










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Hematología, Inmunología y Hemoterapia
versión On-line ISSN 1561-2996
Rev Cubana Hematol Inmunol Hemoter v.21 n.2 Ciudad de la Habana Mayo-ago. 2005
Universidad de Antioquia, Colombia
Importancia de los ratones knock-out en el estudio del sistema NADPH oxidasa
Est. Julián Camilo Arango Rincón, Dr. Andrés Augusto Arias Sierra y Dr. Pablo Javier Patiño
Resumen
El anión superóxido producido por el sistema NADPH oxidasa de las células fagocíticas es parte de las especies reactivas del oxígeno (ROS) involucradas en la defensa contra microorganismos patógenos. Su importancia se evidencia en la enfermedad granulomatosa crónica (EGC), una inmunodeficiencia primaria ocasionada por mutaciones en los genes que codifican las proteínas del sistema NADPH oxidasa. Los ratones knock-out para algunas de las proteínas del sistema NADPH oxidasa han permitido comprender la función de las ROS en la destrucción de microorganismos patógenos, la regulación del proceso inflamatorio y la fisiología del sistema NADPH oxidasa. Se espera que esta clase de modelos animales proporcionen avances en la comprensión de los eventos comprendidos en la terapia génica para la EGC, de los nuevos paradigmas acerca del papel de las ROS en la defensa del hospedero y en la descripción de las vías de señalización comprometidas con la activación de este sistema.
Palabras clave: sistema NADPH oxidasa, enfermedad granulomatosa crónica, ratones knock-out, terapia génica, especies reactivas del oxígeno (ROS).
Una de las principales formas de usar modelos murinos para el estudio de los genes es la técnica knock-out, la cual consiste en suprimir selectivamente un gen de interés, originando de esta manera alteraciones fisiológicas que ofrecen información con respecto a la función in vivo del gen en estudio. Basados en este principio, en el ratón se pueden generar deficiencias inmunológicas congénitas debidas a alteraciones en uno o varios genes; este es el caso de la enfermedad granulomatosa (EGC), donde el uso de ratones knock-out ha permitido comprender el papel de los genes y proteínas del sistema NADPH oxidasa.2, 3
El presente trabajo describe el sistema NADPH oxidasa como el mecanismo inmune alterado en la EGC, la metodología necesaria para la creación de ratones knock-out y los hallazgos importantes que se han logrado gracias a este modelo murino en el entendimiento del sistema NADPH oxidasa.
SISTEMA NADPH OXIDASA Y ENFERMEDAD GRANULOMATOSA CRÓNICA
El sistema NADPH oxidasa es un complejo enzimático presente en células fagocíticas como neutrófilos, eosinófilos, monocitos y macrófagos y en células no fagocíticas como linfocitos B, linfocitos T, fibroblastos, células de músculo liso, endoteliales, del cuerpo carotídeo, de pulmón, de riñón, e inclusive en células vegetales.4,5 Este sistema se encuentra conformado por 5 subunidades proteicas, 3 de ellas (p40phox, p47phox y p67phox) se encuentran en el citoplasma formando un complejo macromolecular, mientras que el resto (gp91phox y p22phox) están localizadas en la membrana de los fagolisosomas y la membrana plasmática, donde constituyen un heterodímero llamado flavocitocromo b558.5 En el funcionamiento del sistema NADPH oxidasa intervienen otras proteínas como Rac1 y Rac2.6 La activación del mismo da comienzo al fenómeno llamado “estallido respiratorio”, que se caracteriza por la liberación del anión superóxido (O-2) y otras especies reactivas del oxígeno (ROS) como el peróxido de hidrógeno (H2O2), el radical hidroxilo (OH-) y ácidos hipohalógenos en la vacuola fagocítica o en el medio extracelular, lo cual induce la muerte de microorganismos fagocitados.5, 7
La EGC es una inmunodeficiencia primaria 8-10 en la que los individuos afectados sufren infecciones crónicas y granulomas inflamatorios causados por microorganismos como Staphylococcus aureus, Salmonella sp, Klebsiela sp, Aerobacter sp, Serratia sp y microorganismos oportunistas. Los signos característicos de la EGC son la presencia de abscesos en la piel, osteomielitis de huesos cortos, otitis media, colitis, enteritis, linfoadenitis, microabscesos hepáticos, así como obstrucción de las vías urinarias y gástrica causadas por granulomas.9,11 La causa de este síndrome es la presencia de mutaciones en cualquiera de los genes que codifican las proteínas del complejo enzimático NADPH oxidasa, excepto en el gen de p40 phox.9, 11 La forma más frecuente de EGC es la causada por mutaciones en el gen de la gp91phox (X-EGC), la cual tiene un patrón hereditario ligado al cromosoma X.12,13 Las demás formas de EGC son de tipo autosómicas recesivas debido a mutaciones en los genes de p22phox, p47phox y p67phox.9, 14, 15 Mutaciones en Rac 2 han sido asociadas con defectos en el estallido respiratorio de las células fagocíticas e infecciones recurrentes.16,17
El manejo inicial de los pacientes con EGC consiste en evitar las infecciones a repetición e indicar un tratamiento agresivo con antibióticos cuando estas aparecen. De la misma manera, el tratamiento profiláctico con interferón ? (IFN-?) puede reducir la frecuencia de infecciones en estos pacientes.12,18 La única alternativa de tratamiento curativo para la EGC es el trasplante de médula ósea, y actualmente se trabaja en la terapia génica. Sin embargo, esta alternativa terapéutica aún se encuentra en fase experimental.3,12
ESTUDIO DE GENES POR MEDIO DE ANIMALES KNOCK-OUT
Durante gran parte del siglo pasado, el análisis genético se realizaba usando modelos animales que poseían mutaciones naturales; tales modelos presentaban fenotipos alterados, los cuales permitieron dilucidar los mecanismos genéticos y moleculares de varios defectos génicos.19,20 A finales del siglo XX, con el auge de las técnicas de biología molecular, se comenzó a estudiar la función del los genes partiendo de las secuencias genómicas hasta llegar a su respectivo fenotipo, proceso que tomó el nombre de “genética inversa” y que se ha valido de las técnicas de manipulación génica para adicionar, inactivar e incrementar la expresión de ciertos genes en células, tejidos y organismos completos, para luego evaluar los efectos de dichas alteraciones y así entender el papel que desempeñan los genes involucrados.2,19,20 La técnica de los knock-out consiste en reprimir o inactivar la función específica de un gen remplazándolo por formas mutadas de este en estadios celulares muy primitivos. 2,21 En el caso de la EGC, se aprovecha el mecanismo de la recombinación homóloga para inactivar algunos de los genes importantes del sistema NADPH oxidasa generando un modelo murino knock-out para esta enfermedad.
