










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Hematología, Inmunología y Hemoterapia
versión impresa ISSN 0864-0289
Rev Cubana Hematol Inmunol Hemoter vol.29 no.4 Ciudad de la Habana oct.-dic. 2013
ARTÍCULO DE REVISIÓN
Aspectos actuales de la organogénesis. Función e involución del timo
Present aspects of organogenesis. Function and thymic involution
Dra. Vianed Marsán Suárez, Lic. Lázaro O del Valle Pérez, DraC. Consuelo Macías Abraham
Instituto de Hematología e Inmunología. La Habana, Cuba.
RESUMEN
El timo es un órgano linfoide central o primario localizado en la parte anterosuperior del tórax. Constituye el principal sitio de diferenciación y maduración de las células T. En esta revisión se detallan aspectos actuales de la embriología, la histología y la función tímica y en la generación de los diferentes subtipos de timocitos y su diferenciación a células T maduras efectoras, la inducción de las células T tímicas reguladoras involucradas en el mantenimiento de la tolerancia a lo propio y la involución que sufre este órgano durante el proceso de inmunosenescencia.
Palabras clave: organogénesis tímica, células T, selección positiva, selección negativa, involución tímica
ABSTRACT
Thymus is a primary organ located in the antesuperior area of the torax. It is the principal place of differentiation and maduration of T cells. In this review present aspects of the embriology, histology and thymic function are detailed, as well as its role in the generation of different kinds of thymic cells; its differentiation to mature cells and of regulator T cells has a crucial role in tolerance induction. Moreover, thymic involution during of immunosenescence process is shown.
Keywords: thymic organogenesis, T cells, positive selection, negative selection, thymic involution.
INTRODUCCIÓN
Según la etimología, el término timo deriva de la palabra griega «thymos», que significa alma o espíritu. 1
Galeno (siglo II d. de Cristo) atribuyó su función a la purificación del sistema nervioso. Versalio (siglo XV) teorizó que el timo actuaba como amortiguador para proteger los importantes vasos del mediastino que se encontraban detrás del esternón. En el siglo XVIII se entendía que el timo regulaba de alguna manera la función pulmonar fetal y neonatal y este era conocido como el órgano vicario de la respiración.2
En 1777, Guillermo Hewson fue el primero en identificar correctamente al timo como una glándula que modifica la linfa. En 1832, Sir Astley Cooper describió la anatomía de este órgano a través de disecciones detalladas de cadáveres.
Hassall y Vanarsdale, en 1846, emplearon el microscopio compuesto para estudiar la histología del timo y describieron las diferencias de este con otros órganos linfoides, específicamente la existencia en él de los corpúsculos de Hassall.2
Sin embargo, fue el inmunólogo australiano Miller, en 1961, quien al demostrar el efecto devastador de la timectomía en el sistema inmunitario, aportó una noción clara de cuál era su verdadera función.3
EMBRIOLOGÍA
El timo se origina de la superficie ventrolateral y de la porción ventral de la tercera y cuarta bolsas faríngeas, respectivamente; elementos derivados de las tres capas germinales.4
Su desarrollo comienza en la sexta semana de gestación. En esta etapa, la bolsa faríngea se divide en dos porciones: una dorsal, que da origen a las glándulas paratiroides inferiores; y una ventral de donde se deriva el primordio tímico.4
En la séptima semana, cada primordio tímico migra de manera caudal y medial, desde el ángulo de la mandíbula hasta el mediastino anterosuperior, formando una estructura tubular llamada tracto o ductus timofaríngeo. Este tracto comienza en el seno piriforme, perfora la membrana tirohioidea y emerge entre la arteria carótida común y el nervio vago. Cursa posterior al nervio glosofaríngeo y lateral a la glándula tiroides y entra al mediastino.5
