, M.g y Z.m
Se determinó la actividad fosfolipasa por titulación potenciométrica según el método previamente descrito por De Haas y Postema.17 Una emulsión de yema de huevo en presencia de desoxicolato de sodio 0,03 M y cloruro de calcio (CaCl2) 0,6 M fue usada como substrato. Durante la realización del ensayo, se titularon los ácidos grasos liberados enzimáticamente con una solución estándar de hidróxido de sodio (NaOH) 0,0975 M en pH 8,0 a temperatura ambiente.
Se realizaron cuatro repeticiones (n= 4) de todos los ensayos de la actividad fosfolipasa (PLA2) acompañadas por controles positivos, realizados solamente con 10 (g de veneno de B. alternatus o B. moojeni diluidas en 100 (L del substrato, y controles negativos, realizados solamente con 100 (g de J.c, 100 (g de M.g o 500 (g de Z.m diluidas en 100 (L del substrato. Las pruebas de inhibición con los extractos se realizaron con las mismas cantidades de substrato y masa de veneno previamente incubados y en proporciones crecientes de veneno:extracto (1:0,5, 1:1, 1:5, 1:10, 1:20) (m:m); cuyas concentraciones variaron de 5 a 100 (g para los extractos de M.g y J.c. Para el extracto de Z.m, las proporciones veneno:extracto fueron de 1:1, 1:2,5, 1:5, 1:10 y 1:50 (m:m), con concentraciones de 10 a 500 (g. La actividad fosfolipasa fue calculada a partir de la cantidad de microequivalentes de base consumida/microgramo de proteína utilizada y los resultados fueron expresados como media y desviación estándar (n= 4) de los porcentajes de inhibición.
Inhibición in vitro de la actividad coagulante por los extractos de J.c, M.g y Z.m
El ensayo de actividad coagulante fue realizado cuatro veces (n= 4), conforme la técnica descrita por Assakura y otros.18 Se registró el tiempo necesario para coagular las mezclas de plasma con el analizador Quick-Timer (Drake), que utiliza un sistema óptico que detecta la variación brusca de densidad óptica de la muestra en el momento de la coagulación y cuyo tiempo de medición máximo es de 120 s. Los controles positivos para este ensayo fueron 150 (L de plasma bovino incubado con 10 (g de veneno de B. alternatus o de B. moojeni. Las pruebas de inhibición con los extractos se ejecutaron con las mismas cantidades de plasma y masa de veneno previamente incubadas con concentraciones crecientes de 10 a 100 (g de los extractos de M.g y J.c y de 10 a 500 (g del extracto de Z.m.
La variación de las proporciones veneno/extracto para los extractos de M.g y J.c fue 1:1, 1:5, 1:10 y 1:20 (m:m) y para el extracto de Z.m, 1:1, 1:2,5, 1:5, 1:10 y 1:50 (m:m). Los controles negativos de cada extracto se realizaron con una mezcla de 150 (L de plasma bovino incubado con 100 (g de extracto de M.g, 100 (g de extracto de J.c o 500 (g del extracto liofilizado de Z.m, respectivamente. El tiempo necesario para la formación de la red de fibrina fue considerado como el tiempo de coagulación, considerando que un aumento en este tiempo es sinónimo de la inhibición de la actividad de coagulación. Todos los resultados fueron presentados con los valores de la media y las respectivas desviaciones estándar.