Para la creación de ratones knock-out (tabla 1), 2,19-21 inicialmente se diseña un vector que posee una copia modificada del gen que se desea inactivar, y genes reporteros que permitan confirmar el éxito en el proceso de incorporación del vector a la célula madre embrionaria (CME) y la recombinación homóloga de este. Los genes reporteros más utilizados son el gen de resistencia a la neomicina (neor) y el gen de la timidina quinasa del virus herpes simple (TkHSV).19,21 Con este vector se procede a realizar una transferencia génica a un grupo de CME de ratón, generalmente de la cepa 129, las cuales previamente fueron aisladas de la capa más interna del blastocisto conocida como blastocele.19,21,22 En estas células, el establecimiento del knock-out se logra por medio del fenómeno de recombinación homóloga, el cual consiste en el intercambio de secuencias similares entre cromátidas no hermanas de un par de cromosomas homólogos por la acción conjunta de un grupo de enzimas denominadas recombinasas. Por lo tanto, la eliminación de las secuencias de interés depende de que las recombinasas faciliten la recombinación homóloga entre el vector y secuencias cromosómicas en las CME. 2,20
Tabla1. Herramientas necesarias para inactivar de forma selectiva un gen (crear knock-out).

Con el propósito de confirmar la incorporación del vector a las CME, estas son cultivadas en un medio que permite una primera selección (selección positiva), la cual ocurre gracias a la presencia en el vector de un gen de resistencia a un antibiótico, por ejemplo a la neomicina, como se mencionó antes. Posteriormente, para corroborar el éxito de la recombinación homóloga, se realiza una segunda selección (selección negativa), mediante la adición al medio de cultivo de un análogo de nucleósido como el ganciclovir, que impide la replicación del DNA en las células que han incorporado a su genoma el gen TkHSV por procesos de recombinación no homóloga o heteróloga.19,21 La recombinación homóloga también se confirma por medio de técnicas como la PCR y el Southern blot.21,23 Después de la selección de las CME que poseen el gen inactivado mediante recombinación homóloga, estas se implantan en un blastocisto que carece de CME; por lo general se utilizan ratones de la cepa C57BL/6; a su vez este blastocisto es implantado en una madre sustituta pseudopreñada de la misma cepa. Este procedimiento genera un ratón quimera (129-C57BL/6), del cual se recupera el fondo genético de la cepa C57BL/6 por medio de cruces consanguíneos o retrocruces (figura). 22, 24

Figura. Procesos involucrados en la generación de ratones knock-out para las proteínas del sistema NADPH oxidasa y sus aplicaciones .
A: el vector con el gen knock-out es transfectado en células madre embrionarias (CME) de ratón cepa 129. B: las CME son cultivadas con neomicina (G418), para seleccionar las células que incorporaron el vector (selección positiva), luego el cultivo es expuesto al ganciclovir, el cual selecciona las células que realizaron una recombinación homologa (selección negativa). C: las CME knock-out son implantadas en un blastocisto de ratón cepa C57BL6 y este a su vez es implantado en una madre pseudopreñada C57BL6; la descendencia corresponde a ratones quiméricos 129/C57BL6; mediante retrocruces se recupera el fondo genético de la cepa C 57BL6. D: los ratones knock-out modelo de enfermedad granulomatosa crónica son empleados en diferentes tipos de estudios.
