










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Helicobacter pylori es una bacteria que al colonizar el epitelio gástrico induce una respuesta inmune tanto innata como adaptativa en el hospedero, siendo la primera la que juega el papel fundamental.1) Sin embargo, la persistencia de la bacteria en el estómago demuestra que esta respuesta no es efectiva en la eliminación de la infección y de manera adicional contribuye a su patogénesis.2) En las últimas décadas han surgido nuevos modelos experimentales de infección por H. pylori con el objetivo de investigar la patogénesis.3 En particular, el macrófago (MØ) constituye un modelo apropiado para las investigaciones antiinflamatorias y antioxidativas puesto que algunas bacterias poseen la habilidad de inducir la muerte de macrófagos activados a través de la estimulación de la producción de especies reactivas del oxígeno (ERO).4 Frente a la colonización por H. pylori se estimula la producción en MØ de algunas ERO como el óxido nítrico, peróxido de hidrógeno y de manera adicional citocinas proinflamatorias como la interleucina-1β.2
Debido a la alta prevalencia de infección por H. pylori a nivel mundial, los costosos tratamientos con antimicrobianos y las elevadas tasas de resistencia antimicrobiana, se han realizado esfuerzos encaminados al diseño de vacunas contra este microorganismo, sin éxito hasta este momento en humanos.5) Por lo tanto resulta esencial la búsqueda de alternativas terapéuticas, especialmente aquellas relacionadas con la nutrición, como los extractos de plantas que contienen abundantes polifenoles, los cuales han mostrado que reducen la inflamación.3) La cúrcuma, compuesto derivado del extracto de Curcuma longa L posee propiedades antiinflamatorias, antioxidantes, anticancerígena, antiproliferativa, antifúngica y antimicrobiana.4 Los estudios realizados en nuestro país con C. longa son escasos,5,6) sin embargo, uno en particular demostró propiedades antibacterianas frente a aislados autóctonos de H. pylori.7) Basado en los resultados previos, la posible variación del contenido de polifenoles de las plantas cultivadas en diferentes zonas geográficas del mundo, la presente investigación tiene como objetivo evaluar el efecto del extracto de C. longa sobre MØ peritoneales infectados con H. pylori.
Métodos
Material biológico y cultivo bacteriano
En el presente estudio experimental la recolección del rizoma de la planta C. longa y la preparación del extracto hidroalcohólico se realizó según lo descrito por Feliciano y colaboradores.8) Para los estudios en macrófagos se utilizó la cepa de referencia de H. pylori ATCC 43504, que se cultivó y re-identificó según lo descrito previamente.9)
Efecto individual y combinado del extracto de C. longa y H. pylori sobre la viabilidad celular de macrófagos peritoneales
Para la obtención de MØ peritoneales se emplearon ratones BALB/c machos de entre 30-35 g, de 8 semanas de nacidos sometidos a eutanasia por dislocación cervical.10) Los MØ se colectaron por lavado peritoneal con medio de cultivo RPMI-1640 (Sigma R8758, EE. UU.) frío (4-8 ºC) y se sembraron a una concentración de 1x105 células/mL en medio suplementado con suero fetal bovino al 10%. El extracto se disolvió en diluciones dobles seriadas hasta obtener concentraciones finales de 3,12, 6,25, 12,5, 25, 50, 100 y 200 μg/mL en el pozo. Las placas de 96 pozos se incubaron durante 24 horas. En paralelo, se utilizó un grupo de células cultivadas en medio libre de extracto de cúrcuma como control, las cuales representaron el 100 % de viabilidad. Para determinar el efecto del extracto de C. longa sobre la viabilidad de los MØ peritoneales se utilizó el ensayo de reducción del bromuro de 3-(4,5-dimetiltiazol-2-il)-2,5-difenil-2H-tetrazolio (MTT).8 Se emplearon 3 réplicas de cada concentración del extracto y se realizaron 3 ensayos. El porcentaje de viabilidad se calculó: A560 (CT)/ A560 (CNT) x100; donde CT es la célula tratada con el extracto y CNT la célula sin tratar con el extracto o el control. Adicionalmente se calcularon los valores de CI50 del extracto (concentración del extracto que reduce un 50 % la viabilidad celular).8
En la evaluación del efecto de H. pylori sobre la viabilidad de los MØ, las células se sembraron como se describió anteriormente y la bacteria se añadió a los MØ con una multiplicidad de infección (MI) de 100:1, 50:1, 25:1, 5:1, 1:1. Los cultivos se incubaron a 37 °C con 5% de CO2 durante 24 h. La viabilidad celular se determinó mediante el ensayo de Rojo neutro.11) Se realizaron tres réplicas y se incluyeron controles de crecimiento tanto para MØ como para la bacteria.