Inhibición in vivo de la actividad hemorrágica por los extractos de J.c y M.g
Este ensayo se realizó según la metodología descrita por Nikai y otros (19 con algunas modificaciones. Para determinar la inhibición de la hemorragia inducida por el veneno de B. moojeni por el extrato de J.c, ratones Balb/c machos de 20-25 g de peso fueron distribuidos inicialmente en 6 grupos (n= 4). Cada animal del grupo I (control positivo) recibió una inyección intradérmica en la región dorsal que contenía 25 (g de veneno de B. moojeni diluido en 50 (L de PBS. Los animales de los grupos GII, GIII, GIV e GV (grupos experimentales) recibieron de la misma manera una inyección que contenía una mezcla de veneno:extracto en las siguientes proporciones 1:1, 1:5, 1:10 y 1:20 (m:m), respectivamente, correspondientes a una concentración creciente de 25 a 500 (g. Los animales del grupo GVI (control negativo) recibieron por la misma vía solamente el extracto de J.c (500 (g). Después de 3 h, los animales fueron anestesiados con quetamina( 10 % (0,05 mL/kg) + xilacina( 2 % (0,025 mL/kg) y su piel fue retirada para medir el halo hemorrágico con un calibre digital DIGMESS 100.174BL y así calcular el área hemorrágica. Posteriormente, a los animales se les realizó la eutanasia por dislocación cervical. Para determinar la inhibición de la actividad hemorrágica por el extracto de M.g, se realizaron los mismos procedimientos metodológicos. Todos los resultados fueron presentados con los valores de las medias y sus respectivas desviaciones estándar.
Análisis estadísticos de los datos
Las medias de cada ensayo y sus respectivas desviaciones estándar fueron obtenidas usando la prueba estadística t-test con un nivel de significancia del 5 % para la comparación entre los grupos. Los análisis estadísticos, así como las figuras, fueron realizados en la versión 4.0 del programa GraphPad Prism (GraphPad Software).
RESULTADOS
Inhibición in vitro de la actividad fosfolipasa inducida por el veneno de B. alternatus por medio de los extractos de M.g y J.c
Como puede observarse en la figura 1, los extractos de J.c y M.g fueron capaces de inhibir la actividad fosfolipasa inducida por el veneno de B. alternatus. En comparación con el veneno bothrópico (VB) crudo (0 % de inhibición), el extracto de J.c (Figura 1A) inhibió la actividad fosfolipasa aproximadamente un 66 %, ya en la razón 1:0,5 (veneno:extracto, m:m). En el resto de las razones, 1:1, 1:5, 1:10 y 1:20 (veneno:extracto, m:m), hubo un 100% de inhibición. Para el extracto de M.g (Figura 1B), se observó un 100 % de inhibición a partir de la razón 1:5 (veneno:extracto, m:m) en comparación con el VB crudo (0 % de inhibición). Ensayos realizados solamente con el extracto vegetal (EV) de J.c o M.g demostraron que estos extractos no inducen este tipo de actividad enzimática (Figuras 1A y 1B, respectivamente).
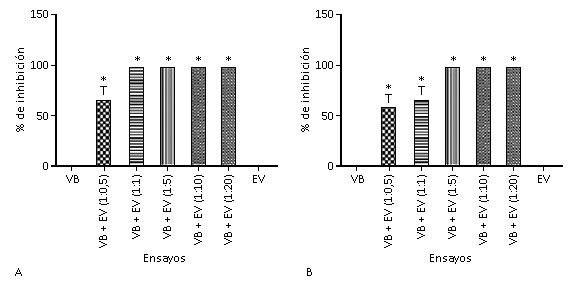 A: extracto de J.c. B: extracto de M.g. VB: veneno bothrópico (control positivo). EV: extracto vegetal (control negativo). VB + EV: veneno bothrópico incubado con extracto de M.g o J.c en diferentes proporciones.*Diferencia estadísticamente significativa en relación al control positivo (p< 0,05), n= 4.
A: extracto de J.c. B: extracto de M.g. VB: veneno bothrópico (control positivo). EV: extracto vegetal (control negativo). VB + EV: veneno bothrópico incubado con extracto de M.g o J.c en diferentes proporciones.*Diferencia estadísticamente significativa en relación al control positivo (p< 0,05), n= 4.Fig. 1 Porcentaje de inhibición de la actividad fosfolipasa inducida por el veneno de B. alternatus sobre los fosfolípidos.