Ratones knock-out como una herramienta para el estudio del sistema NADPH oxidasa
El conocimiento sobre la función y la interacción entre los componentes del sistema NADPH oxidasa nos permite entender los mecanismos moleculares implicados en la producción de las especies reactivas del oxígeno y la generación de estrategias relacionadas con la modulación positiva o negativa de la producción de estos agentes reactivos. Estudios previos del ensamble y activación del sistema NADPH oxidasa, han usado células hematopoyéticas humanas y murinas, o líneas celulares hematopoyéticas inmortalizadas. 25-27 Los neutrófilos no son fáciles de modificar genéticamente y las líneas inmortalizadas tienen limitaciones de manipulación, transfección y expresión a largo plazo de proteínas heterólogas. Otros grupos como el nuestro, han utilizado sistemas no hematopoyéticos basado en células COS para determinar el efecto de mutaciones en los genes que codifican para las proteínas del sistema NADPH oxidasa en la producción de especies reactivas del oxígeno (ROS). 28-30 Por otro lado, los sistemas libres de células ofrecen flexibilidad para el uso de proteínas recombinantes. 31,32
Sin embargo, existen diferencias entre la activación en un sistema reconstituido libre de células comparado con el ambiente fisiológico que se da en los sistemas donde se usan células completas. De la misma manera, la EGC es una inmunodeficiencia primaria poco frecuente, lo que dificulta el estudio de su patogénesis y sus características moleculares. Teniendo en cuenta esto, resulta de gran utilidad los modelos experimentales in vivo, que además de responder a esta necesidad, permitan comprender los procesos bioquímicos y moleculares asociados con el sistema NADPH oxidasa. Para tales propósitos se dispone de los ratones knock-out para gp91phox p47phox y Rac 2. 3,33-35
Ratones knock-out para gp91phox (gp91phoxKO)
Pollock y colaboradores en 1995 crearon el primer modelo de enfermedad granulomatosa crónica ligada al cromosoma X (EGC-X) al generar ratones knock-out para gp91phox. Este tipo de ratones ha sido útil para evaluar el papel que poseen las ROS en infecciones crónicas, en la regulación del proceso inflamatorio y como modelo experimental para la terapia génica en la EGC-X.3
Cuando los ratones gp91phoxKO son infectados por vía aérea con Aspergillus fumigatus, desarrollan necrosis peribronquial y neumonía alveolar caracterizada por un infiltrado inflamatorio denso con presencia de hifas, abscesos en hígado, riñón y hueso; algunos de estos ratones mueren a las 72 horas después de la inoculación. A diferencia de los ratones gp91phoxKO, los ratones tipo silvestre infectados con este mismo microorganismo sufren de neumonía peribronquial, que se resuelve 7 días posinfección. El cuadro clínico del ratón gp91phoxKO mencionado inicialmente es similar al que suelen presentar los humanos con EGC-X, caracterizado por neumonías graves, abscesos en pulmón, nódulos linfáticos, hígado y hueso.3
Con el fin de evaluar la regulación del proceso inflamatorio en la EGC-X, los ratones gp91phoxKO se inocularon intraperitonealmente con tioglicolato de sodio. Esta sustancia irritante permite evaluar la quimiotaxis de neutrófilos a un determinado sitio sin necesidad de causar una infección con microorganismos. Como resultado, se evidenció una peritonitis crónica y un infiltrado de neutrófilos de 2 a 3 veces mayor en los ratones gp91phoxKO comparado con ratones tipo silvestre a las 18 y 24 horas posinoculación. Este hallazgo demuestra que los mecanismos dependientes del oxígeno, además de mediar un papel importante en la destrucción de microorganismos como A. fumigatus, están involucrados en la regulación del proceso inflamatorio aún cuando el microorganismo ya haya sido eliminado.36
Una característica importante de los ratones gp91phoxKO consiste en que no sufren infecciones espontáneas por microorganismos propios de su flora normal; algo similar ocurre con los ratones knock-out para la enzima óxido nítrico-sintasa (NOS2). Estos hallazgos sugieren la existencia de otros mecanismos microbicidas que compensan la deficiencia de las ROS y las especies reactivas del nitrógeno (RNS) en la respuesta contra infecciones por flora normal.37 Para comprobar esta hipótesis, Shiloh y colaboradores crearon un ratón doblemente deficiente para gp91phox y NOS2. Después de estar en condición libre de microorganismos externos, estos ratones desarrollaron infecciones espontáneas por microorganismos de la flora normal del tracto digestivo, mientras que los ratones deficientes para una sola proteína, gp91phox o NOS2, no sufrieron ningún tipo de infección bajo las mismas condiciones. Esto demuestra que en el modelo murino las ROS y RNS son esenciales para el control de la flora normal. Sin embrago, existe redundancia entre ambos mecanismos inmunes en etapas tempranas de la respuesta inmune innata contra estos microorganismos.37
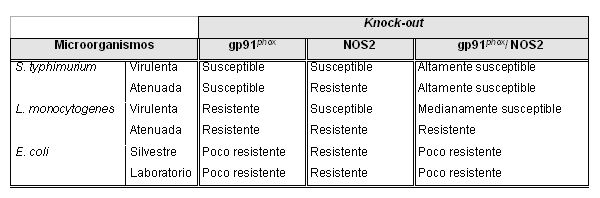
De igual manera se evaluó la susceptibilidad in vivo de los knock-out doblemente deficientes a cepas virulentas y no virulentas de Salmonella typhimurium, Listeria monocytogenes y Escherichia coli. Este estudio demostró que las ROS y RNS atacaban microorganismos de forma específica (tabla 2); por ejemplo, las ROS tuvieron actividad antimicrobicida ante S. typhimurium, mientras que las RNS participaron en la resistencia contra L. monocytogenes. Adicionalmente se comprobó que las ROS se consumían más rápido, entre 1 y 3 horas, mientras que las RNS tardaban entre 1 y 3 días. Con este y otros resultados se ha determinado que p47phox y gp91phox son esenciales en la respuesta a S. typhimurium, A. fumigatus y S. aureus, mientras que la NOS2 es vital para la protección contra microorganismos como Mycobacterium tuberculosis, Leishmania major, Leismania donovani, que tienen como blanco los fagosomas de las células fagocíticas.37, 38
Tabla 2. Susceptibilidad de ratones gp91phoxKO, NOS2KO y gp91phox/NOS2 KO a S. typhimurium, E. coli y L. monocytogenes.