Hacia la octava semana, los primordios tímicos se fusionan en la línea y descienden, adoptando su posición característica en el mediastino anterosuperior; la porción cefálica, usualmente involuciona.4,5
Alrededor de la novena semana está constituido solo por células epiteliales y no es hasta la décima semana que es invadido por pequeñas células linfoides que migran desde el timo fetal y la médula ósea (MO), formando el tejido linfoide tímico. La diferenciación celular se completa entre las catorce y dieciséis semanas de gestación.2,4,5
El timo crece rápidamente y alcanza un gran peso antes del nacimiento, en relación con el peso corporal. Está ubicado en la parte superior del mediastino anterior y se apoya sobre el pericardio, al nivel del nacimiento de los grandes vasos. 5
HISTOLOGÍA
El timo es un órgano grueso y bilobulado, rodeado de una cápsula de tejido conectivo laxo. Cada lóbulo está dividido en lobulillos, por tabiques fibrosos y organizados en dos compartimentos: corteza y médula. 2,5-7
La corteza está compuesta primariamente de linfocitos pequeños íntimamente empaquetados (timocitos) y esparcidas entre estos, escasas células más voluminosas: epiteliales (corticales) y mesenquimales. 5-7
La médula está constituida por un gran número de células epiteliales (medulares) y pocos linfocitos pequeños. Las células epiteliales componen el esqueleto o armazón del timo, son llamadas células cuidadoras (del inglés «nurse cells») y son funcionalmente esenciales para la maduración de los timocitos. La médula tímica posee, además, nidos de células epiteliales maduras queratinizantes denominados corpúsculos de Hassall. También contiene células dendríticas, macrófagos y células miodes, que aparentemente degeneran y desaparecen durante el desarrollo fetal, pero constituyen una posible fuente de antígenos musculares y son de gran interés por su participación en la patogenia de la miastenia gravis. 6-9
El timo presenta un gran número de vasos sanguíneos flexibles y de linfáticos eferentes, que drenan a los nódulos linfoides mediastinales y carece de folículos linfoides que aparecen cuando existe una hiperactividad tímica. 8,9
Se han descrito dos modelos de desarrollo de las células epiteliales tímicas. El primero se refiere a que células progenitoras endodérmicas dan origen a una célula progenitora tímica o «stem cell» y que ésta, a su vez, origina dos células progenitoras epiteliales: cortical y medular, respectivamente, que dan lugar a estos dos tipos celulares epiteliales. El segundo modelo propone que la célula progenitora endodérmica origina directamente a los progenitores epiteliales cortical y medular. 10 (Figura 1).
En este proceso de organogénesis del timo participan diferentes genes que codifican para factores transcripcionales, entre ellos: Hoxa3, Pax1, Pax9, Eya1 y Foxn1. Estos productos génicos son indispensables para desencadenar una cascada de eventos que incluyen: iniciación, posición, crecimiento, separación y diferenciación del tejido tímico. 8-10
EL TIMO EN LA MADURACIÓN DE LAS CÉLULAS T
El timo es el principal sitio de maduración de las células T. Esta función se definió por el hecho de que algunas deficiencias inmunológicas en el humano estaban asociadas a su ausencia o a su escaso desarrollo. Experimentos realizados en ratones neonatos a los cuales se les extirpó el timo, demostraron la ausencia de células T maduras. 9,10
Las células T se originan a partir de una célula madre pluripotencial derivada de la MO, que migra al timo y una vez allí, atraviesa por diferentes estados de diferenciación. Las células epiteliales y mesenquimales tímicas, los macrófagos y las células dendríticas proveen a los timocitos de los estímulos necesarios para su atracción, proliferación, expansión, migración, diferenciación y maduración. Las células estromales tímicas, que incluyen a las epiteliales, secretan citocinas, entre ellas, interleucina-7 (IL-7), que estimulan la proliferación de las células T inmaduras. 9-11