Para la evaluación del efecto combinado del extracto de C. longa y la bacteria sobre los macrófagos se utilizó el rango de concentraciones del extracto divididas en dos grupos. El primero con concentraciones del extracto por debajo de 12,5 μg/mL y el segundo por encima de 25 μg/mL con las diferentes MI evaluadas previamente. La viabilidad celular se determinó mediante el ensayo de Rojo neutro.11
Determinación del evento de apoptosis
El efecto de los tratamientos en la ocurrencia de la muerte celular por apoptosis se verificó mediante el empleo del colorante DAPI (4',6‑diamidino‑2‑fenilindol) (Sigma). La tinción se realizó según lo descrito previamente11 y el análisis de las células apoptóticas se realizó mediante el empleo de un microscopio de fluorescencia (Olympus IX-71, Japón) empleando el filtro de 480 nm.
Efecto de H. pylori sobre el estado de activación de los macrófagos peritoneales
Las bacterias a la MI evaluadas previamente se adicionaron al cultivo de macrófagos en presencia y ausencia del extracto de C. longa. A las 24 h se determinó los niveles de peróxido de hidrogeno y por otra parte, se colectó el sobrenadante de cultivo para la determinación de los niveles de óxido nítrico (ON) y citocina IL-1β.12 En cada experimento se utilizaron un grupo de células cultivadas en medio libre de tratamiento como control negativo y con LPS (Sigma) a una concentración de 10 µg/mL como control del estado de activación de MØ.
Análisis estadístico de los resultados
Los valores de CI50 del extracto se determinaron a partir del análisis de regresión sigmoidal. Para los análisis estadísticos realizados, se utilizó el programa GraphPad Prism versión 5.01 (GraphPad Software Inc., San Diego, CA). Se aplicó el método no paramétrico Kruskall-Wallis y las comparaciones múltiples se realizaron la prueba paramétrica ANOVA de dos vías (factor grupo de tratamiento) y como método de comparación múltiple la prueba de Bonferroni. Las diferencias se consideraron significativas cuando p< 0,05.
Consideraciones éticas en el uso de animales de experimentación
Este estudio fue aprobado por el Comité de ética del IPK (CE-IPK 72-18) y estuvo en concordancia con la directiva del Parlamento Europeo 2010/63/ue relativa a la protección de los animales utilizados para fines científicos.10
Resultados
El efecto del tratamiento individual y combinado del extracto de C. longa y de las multiplicidades de infección de H. pylori sobre la viabilidad de MØ posterior a las 24 h de incubación se muestra en la figura 1. Al analizar el efecto individual se obtuvo un valor de CI50 de 31± 2 para el extracto de C. longa. Solo a la concentración de 100 µg/mL disminuyó significativamente la viabilidad celular en comparación con las células no tratadas (p<0,001) (Fig. 1, A) y partir de la relación H. pylori/MØ de 25:1 con diferencias estadísticamente significativas (Fig. 1, B). La combinación de concentraciones del extracto de C. longa inferiores a 12,5 µg/mL con la relación de 25, 50 y 100 de H. pylori/MØ, mantuvo la viabilidad celular en aproximadamente el 70% (Fig. 1, C). Sin embargo, al combinar las más altas multiplicidades de H. pylori con concentraciones del extracto superiores a 25 µg/mL se observó una pérdida de la viabilidad de MØ similar al control celular inoculado con la bacteria (Fig. 1, D). Concentraciones de C. longa entre 3,12-12,5 µg/mL permitieron mantener la viabilidad de MØ frente a una alta carga bacteriana.