Ratones knock-out para p47phox (p47phoxKO)
En 1995 Jackson y colaboradores crearon el primer ratón knock-out deficiente de la proteína p47phox.33 Este modelo animal de EGC ha posibilitado el estudio del proceso inflamatorio, de la formación de granulomas en esta enfermedad y la relación del sistema NADPH oxidasa con enfermedades como aterosclerosis, carcinogénesis, daños por isquemia-reperfusión y la formación de cataratas.33
Se ha reportado que los ratones p47phoxKO desarrollan un infiltrado de leucocitos 2 veces mayor al de los ratones normales heterocigotos, demostrando una falta de modulación del proceso inflamatorio durante la respuesta anti-infecciosa.36
Inicialmente, se pensó que esta desregulación se debía a una infección persistente donde el microorganismo patógeno sobrevive y continúa estimulando la migración de neutrófilos y macrófagos al sitio de la infección por la carencia de ROS. 36
Esta hipótesis no era del todo satisfactoria para explicar la desregulación, ya que los granulomas de algunos pacientes con EGC eran estériles y que además respondían a tratamiento con esteroides sin antibiótico, lo cual indicaba que la inflamación persistente causada por el microorganismo no era el principal factor del estímulo inflamatorio.36 Algunos investigadores postularon que este proceso se debía a una respuesta inmune mediada por citocinas producidas por linfocitos T disfuncionales. Esta nueva hipótesis se evaluó usando ratones p47phoxKO y silvestres, en los cuales se estudió la producción de citocinas Th1 y Th2 frente a infecciones con Mycobacterium avium y Schistosoma mansoni, respectivamente. Los resultados demostraron que tanto el ratón silvestre como el p47phoxKO mostraban un patrón de citocinas similar en cada uno de los estímulos. Estos resultados sugieren que las citocinas producidas por linfocitos T, son normales en individuos con EGC, descartando la posibilidad antes mencionada. Entonces la explicación a este fenómeno se fundamentó en la capacidad que poseen las ROS para degradar factores proinflamatorios, de modo que ante la disminución o ausencia de ROS, las citocinas y factores quimiotácticos responsables del proceso inflamatorio se acumulan, provocando la formación de granulomas y la inflamación persistente. Dicha teoría es respaldada por ensayos in vitro en los cuales estos oxidantes pueden inactivar factores proinflamatorios como IL-8, leucotrienos, la fracción C5a del complemento y los péptidos N-formilados.
De esta forma, se reafirmó que las ROS, además de destruir los microorganismos fagocitados, tienen un importante papel en la regulación del proceso inflamatorio.36
Los pacientes con EGC presentan una alta susceptibilidad a bacterias y hongos productores de catalasa, a diferencia de las infecciones por microorganismos catalasa negativos. Usando el modelo knock-out para p47phox, Chang y colaboradores evaluaron la capacidad de virulencia de los microorganismos catalasa negativos frente a los catalasa positivos empleando cepas mutantes de Aspergillus nidulans que carecían de catalasa A, B o ambas. Con A. nidulans silvestre el resultado fue una infección fatal con ambas cepas de hongos en los ratones knock-out, mientras que los ratones tipo silvestre resolvieron fácilmente la infección por cualquiera de estos microorganismos. Este experimento demostró que las catalasas de A. nidulans no influyen en la patogénesis y severidad de las lesiones sobre ratones con EGC, a diferencia de lo que se presenta en pacientes con EGC, donde aún no está claro por qué son más frecuentes las infecciones con microorganismos catalasa positivos.39
Otro aspecto que se ha evaluado con el uso de ratones p47phoxKO, es la función que tienen las ROS en la infección por M. tuberculosis, en los cuales se observó que en etapas tempranas de la infección había un mayor crecimiento bacteriano comparado con los ratones tipo silvestre; además, los ratones p47phoxKO presentaban un incremento de neutrófilos en el sitio de inflamación y la aparición de granulomas. Estos hallazgos han corroborado la importancia de las ROS en las etapas iniciales de una infección causada por M. tuberculosis al menos en los ratones.40
En el año 2000, Hsich y colaboradores buscando establecer alguna relación entre la producción de ROS por células endoteliales y las lesiones causadas por aterosclerosis, crearon un ratón knock-out doblemente mutado para apoE (modelo murino de aterosclerosis) y p47phox, en los que se evaluó la producción basal O2- en los anillos aórticos tanto de los ratones silvestres como los mutados.41 Como resultado, se encontró una baja producción de este radical sin diferencia significativa en los niveles de ambos tipos de ratones; de la misma forma, al evaluar la patología y fisiología vascular, tampoco se encontraron diferencias significativas en el tamaño de las lesiones aórticas y en los niveles de colesterol. Estos resultados demuestran que existe una pequeña actividad basal del sistema NADPH oxidasa en el tejido vascular y que posiblemente las lesiones causadas por aterosclerosis se deban a la acción de otros mecanismos diferentes a una alta producción de ROS en células endoteliales.41