Dos tipos de receptores de quimocinas CCR7 y CCR9, realizan una importante función en el asentamiento de progenitores en el timo, CCR7 y su ligando CCL25, que son expresados tempranamente en el estroma tímico. Los defectos en la expresión de la molécula CCR9 fueron encontrados en el ratón Rag-/-, donde existe una pobre reconstitución del timo. Se demostró que CCR9 emite señales indispensables para aumentar la afinidad de la expresión de otras moléculas que median el arresto celular al endotelio. 11-13
Dos tipos de moléculas producidas por las células tímicas no linfoides son también muy importantes para la proliferación y maduración de los timocitos: las moléculas del sistema principal de histocompatibilidad (SPH) y las citosinas. 14
En la corteza tímica tiene lugar un alto grado de proliferación celular y de muerte celular pro apoptosis; el 95 % de las células mueren por apoptosis antes de llegar a la médula. Esta muerte programada se debe a una combinación de defectos en la expresión de receptores de antígenos funcionales. 15
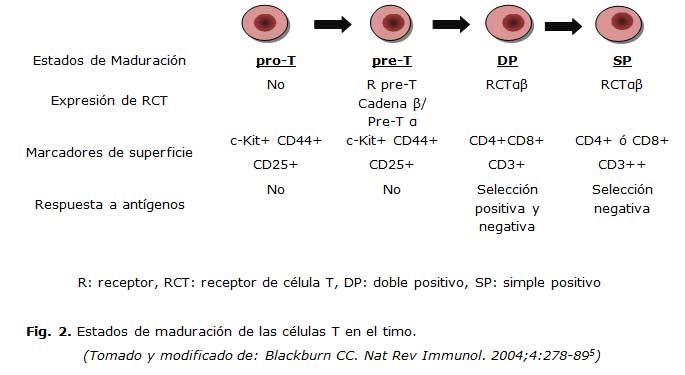
Diferentes eventos secuenciales tienen lugar durante este proceso de maduración celular, que incluyen: la recombinación somática y la expresión de los genes para los receptores de células T. Diversas moléculas celulares son expresadas sobre los timocitos en maduración, entre las que se encuentran: c-Kit, CD44, CD25, CD4, CD8 y CD3, las que definen los diferentes estados de maduración de las células T: pro-T, pre-T o doble negativa (DN) CD4-/CD8-, cortical o doble positiva (DP) CD4+/CD8+, T inmadura o simple positiva (SP) CD4+ o CD8+ y T madura que emerge a la sangre periférica y posteriormente va a poblar los órganos linfoides periféricos. 14,15 (Figura 2).
Los subtipos DN se dividen además en 4 estadios: DN1-3 y DN 4/pre-DP, de acuerdo a con la expresión diferenciada de CD44 y CD25. 16,17
La expresión del receptor de célula T (RCT), heterodímero áâ, tiene lugar en la población de timocitos DP, justo antes o durante la migración de los timocitos desde la corteza hacia la médula y es precedido de la inducción de Foxp3. 16
En el timo tienen lugar dos importantes eventos de selección: positiva y negativa. 17
La selección positiva es el proceso en el cual los timocitos cuyos RCT que se unen con una baja afinidad a las moléculas del SPH expresadas sobre las células epiteliales, son rescatadas de la muerte celular programada y estimuladas a sobrevivir. Aquellos timocitos con RCT que no reconocen a la molécula del SPH propia, mueren por apoptosis. 16,17
Este tipo de selección promueve la supervivencia y la expansión de los timocitos DP y su conversión a SP, según el tipo de molécula del SPH que ha contactado con ellos; si es de clase I, se diferencian a timocitos CD8 positivos y si es de clase II, a timocitos CD4 positivos. 17
La selección negativa es el proceso en el cual los timocitos cuyos RCT reconocen con alta afinidad el complejo SPH-péptido propio, son delecionados y programados a morir. De esta manera, se elimina el desarrollo de células T potencialmente autorreactivas contra antígenos propios que se encuentran en altas concentraciones en el timo. Este proceso es denominado deleción clonal y da lugar a la tolerancia central. 15-17
Subtipo de timocitos con RCT ãæ
Los timocitos que expresan RCT áâ y ãæ derivan de un precursor común pero constituyen dos linajes diferentes de timocitos. En el timo fetal el primer reordenamiento de genes para el RCT involucra al RCT ãæ. Esto sugiere que las células autorreactivas que superan el umbral de afinidad para la selección negativa estarían sometidas a un proceso de selección adicional al final de su maduración. Las células T que expresan las cadenas ãæ funcionales no expresan RCT áâ y viceversa, fenómeno conocido como exclusión alélica. El porcentaje de células T ã æ varía en diferentes tejidos y especies. Alrededor del 5 % del total de células T en el humano expresan este tipo de RCT. En los linfocitos T intraepiteliales intestinales representan alrededor del 10 %. 18