Inhibición in vitro de la actividad coagulante inducida por el veneno de B. alternatus por medio de los extractos de M.g y J.c
La figura 2 muestra la inhibición, gracias a la acción de los extractos de M.g y J.c, de la conversión del fibrinógeno soluble, existente en el plasma bovino, en fibrina insoluble inducida por el veneno de la serpiente B. alternatus. En comparación al control positivo (VB), el extracto de M..g (Fig. 2, A) prolongó el tiempo de coagulación del plasma de forma parcial, pero significativa, en las proporciones veneno:extracto 1:5 y 1:10 (m:m). En la proporción 1:20 (m:m), el plasma fue incoagulable. En cuanto al extracto de J.c (Figura 2B), fue capaz de prolongar parcial y significativamente el tiempo de coagulación del plasma en las proporciones 1:1 y 1:5 (m:m). En las otras proporciones, 1:10 y 1:20 (m:m), se observó que el plasma era incoagulable. Por su parte, los controles negativos, únicamente extractos de M.g (Figura 2A) o J.c (Figura 2B), no indujeron la coagulación del plasma.
 A: Extracto de M.g. B: extracto de J.c. VB: veneno bothrópico (control positivo). EV: extracto vegetal (control negativo). VB + EV: veneno bothrópico incubado con extracto de M.g o J.c en diferentes proporciones.*Diferencia estadísticamente significativa en relación al control positivo (p< 0,05), n= 4.
A: Extracto de M.g. B: extracto de J.c. VB: veneno bothrópico (control positivo). EV: extracto vegetal (control negativo). VB + EV: veneno bothrópico incubado con extracto de M.g o J.c en diferentes proporciones.*Diferencia estadísticamente significativa en relación al control positivo (p< 0,05), n= 4.Fig. 2 Inhibición de la actividad coagulante inducida por el veneno de B. alternatus sobre el plasma bovino citratado.
Inhibición in vivo de la actividad hemorrágica inducida por el veneno de B. alternatus por medio de los extractos de J.c y M.g
La inhibición de la actividad hemorrágica inducida en ratones por la administración del veneno crudo de B. alternatus debida a los extractos de J.c y M.g se muestra en la figura 3. La inhibición de esta actividad por el extracto de J.c fue significativa a partir de la proporción 1:5 (m:m), mientras que a partir de la proporción 1:10 (m:m) alcanzó una inhibición del 100% (Fig. 3A). En relación con el extracto de M..g, la inhibición fue efectiva a partir de la razón 1:1 (m:m) alcanzando el 100 % en las razones 1:10 y 1:20 (m:m) (Fig. 3, B), en comparación con el control positivo (VB). Como se observa en las figuras 3, A y 3, B, la aplicación exclusiva de los extractos de J.c y M.g no indujo hemorragia en los animales usados en el experimento (control negativo, EV).
 A: extracto de J.c. B: extracto de M.g. VB: veneno bothrópico (control positivo). EV: extracto vegetal (control negativo). VB + EV: veneno bothrópico incubado con extracto de J.c o M.g en diferentes proporciones.*Diferencia estadísticamente significativa en relación al control positivo (p< 0,05); n = 4.
A: extracto de J.c. B: extracto de M.g. VB: veneno bothrópico (control positivo). EV: extracto vegetal (control negativo). VB + EV: veneno bothrópico incubado con extracto de J.c o M.g en diferentes proporciones.*Diferencia estadísticamente significativa en relación al control positivo (p< 0,05); n = 4.Fig. 3 Inhibición del área hemorrágica inducida por el veneno de B. alternatus bajo la piel, en el dorso de ratones.
Inhibición in vitro de la actividad coagulante inducida por el veneno de B. moojeni por medio del extracto de Z.m
Según los datos presentados en la tabla, el extracto de Z.m y la solución de metanol (SA) no provocaron coagulación plasmática durante el tiempo de experimentación (controles negativos), mientras que el veneno de B. moojeni indujo la coagulación del plasma bovino en aproximadamente 18 segundos. Sin embargo, cuando el veneno fue incubado con extracto de Z.m (veneno:extracto, m:m), hubo una prolongación en el tiempo de coagulación dependiente de la dosis a partir de la razón 1:10 (veneno:extracto, m:m) que fue estadísticamente significativa.