Estudios realizados por Reeves y colaboradores empleando ratones knock-out para p47phox y ensayos físico-químicos, han permitido proponer un papel secundario de las ROS en la eliminación de los microorganismos. Esta nueva teoría se basa en que el O2- activa señales para la entrada de iones K+ a la vacuola fagocítica, lo que a su vez permite la generación de un medio hipertónico y básico en el fagolisosoma; esta condición permite la solubilización y activación de proteínas catiónicas y líticas como la catepsina G y la elastasa, que son las principales mediadoras en la destrucción de los microorganismos.42
Ratones knock-out para Rac2 (Rac2KO)
Rac2 es una GTPasa necesaria en la activación del sistema NADPH oxidasa; su importancia en la activación de este sistema y su papel en otros procesos fisiológicos de las células fagocíticas ha sido demostrado con el empleo de ratones Rac2KO.6, 34, 35 Los neutrófilos extraidos de ratones deficientes para esta proteína, presentaron defectos en la quimiotaxis al ser puestos en contacto con sustancias quimoatrayentes como el formil-metionil-leucil-fenilalanina (fMLP), IL-8 y leucotrieno B4. Adicionalmente presentaron defectos en la interacción con moléculas del endotelio especialmente con la L-selectina y el Glycam1, así como deficiencias en la polimerización de actina y en la activación de las proteínas quinasas activadas por mitógenos (MAPK): p42, p44, ERK½ y p38 en respuesta a quimoatrayentes. 6,35 Estos hallazgos son de gran interés, ya que neutrófilos aislados de pacientes con una mutación dominante negativa de Rac2 presentan características similares.16, 17
La actividad del sistema NADPH oxidasa en neutrófilos de médula ósea preactivados con TNF-a y estimulados con formol-meristato-acetato (PMA), se encontró significativamente disminuida cuando se utilizaron ratones Rac2KO en comparación con el control de tipo silvestre bajo las mismas condiciones. Sin embargo, en neutrófilos Rac2KO de exudado peritoneal estimulados con tioglicolato de sodio la producción de O2- fue similar a los controles silvestres. De la misma manera se encontró que estos ratones presentaban una disminución de la inflamación, defectos en la quimiotaxis, leucocitosis y neutrofilia, y un incremento en la mortalidad al ser infectados con A. fumigatus, lo cual demostraba que aunque estos ratones producían ROS, eran sensibles a la infección por A. Fumigatus posiblemente por una disminución de la migración celular al sitio de infección, lo que conlleva a una reducción de la inflamación y a neutrofilia en el torrente sanguíneo.35
Li y colaboradores evaluaron la quimiotaxis y la producción de O2- en neutrófilos provenientes de ratones Rac2KO-/-, Rac2KO-/+ (heterocigotos), y de ratones silvestres. En neutrófilos obtenidos de ratones Rac2-/- se observó una baja producción de anión superóxido y defectos en la respuesta quimiotáctica al compararse con ratones tipo silvestre ante diferentes estímulos como el fMLP; asimismo, neutrófilos procedentes de ratones Rac2KO-/+ presentan una actividad intermedia entre los ratones knock-out para Rac 2 y ratones tipo silvestre. 34 Estos resultados demuestran la importancia de Rac2 y la falta de redundancia con otras moléculas en funciones tales como la movilidad celular y la quimiotaxis; asimismo se comprobó que Rac2 lleva a cabo estas funciones de una forma “dependiente de dosis-génica”. 34
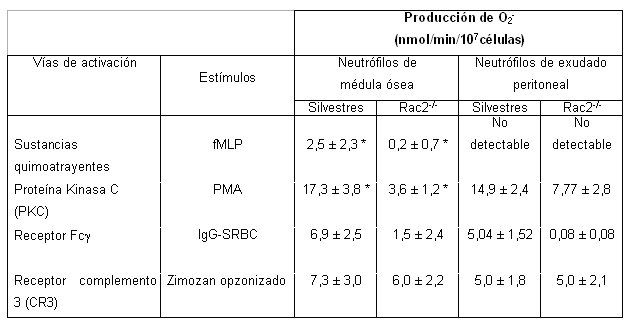
También Kim y colaborares investigaron en qué vías de activación, y ante qué clase de estímulos era esencial la presencia de Rac2. Al evaluar las principales vías para la activación del sistema NADPH oxidasa en ratones Rac2KO, se evidenciaron deficiencias en 3 de ellas: la del receptor Fc?, la de los receptores de sustancias quimoatrayentes y la vía de la proteína quinasa C (PKC) inducida por PMA (tabla 3). Estos resultados reflejan la importancia de Rac2 en la activación de estas vías. Por su parte, la vía de del receptor del complemento 3 (CR3), no resultó alterada, lo cual indica que hay otros mecanismos que compensan la ausencia de Rac2, o que Rac2 no es esencial para el funcionamiento de esta vía de activación. En la tabla 3 se puede observar que ante el estímulo con PMA en neutrófilos de ratones Rac2KO, había una gran disminución en la producción de O2-. Esto significa que Rac2 está relacionada con la fosforilación de quinasas corriente abajo de la PKC, necesarias para la activación del sistema NADPH oxidasa.6
Tabla 3. Importancia de Rac 2 en diferentes vías de activación
Los diversos hallazgos en los ratones knock-out para Rac 2 han ayudado a comprender su papel en la fisiología de las células fagocíticas. Ahora se sabe que Rac2 es una molécula necesaria para la producción de anión superóxido solo en determinadas vías, especialmente en respuesta al receptor Fc?. 6 Sin embargo, en situaciones en las que no se altera la actividad del sistema NADPH oxidasa o esta es llevada a cabo parcialmente, se cree que existen otras moléculas que compensen la función de Rac2, por ejemplo Rac1, la cual es una proteína que se expresa en todas la células y posee una homología en la secuencia de aminoácidos con Rac2 del 92 %. 6,34,35
Terapia génica en ratones knock-out