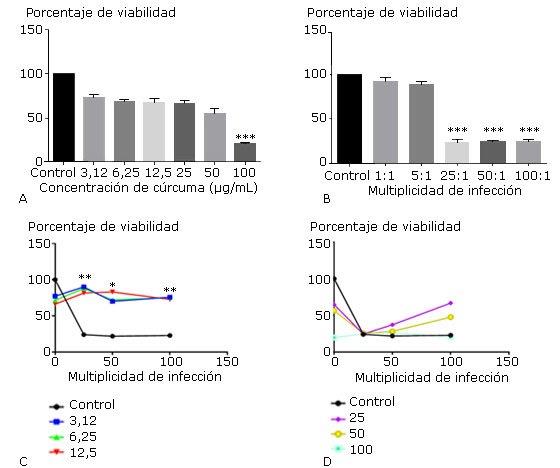 Los gráficos muestran la media ± desviación estándar del porcentaje de células viables tratadas con diferentes concentraciones del extracto de C. longa (A), diferentes multiplicidades de infección por la cepa de H. pylori ATCC 43504 (B), porcentaje de células viables tratadas con diferentes multiplicidades de infección de H. pylori ATCC 43504 y la combinación con 3,12, 6,25 y 12,5 (C), así como 25, 50 y 100 µg/mL de C. longa (D) en 24 h. Los controles representan las células con medio de cultivo (A y B) y el porcentaje de viabilidad de macrófagos cultivados con cada multiplicidad (C y D). Las diferencias se consideraron significativas cuando p< 0,05(*), p< 0,01(**) y p< 0,001(***).
Los gráficos muestran la media ± desviación estándar del porcentaje de células viables tratadas con diferentes concentraciones del extracto de C. longa (A), diferentes multiplicidades de infección por la cepa de H. pylori ATCC 43504 (B), porcentaje de células viables tratadas con diferentes multiplicidades de infección de H. pylori ATCC 43504 y la combinación con 3,12, 6,25 y 12,5 (C), así como 25, 50 y 100 µg/mL de C. longa (D) en 24 h. Los controles representan las células con medio de cultivo (A y B) y el porcentaje de viabilidad de macrófagos cultivados con cada multiplicidad (C y D). Las diferencias se consideraron significativas cuando p< 0,05(*), p< 0,01(**) y p< 0,001(***).Fig. 1 Efecto individual y combinado del extracto de Curcuma longa y de Helicobacter pylori sobre la viabilidad macrófagos peritoneales.
En la figura 2 se muestran las microfotografías de los tratamientos de macrófagos con, valores superiores e inferiores a la CI50 del extracto de C. longa, las multiplicidades de infección por H. pylori, y la combinación de ambas. A diferencia de lo que ocurre en las células no tratadas, se detectó la presencia de núcleos con la cromatina altamente condensada y la formación de cuerpos apoptóticos (flechas amarillas), estos cambios morfológicos caracterizan la muerte celular por apoptosis. Sin embargo, estas características se observan con menor intensidad cuando se incuban las células infectadas con altas multiplicidades (≥ 25) y concentraciones del extracto de cúrcuma superiores a 12,5 µg/mL.
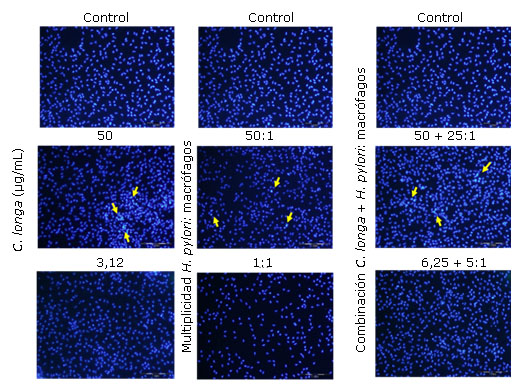 Las flechas amarillas indican la presencia de células apoptóticas. Aumento 400x.