Tabla Inhibición de la actividad coagulante inducida por el veneno de B. moojeni por medio del extracto de Z.m sobre plasma bovino citratado
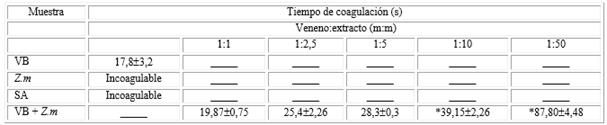
VB: veneno bothrópico (control positivo). EV: extracto vegetal (control negativo).
SA: solución de metanol (control negativo). VB + EV: veneno de B. moojeni incubado con extracto de Z.m en diferentes proporciones.
*Diferencia estadísticamente significativa en relación al control positivo (p< 0,05), n= 4.
Actividad fosfolipasa
El veneno de B. moojeni presentó una intensa actividad fosfolipasa (control positivo), mientras que el extracto de Z.m y la solución de metanol no indujeron la hidrólisis de los fosfolípidos del substrato utilizado (controles negativos). En los ensayos experimentales de inhibición, el veneno incubado con el extracto de Z.m en diferentes proporciones (veneno:extracto, m:m) indujo una disminución estadísticamente significativa de la propiedad enzimática estudiada a partir de la proporción 1:5 en comparación con el control positivo (Fig. 4).
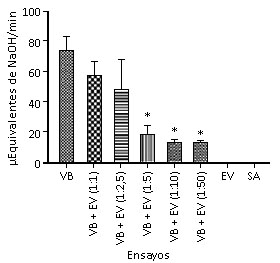 VB: veneno bothrópico (control positivo). EV: extracto vegetal (control negativo). SA: solución de metanol (control negativo). VB+EV: veneno bothrópico incubado con extracto vegetal en diferentes proporciones.*Diferencia estadísticamente significativa en relación al control positivo (p< 0,05), n= 4.
VB: veneno bothrópico (control positivo). EV: extracto vegetal (control negativo). SA: solución de metanol (control negativo). VB+EV: veneno bothrópico incubado con extracto vegetal en diferentes proporciones.*Diferencia estadísticamente significativa en relación al control positivo (p< 0,05), n= 4.Fig. 4 Inhibición de la actividad fosfolipasa inducida por el veneno de B. moojeni por medio del extracto de Z.m.
DISCUSIÓN
La importancia médica y económica de las plantas con potencial medicinal ha motivado varios estudios científicos y etnofarmacológicos, los cuales han resultado en la identificación de muchas moléculas con propiedades farmacológicas.13 Con base en la botánica, varias plantas han sido utilizadas desde la antigüedad con el propósito de neutralizar efectos deletéreos provocados por los venenos de serpientes, principalmente en regiones donde la sueroterapia es escasa.20
Dependiendo de la familia y del género de la serpiente agresora, la víctima puede desarrollar tanto efectos locales como sistémicos.21 Entre los diversos componentes de los venenos de las serpientes pertenecientes a la familia Viperidae, y principalmente al género Bothrops, se destacan las fosfolipasas A2 (PLA2).11 Estas fosfolipasas A2 son enzimas responsables por la hidrólisis de fosfolípidos de las bicapas lipídicas y por la desestructuración del ambiente celular, desencadenando así los efectos iniciales del envenenamiento, como dolor, edema e hiperemia que posteriormente evolucionan para un típico proceso de inflamación aguda.22 En el presente estudio, se ha verificado que los venenos de las serpientes B. alternatus y B. moojeni son fuentes de fosfolipasas A2 y que inducen actividad fosfolipasa in vitro. Además, cuando estos venenos son incubados con los extractos de J.c y M. g, el 100 % de la actividad fosfolipasa es inhibida en las proporciones 1:1 y 1:5 (m:m). Sin embargo, el veneno de B. Moojeni tratado con el extracto de Z.m es parcialmente inhibido.