Los ratones knock-out para p47phox, gp91phox y Rac2, además de ser modelos para entender la fisiopatología de la EGC y el funcionamiento básico del sistema NADPH oxidasa, han sido importantes en el desarrollo de terapias para la EGC. Una de ellas es la terapia génica, la cual consiste en el remplazo del material genético alterado en las células de los pacientes con esta enfermedad. En estudios en los cuales se han empleado ratones knock-out deficientes de gp91phox como modelos de terapia génica, se ha obtenido una corrección parcial de la actividad oxidasa entre el 50 % - 80 % de los neutrófilos de torrente sanguíneo, con una duración hasta de 18 meses. 43,44 En el caso de ratones p47phoxKO, se ha observado que entre el 8,4 y 17,3 % de los neutrófilos de torrente sanguíneo, presentan actividad oxidasa, que perdura hasta un mes después de la transferencia génica; después de 3 meses el porcentaje se reduce entre el 2,6 y el 10 %.45 Al realizar pruebas funcionales en estos ratones se observó que 2 de cada 9 ratones manipulados con terapia génica para p47phox resolvían una infección con B. cepacia, 45 mientras que en los ratones con terapia génica para gp91phox se evitaba el desarrollo de neumonía por A. fumigatus. 43
Los ensayos clínicos de terapia génica de fase I en humanos no han sido exitosos, ya que los pacientes X-EGC solo presentan del 0,004 al 0,05 % de neutrófilos circulantes con actividad oxidasa.46 Algo similar sucedió con la terapia génica para p47phox, en la cual los pacientes deficientes de esta proteína presentaron del 0,06 al 0,2 % de neutrófilos circulantes con actividad oxidasa.46 Estos resultados preliminares indican que aún son necesarios más estudios en esta área, y que tal vez con el mejoramiento de las técnicas de transferencia génica será posible lograr un tratamiento adecuado para los pacientes con EGC. Los ratones knock-out han sido útiles para evaluar diversos aspectos relacionados con la terapia génica, tales como efectos adversos, consecuencias de los métodos de reimplante de células progenitoras, efectos de la expresión constitutiva de transgenes en diferentes tejidos, el efecto fisiológico del transgen y su grado de expresión.47 Sin embargo, estos resultados no son totalmente aplicables en humanos, ya que existen múltiples diferencias entre los modelos murinos y el humano en lo referente a células fagocíticas. Por ejemplo, en el humano hay un mayor porcentaje de neutrófilos en el torrente sanguíneo que en el ratón, igualmente las vías de señalización por receptores tipo Toll y la producción de óxido nítrico por macrófagos es diferente entre los ratones y los humanos. Como estas, existen otras diferencias en la inmunidad innata y adquirida que demuestran la importancia de manejar estas discrepancias a la hora de interpretar y aplicar los resultados preliminares de la terapia génica obtenida en ratones.48
En resumen, se puede concluir que los ratones knock-out para p47phox, gp91phox y Rac2 han resultado ser modelos animales de gran utilidad en la comprensión de la fisiología de las células fagocíticas, del sistema NADPH oxidasa y de los mecanismos patogénicos de la EGC. Igualmente, estos modelos animales han permitido avances significativos en la búsqueda de un tratamiento definitivo para esta entidad basado en terapia génica.
En el futuro se espera que esta clase de modelos animales proporcionen avances importantes en campos diferentes. El primero es el entendimiento de algunos eventos involucrados en la terapia génica, luego está la evaluación de nuevos paradigmas en cuanto al papel de las ROS en la defensa del hospedero contra microorganismos, y por último, la descripción de las vías de señalización, en las que están incluidas las proteínas del sistema NADPH oxidasa y las Rac-GTPasas.
Summary
Importance of the knock out mice in the study of the nadph oxidase system
The superoxide anion produced by the NADPH oxidase system of the phagocytic cells is part of the reactive oxygen species involved in the defense against pathogenic microorganisms. Its importance is evidenced in the chronic granulomatous disease (CGD), a primary immunodeficiency caused by mutations in the genes that code the proteins of the NADPH oxidase system. The knock-out mice for some of the proteins of the NADPH oxidase system have allowed to understand the function of the ROS in the destruction of pathogenic microorganisms, the regulation of the inflammatory process, and the physiology of the NADPH oxidase system, It is expected that this class of animal models makes possible to attain advances in the understanding of the events included in the genic therapy for the CGD, of the paradigms about the role of the ROS in the defense of the host, and in the description of the signaling pathways involved in the activation of this system.
Key words: NADPH oxidase system, chronic granulomatous disease, knock-out mice, genic therapy, reactive oxygen species (ROS).
Referencias bibliográficas
1. Benavides F, Guenet JL. Murine models for human diseases. Medicina (B Aires) 2001;61(2):215-31.
2. Lodish H, Berk A, Zipursky SL, Matsudaira P, Baltimore D, Darnell J. Molecular cell biology. W. H. Freeman and Company; 1999.
3. Pollock JD, Williams DA, Gifford MA, Li LL, Du X, Fisherman J, et al. Mouse model of X-linked chronic granulomatous disease, an inherited defect in phagocyte superoxide production. Nat Genet 1995;9(2):202-9.