Las flechas amarillas indican la presencia de células apoptóticas. Aumento 400x.Fig. 2 Marcaje con DAPI de los macrófagos peritoneales tratados con distintas concentraciones del extracto de Curcuma longa y multiplicidades de infección de Helicobacter pylori.
Se observó un aumento de la producción de peróxido de hidrógeno proporcional a la relación bacterias/MØ con valores estadísticamente significativos en 100 de MI (Fig. 3, A), sin embargo, el tratamiento de las células con el extracto de C. longa no estimuló la producción del metabolito cuyos valores fueron similares al del control (Fig. 3, B). No obstante, a partir de la relación 50 bacterias/MØ con la combinación de las diferentes concentraciones del extracto de C. longa se apreció una reducción en la producción de peróxido de hidrógeno celular, que resultó no estadísticamente significativa (Fig. 3, C y D).
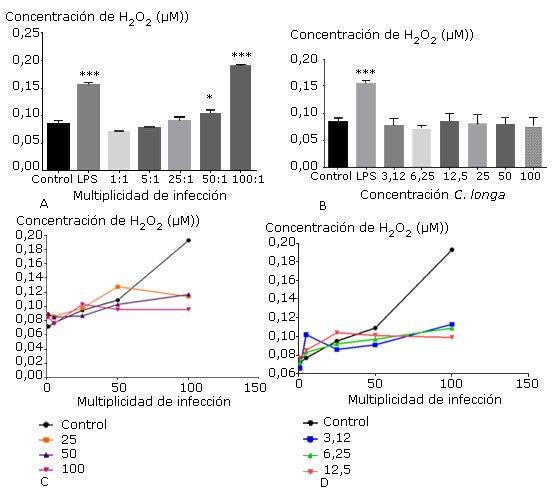 Los gráficos muestran la media ± desviación estándar del porcentaje de células viables tratadas con diferentes multiplicidades de infección por H. pylori ATCC 43504 (A) y diferentes concentraciones del extracto de C. longa (B); los gráficos C y D muestran la producción de H2O2 de las células tratadas con diferentes concentraciones del extracto de C. longa y multiplicidades de infección de H. pylori ATCC 43504. Los controles representan las células con medio de cultivo (negativo), el LPS 10 µg/mL (positivo) (A y B) y el porcentaje de viabilidad de macrófagos cultivados con cada multiplicidad (C y D). Las diferencias se consideraron significativas cuando p< 0,05(*) y p< 0,001(***).
Los gráficos muestran la media ± desviación estándar del porcentaje de células viables tratadas con diferentes multiplicidades de infección por H. pylori ATCC 43504 (A) y diferentes concentraciones del extracto de C. longa (B); los gráficos C y D muestran la producción de H2O2 de las células tratadas con diferentes concentraciones del extracto de C. longa y multiplicidades de infección de H. pylori ATCC 43504. Los controles representan las células con medio de cultivo (negativo), el LPS 10 µg/mL (positivo) (A y B) y el porcentaje de viabilidad de macrófagos cultivados con cada multiplicidad (C y D). Las diferencias se consideraron significativas cuando p< 0,05(*) y p< 0,001(***).Fig. 3 Efecto individual y combinado del extracto de C. longa y de H. pylori sobre la producción de peróxido de hidrógeno (H2O2) en macrófagos peritoneales.
La producción de óxido nítrico resultó ser elevada frente a concentraciones del extracto inferiores a 12,5 µg/mL y superiores a 25 µg/mL (Fig. 4, A) así como a MI superiores a 5 bacterias/MØ (Fig. 4, B). Al analizar las combinaciones, la producción de óxido nítrico se comportó de manera diferente pues se constató la disminución de su producción en un rango de concentración del extracto de 6,25-50 µg/mL para todas multiplicidades de infección evaluadas, siendo estadísticamente significativo en la relación de 100 bacterias/MØ (Fig. 4, C y D).