En la literatura, hay miles de investigaciones en relación con la inhibición de la toxicidad de venenos de serpientes de diferentes géneros y enzimas fosfolipasas A2 a partir del extracto bruto de plantas pertenecientes a diversas familias como Piper longum L.,23Connarus favosus,24Rosmarinus officinalis L.25 y Cassampelos pareira; (26) al igual que a partir de principios activos con potencial antiofídico como el ácido rosmarínico, (27 el tanino aislado de Mimosa pudica, (28 el ácido arjunólico29 y el (-sitosterol,30 entre otros.
De acuerdo con White,31 los venenos de las serpientes, especialmente los bothrópicos, presentan componentes que afectan directamente a la hemostasia. Por presentar acción semejante a la trombina, estos componentes son llamados de thrombin-like y son responsables por la conversión in vivo del fibrinógeno plasmático soluble en fibrina. Estas informaciones fueron totalmente corroboradas por este trabajo, ya que los venenos bothrópicos estudiados indujeron fuertemente la coagulación del plasma. Sin embargo, cuando fue incubado previamente el veneno de B. alternatus con extractos de J.c o M.g, este efecto fue parcial o totalmente revertido. A su vez, el extracto de Z.m inhibió parcialmente la coagulación plasmática provocada por el veneno de B. moojeni. Estos resultados están en concordancia con otras investigaciones sobre extractos vegetales que demostraron una inhibición significativa de los efectos de venenos sobre las proteínas que coordinan la cascada de coagulación.32,33,34,35,36,37
Relacionado también con la hemostasia, es común que las víctimas de accidentes bothrópicos desarrollen trastornos hemorrágicos; pues los venenos de estas serpientes presentan enzimas proteolíticas responsables por la degradación de proteínas en la matriz extracelular, generando desestructuración en la membrana del vaso y extravasación de sangre.38 En nuestro estudio, este tipo de problema hemorrágico causado por los venenos de B. alternatus en ratones, fue inhibido completamente en la presencia de los extractos de J.c y M.g a partir de la proporción 1:10 (m:m). Asimismo, procesos hemorrágicos inducidos por venenos de serpientes clasificadas en diferentes familias, géneros o especies fueron inhibidos por diversos extractos vegetales, según diferentes trabajos científicos;23,24,25,39 lo que confirma que las plantas poseen moléculas con elevado potencial antiofídico.
Ensayos de modelaje molecular realizados por Ticli y otros (22 demostraron que el ácido rosmarínico incubado con fosfolipasas A2 aisladas de venenos de serpientes se unen al sitio activo de estas enzimas, inhibiendo sus efectos deletéreos. Observaciones similares fueron informadas por Mendes y otros20 cuando trataron una enzima metaloproteinasa de venenos de serpientes con el compuesto de origen vegetal p-cumarato de triacontilo, que inhibía completamente su actividad en ensayos in vivo e in vitro; además confirmaron, por medio de técnicas de simulación molecular, que esta molécula se unía al sitio activo de la enzima.
Los extractos acuosos de las plantas M.g y J.c fueron altamente eficaces en la neutralización de las actividades coagulante, fosfolipasa y hemorrágica inducidas por el veneno de la serpiente B. alternatus, mientras que el extracto metanólico de Z.m inhibió parcialmente las actividades fosfolipasa y coagulante inducidas por el veneno de B. moojeni. Posiblemente, estos extractos presentan metabolitos capaces de interactuar con proteínas existentes en el veneno, haciéndolas menos efectivas con respecto a su toxicidad. En el futuro, estos metabolitos podrán contribuir para el desarrollo de nuevos agentes terapéuticos para el tratamiento de accidentes ofídicos.