4. Jackson SH, Devadas S, Kwon J, Pinto LA, Williams MS. T cells express a phagocyte-type NADPH oxidase that is activated after T cell receptor stimulation. Nat Immunol 2004;5(8):818-27.
5. Vignais PV. The superoxide-generating NADPH oxidase: Structural aspects and activation mechanism. Cell Mol Life Sci 2002;59(9):1428-59.
6. Kim C, Dinauer MC. Rac2 is an essential regulator of neutrophil nicotinamide adenine dinucleotide phosphate oxidase activation in response to specific signaling pathways. J Immunol 2001;166(2):1223-32.
7. Klebanoff S. Inflammation: Basic principles and clinical correlates. En: Gallin JI, Snyderman R, eds. 3 ed; 1999.
8. Segal BH, Leto TL, Gallin JI, Malech HL, Holland SM. Genetic, biochemical, and clinical features of chronic granulomatous disease. Medicine (Baltimore) 2000;79(3):170-200.
9. Heyworth PG, Cross AR, Curnutte JT. Chronic granulomatous disease. Curr Opin Immunol 2003;15(5):578-84.
10. Montoya CJ, Henao J, Salgado H, Olivares MM, López JA, Rugeles C, et al. Phenotypic diagnosis of primary immunodeficiencies in Antioquia, Colombia, 1994-2002. Biomedica 2002;22(4):510-8.
11. Zambrano E, Esper F, Rosenberg R, Kim J, Reyes-Mugica M. Chronic Granulomatous disease. Pediatr Dev Pathol 2003;6(6).
12. Dinauer MC, Lekstrom-Himes JA, Dale DC. Inherited neutrophil disorders: Molecular basis and new therapies. Hematology (Am Soc Hematol Educ Program) 2000:303-318.
13. Patino PJ, Perez JE, López JA, Condino-Neto A, Grumach AS, Botero JH, et al. Molecular analysis of chronic granulomatous disease caused by defects in gp91-phox. Hum Mutat 1999;13(1):29-37.
14. Cornejo De Luigi M, Lopez JA, Navarro S, García D, Patino PJ. Clinical and molecular characterization of autosomal recessive chronic granulomatous disease caused by p47-phox deficiency. Rev Med Chil 2000;128(5):491-8.
15. Patino PJ, Rae J, Noack D, Erickson R, Ding J, de Olarte DG, et al. Molecular characterization of autosomal recessive chronic granulomatous disease caused by a defect of the nicotinamide adenine dinucleotide phosphate (reduced form) oxidase component p67-phox. Blood 1999;94(7):2505-14.
16. Williams DA, Tao W, Yang F, Kim C, Gu Y, Mansfield P, et al. Dominant negative mutation of the hematopoietic-specific Rho GTPase, Rac2, is associated with a human phagocyte immunodeficiency. Blood 2000;96(5):1646-54.
17. Ambruso DR, Knall C, Abell AN, Panepinto J, Kurkchubasche A, Thurman G, et al. Human neutrophil immunodeficiency syndrome is associated with an inhibitory Rac2 mutation. Proc Natl Acad Sci USA 2000;97(9):4654-9.
18. Condino-Neto A, Newburger PE. Interferon-gamma improves splicing efficiency of CYBB gene transcripts in an interferon-responsive variant of chronic granulomatous disease due to a splice site consensus region mutation. Blood 2000;95(11):3548-54.
19. Strachan T, Read AP. Human molecular genetics. 2 ed. Bios Scientific Publishers Ltd; 1999.
20. Griffiths AJF, Miller JH, Suzuki DT, Lewontin RC, Gelbart WM. An introduction to genetic analysis. 7 ed. W. H. Freeman; 2000.
21. Watson JD, Gilman M, Witkowski J, Zoller M. Recombinant DNA. 2 ed. Scientific American Books; 1992.
22. Seong E, Saunders TL, Stewart CL, Burmeister M. To knockout in 129 or in C57BL/6: that is the question. Trends Genet 2004;20(2):59-62.
23. Capecchi MR. Targeted gene replacement. Sci Am 1994;270(3):52-9.
24. Wong GT. Speed congenics: applications for transgenic and knock-out mouse strains. Neuropeptides 2002;36(2-3):230-6.
25. De Mendez I, Adams AG, Sokolic RA, Malech HL, Leto TL. Multiple SH3 domain interactions regulate NADPH oxidase assembly in whole cells. Embo J 1996;15(6):1211-20.
26. Dusi S, Nadalini KA, Donini M, Zentilin L, Wientjes FB, Roos D, et al. Nicotinamide-adenine dinucleotide phosphate oxidase assembly and activation in EBV-transformed B lymphoblastoid cell lines of normal and chronic granulomatous disease patients. J Immunol 1998;161(9):4968-74.
27. Paclet MH, Coleman AW, Burritt J, Morel F. NADPH oxidase of Epstein-Barr-virus immortalized B lymphocytes. Effect of cytochrome b(558) glycosylation. Eur J Biochem 2001;268(19):5197-208.
28. Arias AA, Matute JD, Jiabin D, Patiño PJ, Dinauer MC. Expresión y actividad de posibles polimorfismos provenientes de individuos normales en la proteína de 67 kd del sistema NADPH oxidasa utilizando el sistema COSphox. Biomedica 2004;24(3):262-72.
29. -----------. Molecular analysis of mutant forms of p67-phox using the COS-phox system. En: Hematology ASo, ed. 44th Annual Meeting American Society of Hematology, 2002. Philadelphia, Pennsylvania: Blood; 2002. p. 458a.