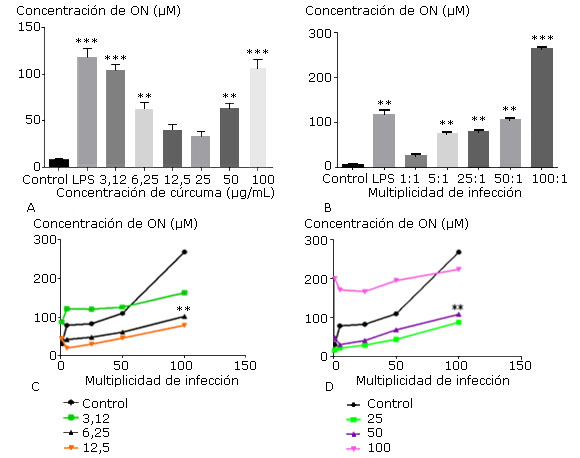 Los gráficos muestran la media ± desviación estándar del porcentaje de células viables de las células tratadas con diferentes concentraciones del extracto de C. longa (A) y diferentes multiplicidades de infección por H. pylori ATCC 43504 (B); los gráficos C y D muestran la producción de ON en las células tratadas con diferentes concentraciones del extracto de C. longa y multiplicidades de infección de H. pylori ATCC 43504. Los controles representan las células con medio de cultivo (negativo), el LPS 10 µg/mL (positivo) (A y B) y el porcentaje de viabilidad de macrófagos cultivados con cada multiplicidad (C y D). Las diferencias se consideraron significativas cuando p< 0,01(**) y p< 0,001(***).
Los gráficos muestran la media ± desviación estándar del porcentaje de células viables de las células tratadas con diferentes concentraciones del extracto de C. longa (A) y diferentes multiplicidades de infección por H. pylori ATCC 43504 (B); los gráficos C y D muestran la producción de ON en las células tratadas con diferentes concentraciones del extracto de C. longa y multiplicidades de infección de H. pylori ATCC 43504. Los controles representan las células con medio de cultivo (negativo), el LPS 10 µg/mL (positivo) (A y B) y el porcentaje de viabilidad de macrófagos cultivados con cada multiplicidad (C y D). Las diferencias se consideraron significativas cuando p< 0,01(**) y p< 0,001(***).Fig. 4 Efecto individual y combinado del extracto de C. longa y de H. pylori sobre la producción de óxido nítrico (ON) en macrófagos peritoneales.
Como se muestra en la figura 5, la producción de IL-1β en el sobrenadante de cultivo celular aumentó en relación con las diferentes MI no siendo así para el tratamiento con cúrcuma (Fig. 5, A y B). El efecto combinado de las diferentes concentraciones de C. longa y las MI denotó una disminución en la producción de IL-1β (Fig. 5, C y D).
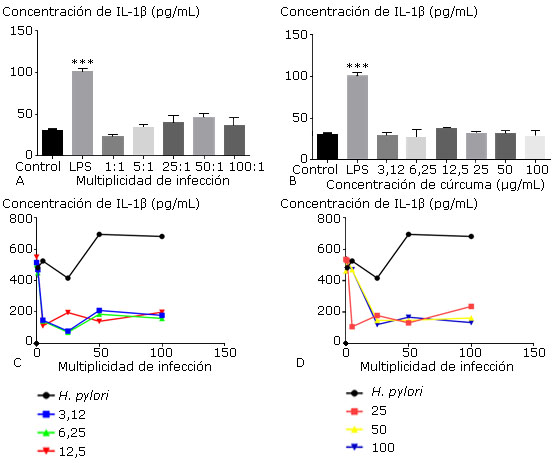 Los gráficos muestran la media ± desviación estándar de la concentración de interleucina IL-1β en las células cultivadas con diferentes multiplicidades de infección de H. pylori ATCC 43504 (A) y con diferentes concentraciones del extracto de C. longa (B); los gráficos C y D muestran el efecto combinado de H. pylori y de C. longa sobre la producción de interleucina IL-1β. Los controles representan las células con medio de cultivo (negativo), el LPS 10 µg/mL (positivo) (A y B) y el porcentaje de viabilidad de macrófagos cultivados con cada multiplicidad (C y D). No se observaron diferencias estadísticamente significativas.