Las células T con RCT ãæ no reconocen péptidos asociados a las moléculas del SPH y sí pequeñas moléculas fosforiladas o lipídicas que pueden estar presentadas por moléculas del SPH de clase I no clásicas, comúnmente encontradas en micobacterias y otros microbios. Otros clones reconocen proteínas o antígenos no proteicos que no requieren procesamiento por parte de las células presentadoras de antígeno para su posterior presentación. La diversidad limitada de las células T ãæ en muchos tejidos sugiere que los ligandos de estos RCT son invariantes y conservados.14,18
Células T tímicas reguladoras
Recientemente se han encontrado en el timo humano fetal y postnatal, dos pequeños subtipos celulares que expresan moléculas CD4+/CD25+ y CD8+/CD25, indistintamente. Estas células presentan además, Foxp3 y GITR mRNA, así como también las moléculas de membrana CCR8 y TNFR2 y la proteína citoplasmática CTLA-4, todas encontradas en las células T maduras reguladoras. 19
Estudios funcionales realizados en timocitos CD4+CD25+ expandidos por una citocina similar a la IL-7, conocida como linfopoyetina estromal tímica (TSLP) derivada de las células epiteliales tímicas, muestran una baja proliferación de estas células frente al estímulo, expresan el factor de transcripción asociado a las células T reguladoras Foxp3 e inhiben la capacidad proliferativa de las células CD4+CD25- estimuladas con anticuerpos anti-CD3 y anti-CD28. TSLP constituye un activador potente de las células dendríticas que se encuentran en el timo. La inducción de las células T reguladoras por TSLP es dependiente de la interacción con moléculas de clase II del SPH, CD80, CD86 y de la presencia de citocinas como la IL-2. 19-24
Los análisis hechos sobre células T vírgenes de la periferia, expandidas con TSLP-CD, muestran que a diferencia de los timocitos, las células periféricas no se diferencian a un fenotipo regulador CD4+ CD25+ Foxp3+, lo cual indica la importancia del microambiente tímico en esta diferenciación celular. 20-24
Al estimular in vitro estas células T tímicas reguladoras, no proliferan ni producen citocinas, pero expresan las moléculas de superficie CTLA-4 y TGF-ß1 y suprimen la proliferación de timocitos autólogos CD4+CD25- tras la estimulación alogénica, por un mecanismo dependiente de contactos celulares relacionados con la acción combinada de CTA-4 y TGF-ß que conducen a la inhibición de la expresión de la cadena á del receptor de interleucina 2 sobre las células T. 20,21
Ambos subtipos de células T reguladoras ejercen una fuerte actividad supresora sobre la población de células Th1 y menor sobre la Th2. Estas células son responsables de mantener la tolerancia a lo propio y prevenir desórdenes autoinmunes. 19-22
INVOLUCIÓN TÍMICA
La involución que sufre el timo en los humanos se inicia durante la pubertad y se completa al final de la sexta década de la vida, de modo que en la edad adulta gran parte del parénquima tímico ha sido reemplazado por grasa. 25
En este proceso disminuye su tamaño, peso y actividad como resultado de la influencia que ejercen sobre él los altos niveles de hormonas sexuales circulantes durante la pubertad, el bajo número de células precursoras derivadas de la MO y los cambios que sufre el microambiente tímico. 26
Este fenómeno tiene implicaciones en el mantenimiento del repertorio de linfocitos T vírgenes, lo cual provoca numerosos defectos funcionales, incluido el acortamiento de telómeros, un repertorio restringido de RCT, poca producción de IL-2 y deficiencias en su diferenciación y proliferación hacia células efectoras. 25,26