30. Price MO, McPhail LC, Lambeth JD, Han CH, Knaus UG, Dinauer MC. Creation of a genetic system for analysis of the phagocyte respiratory burst: high-level reconstitution of the NADPH oxidase in a nonhematopoietic system. Blood 2002;99(8):2653-61.
31. Cross AR, Erickson RW, Ellis BA, Curnutte JT. Spontaneous activation of NADPH oxidase in a cell-free system: Unexpected multiple effects of magnesium ion concentrations. Biochem J 1999;338 ( Pt 1):229-33.
32. Peng G, Huang J, Boyd M, Kleinberg ME. Properties of phagocyte NADPH oxidase p47-phox mutants with unmasked SH3 (Src homology 3) domains: Full reconstitution of oxidase activity in a semi-recombinant cell-free system lacking arachidonic acid. Biochem J 2003;373(Pt 1):221-9.
33. Jackson SH, Gallin JI, Holland SM. The p47phox mouse knock-out model of chronic granulomatous disease. J Exp Med 1995;182(3):751-8.
34. Li S, Yamauchi A, Marchal CC, Molitoris JK, Quilliam LA, Dinauer MC. Chemoattractant-stimulated Rac activation in wild-type and Rac2-deficient murine neutrophils: Preferential activation of Rac2 and Rac2 gene dosage effect on neutrophil functions. J Immunol 2002;169(9):5043-51.
35. Roberts AW, Kim C, Zhen L, Lowe JB, Kapur R, Petryniak B, et al. Deficiency of the hematopoietic cell-specific Rho family GTPase Rac2 is characterized by abnormalities in neutrophil function and host defense. Immunity 1999;10(2):183-96.
36. Segal BH, Doherty TM, Wynn TA, Cheever AW, Sher A, Holland SM. The p47(phox-/-) mouse model of chronic granulomatous disease has normal granuloma formation and cytokine responses to Mycobacterium avium and Schistosoma mansoni eggs. Infect Immun 1999;67(4):1659-65.
37. Shiloh MU, MacMicking JD, Nicholson S, Brause JE, Potter S, Marino M, et al. Phenotype of mice and macrophages deficient in both phagocyte oxidase and inducible nitric oxide synthase. Immunity 1999;10(1):29-38.
38. Nathan C, Shiloh MU. Reactive oxygen and nitrogen intermediates in the relationship between mammalian hosts and microbial pathogens. Proc Natl Acad Sci USA 2000;97(16):8841-8.
39. Chang YC, Segal BH, Holland SM, Miller GF, Kwon-Chung KJ. Virulence of catalase-deficient aspergillus nidulans in p47(phox)-/- mice. Implications for fungal pathogenicity and host defense in chronic granulomatous disease. J Clin Invest 1998;101(9):1843-50.
40. Cooper AM, Segal BH, Frank AA, Holland SM, Orme IM. Transient loss of resistance to pulmonary tuberculosis in p47(phox-/-) mice. Infect Immun 2000;68(3):1231-4.
41. Hsich E, Segal BH, Pagano PJ, Rey FE, Paigen B, Deleonardis J, et al. Vascular effects following homozygous disruption of p47(phox) : An essential component of NADPH oxidase. Circulation 2000;101(11):1234-6.
42. Reeves EP, Lu H, Jacobs HL, Messina CG, Bolsover S, Gabella G, et al. Killing activity of neutrophils is mediated through activation of proteases by K+ flux. Nature 2002;416(6878):291-7.
43. Bjorgvinsdottir H, Ding C, Pech N, Gifford MA, Li LL, Dinauer MC. Retroviral-mediated gene transfer of gp91phox into bone marrow cells rescues defect in host defense against Aspergillus fumigatus in murine X-linked chronic granulomatous disease. Blood 1997;89(1):41-8.
44. Dinauer MC, Li LL, Bjorgvinsdottir H, Ding C, Pech N. Long-term correction of phagocyte NADPH oxidase activity by retroviral-mediated gene transfer in murine X-linked chronic granulomatous disease. Blood 1999;94(3):914-22.
45. Mardiney M, Jackson SH, Spratt SK, Li F, Holland SM, Malech HL. Enhanced host defense after gene transfer in the murine p47phox-deficient model of chronic granulomatous disease. Blood 1997;89(7):2268-75.
46. Malech HL, Maples PB, Whiting-Theobald N, Linton GF, Sekhsaria S, Vowells SJ, et al. Prolonged production of NADPH oxidase-corrected granulocytes after gene therapy of chronic granulomatous disease. Proc Natl Acad Sci U S A 1997;94(22):12133-8.
47. Goebel WS, Kume A, Dinauer MC. Gene therapy for choronic granulomatous disease and leukocyte adhesion deficiency. Intern J Ped Hematol Oncol 2001;7(5-6):359-372.
48. Mestas J, Hughes CC. Of mice and not men: Differences between mouse and human immunology. J Immunol 2004;172(5):2731-8.
Recibido: 29 de julio de 2005. Aprobado: 18 de agosto de 2005
Julián Camilo Arango Rincón. Street 62 # 52-59 (SIU) Lab: 530. Tel: 210 6475. Universidad de Antioquia. e-mail: mjcar610@udea.edu.co; arjulian2@hotmail.com
http://medicina.udea.edu.co/Investigacion/Grupos/inmunopri/index.htm