Los gráficos muestran la media ± desviación estándar de la concentración de interleucina IL-1β en las células cultivadas con diferentes multiplicidades de infección de H. pylori ATCC 43504 (A) y con diferentes concentraciones del extracto de C. longa (B); los gráficos C y D muestran el efecto combinado de H. pylori y de C. longa sobre la producción de interleucina IL-1β. Los controles representan las células con medio de cultivo (negativo), el LPS 10 µg/mL (positivo) (A y B) y el porcentaje de viabilidad de macrófagos cultivados con cada multiplicidad (C y D). No se observaron diferencias estadísticamente significativas.Fig. 5 Efecto individual y combinado del extracto de C. longa y de H. pylori sobre la producción de citocina IL-1β en sobrenadante de cultivo de macrófagos peritoneales.
Discusión
En la presente investigación bajas multiplicidades de infección no afectan la viabilidad de los macrófagos. Este comportamiento resulta similar a lo observado por Lin y colaboradores13) con la infección por H. pylori a una MI baja de la línea de macrófago murino RAW264.7 que provoca la supresión de la expresión de genes que codifican la síntesis de ADN y de moléculas asociadas al ciclo celular, sin afectar la viabilidad celular.
Según reporta la literatura,14 la curcumina reduce significativamente las moléculas coestimulantes y también inhibe la activación de MAPK y la translocación de NF-κB p65, mecanismos que explican la recuperación de la viabilidad del macrófago al combinar la bacteria a diferentes multiplicidades de infección con concentraciones del extracto inferiores a 12,5 µg/mL, en el presente estudio. Los resultados obtenidos coinciden con el reporte de autores que señalan la existencia de muerte celular abrupta de monocitos y macrófagos causada por altas multiplicidades de infección por H. pylori.14,15 Dichos reportes indican que este fenómeno ocurre a través de la activación de las vías Erk, arginasa II o la vía dependiente de mitocondria. También se ha reconocido la activación de la muerte celular mediante el mecanismo de autofagia la cual se caracteriza por la formación de vesículas "autofagosomas", que envuelven proteínas desnaturalizadas, orgánulos celulares, agregados de proteínas desintegrados y dañados.(15 El contenido de autofagosomas es posteriormente vertido en lisosomas para su posterior degradación enzimática.13
La medición de la apoptosis se realiza frecuentemente mediante microscopía de fluorescencia, que muestra imágenes de cuerpos apoptóticos y condensación de cromatina versus células de control no apoptóticas.15 Estudios recientes muestran buenos efectos en la prevención de apoptosis con el empleo de bajas dosis de curcumina.16,17 La proporción de apoptosis temprana en los grupos tratados con dosis medias y altas de curcumina aumentó significativamente, mientras que la proporción de apoptosis tardía disminuyó de manera significativa. El efecto de la curcumina para prevenir la transformación de los estadios de la apoptosis de tardía a temprana necesita aún ser investigada.
El exceso de ERO bajo estrés oxidativo reacciona biológicamente con moléculas activas las cuales pueden causar daño oxidativo a las células y producir cáncer e inflamación crónica. El mantenimiento y mejoramiento del estado oxidativo, especialmente a través del uso de productos naturales, son esenciales para el funcionamiento de los procesos fisiológicos en animales.18
La curcumina, componente activo principal del extracto de C. longa, estructuralmente contiene grupos hidroxilos en la posición orto de anillos aromáticos que participan en la regulación de las enzimas de la fase 2 de detoxificación. Un estudio muestra que 10 μM de curcumina puede disminuir los niveles de ERO en macrófagos peritoneales de ratas.18 Otro estudio observa un efecto dual de la curcumina sobre las ERO.19 Dosis medias y bajas de curcumina tienen un efecto contundente en la eliminación de ERO mientras altas dosis de curcumina intensifican la producción de ERO. Esto se debe a que la curcumina incrementa la actividad de las enzimas catalasa, superóxido dismutasa y glutatión peroxidasa, responsables de la disminución de los niveles celulares de las ERO.
Una posible explicación a la disminución de los niveles de producción de peróxido de hidrógeno y óxido nítrico con todas las concentraciones de extracto de C. longa excepto 25 µg/mL puede deberse a que la curcumina inhibe la activación del canal TRPM2 (del inglés, Transient receptor potential melastatin 2). Este es un canal no selectivo, permeable a cationes que se activa de manera directa mediante la ribosa adenosina difosfato e indirectamente bajo condiciones de estrés oxidativo por la vía de formación de H2O2.20 Este canal responde frente a las ERO secretadas en sitios inflamatorios mediante la secreción de citocinas y quimoquinas esenciales en el control de la inflamación, especialmente en tejidos mucosales. El impacto del canal TRPM2 en la producción de ERO y citocinas inflamatorias pudiera propiciar la colonización gástrica exitosa de la bacteria y el daño epitelial.
La apoptosis de macrófagos puede ser inducida por numerosas moléculas efectoras, incluyendo el óxido nítrico. Estudios in vivo demuestran que la mayor expresión y actividad de la óxido nítrico sintasa ocurre en macrófagos, fuente fundamental de la enzima, por lo tanto la regulación de su producción modula los niveles de apoptosis de estas células.19 La enzima inducible óxido nítrico sintasa y la generación de óxido nítrico son factores clave en la muerte de H. pylori en el interior del fagocito. La arginasa bacteriana, codificada por el gen rocF, es utilizada por H. pylori como estrategia para disminuir la disponibilidad de L-arginina, que es necesaria para la producción de óxido nítrico y por ende, regular su efecto antibacteriano.18
La IL-1β se genera y libera rápidamente por los macrófagos en respuesta a señales inflamatorias y puede actuar como un amplificador de la señal. Adicionalmente, esta molécula es necesaria para el inicio tanto de la respuesta innata como la adaptativa mediante el reclutamiento de células defensoras al sitio de infección, y el paso de leucocitos del torrente circulatorio al tejido para eliminar la bacteria. Es importante destacar que, la pérdida de macrófagos activados es similar a la disminución de la respuesta inmunológica frente al patógeno20) lo que contribuiría a la imposibilidad del hospedero de eliminar la infección. Afortunadamente, el extracto de C. longa puede estimular la defensa celular a través de la producción de IL-1β y al mismo tiempo, aumentar la actividad fagocítica de los macrófagos.19,21 Sería útil en próximos estudios evaluar otras citocinas inflamatorias y antiinflamatorias.
Los ensayos preclínicos son fundamentales para la posible identificación y luego utilización de compuestos activos derivados de productos herbarios. La literatura internacional aboga por la necesidad de ensayos clínicos más controlados con estos productos.2C. longa es uno de los compuestos que han sido evaluados de forma combinada con los antimicrobianos, lo que ha mejorado el estrés oxidativo y los cambios histológicos en la gastritis asociada a H. pylori.18) En Cuba son escasos los estudios que evalúan las potencialidades medicinales de la planta C. longa L. y hasta donde conocemos, este constituye el primer reporte que evalúa su efecto antiinflamatorio y antioxidante.
Conclusiones
El tratamiento de los macrófagos peritoneales infectados por H. pylori con el extracto de C. longa provocó una disminución de la citotoxicidad celular, hecho que resulta positivo para una mejor respuesta inmune innata. La disminución de la producción de especies reactivas del oxígeno y de los niveles de producción de IL-1β en macrófagos peritoneales frente al tratamiento con el extracto de cúrcuma sugiere su uso como producto antioxidante y antiinflamatorio que propicia una mayor respuesta inmunológica frente a los mecanismos de persistencia de la infección por H. pylori. Sería útil en estudios futuros determinar otros eventos de muerte celular y evaluar la secreción de otras citocinas en macrófagos peritoneales.