Se han descrito diversos cambios asociados con la involución tímica. Entre los más estudiados, se encuentra: la reducción en el número total de linfocitos T vírgenes que expresan el fenotipo CD45RA+, CD62L+ CD27+, CD28+ y CD11a+, por disminución en la timopoyesis; y el incremento en el número de linfocitos T de memoria, como una consecuencia de la experiencia inmunológica que se adquiere durante la vida. El efecto de una reducción de linfocitos T vírgenes en sangre periférica es el empobrecimiento en el repertorio total de linfocitos T, lo cual puede llevar a una limitada respuesta hacia los nuevos antígenos. 27
Las células que recién emigran del timo, pueden ser identificadas por la presencia de pequeños fragmentos de ADN que permanecen durante el reordenamiento del RCT, los cuales se denominan círculos de excisión de señales de unión del RCT. La generación de linfocitos T que expresan estos círculos, permanece relativamente constante a lo largo de la vida y persiste en individuos mayores de 50 años, aun después de haber recibido un trasplante de MO. 26,27
Experimentos de trasplante de tejido tímico y de MO procedentes de ratones jóvenes a ratones viejos, demuestran que la maduración de los protimocitos a linfocitos T está severamente comprometida a partir de los dos meses de edad del ratón. Por otro lado, el número de protimocitos en la médula no parece variar con la edad, aunque sí sufren un proceso madurativo diferente. Así, el conjunto de precursores DN, que expresan CD44 pero no CD25, no está alterado con la edad. Sin embargo, su progenie inmediata (CD44+/CD25+, CD44-/CD25+, CD44-/CD25) sufre una declinación con el envejecimiento. Estos datos muestran cómo la maduración tímica en edades avanzadas parece estar bloqueada en las primeras etapas, justo en el momento previo al reordenamiento del gen Vß del RCT, en los constituyentes de la línea germinal. 25-28
El envejecimiento que sufre el sistema inmunitario genera cambios en el repertorio de los linfocitos T. Diferentes subpoblaciones de linfocitos T modifican su fenotipo durante el envejecimiento, lo que incrementa la proporción de linfocitos T de memoria. Este cambio se presenta tanto en linfocitos T CD4+ como en linfocitos T CD8+ .25,26
La expresión de marcadores de memoria CD44 y CD62L (L-selectina) también puede ser modificada. Aunque los linfocitos T con fenotipo de memoria se incrementan con la edad, existe poca evidencia que sustente que su función esté disminuida. Uno de los cambios cualitativos más relevantes existentes en la población de linfocitos T de memoria es la aparición de múltiples expansiones clonales dentro de los linfocitos T CD8+, así como la pérdida de moléculas coestimuladoras como CD40L y CD28, con un espectro de especificidad altamente restringido, de tal forma que las células T deterioradas manifiestan poca citotoxicidad y un desbalance en la producción de citocinas, lo que genera condiciones proinflamatorias. 27-29
La involución del timo con la consiguiente disminución de las células T cooperadoras, afecta además la función y la diversidad de las células B, que conduce a una débil y deficiente respuesta de anticuerpos y un incremento en la producción de autoanticuerpos. 26,28
La atrofia del timo y la disminución de su actividad constituyen procesos naturales que suceden a lo largo de la vida, los cuales provocan alteraciones en las respuestas de células T y B, que dan como resultado un aumento en la susceptibilidad para adquirir infecciones debido a la disminución en la capacidad para eliminar diferentes patógenos, la disminución de la memoria inmunológica y un alto riesgo de padecer enfermedades autoinmunes y cáncer. 25-29
REFERENCIAS BIBLIOGRÁFICAS
1. Jacobs MT, Frush DP, Donnetty LF. The right place at the wrong time: historical perspective of the thymus gland and pediatric radiology. Radiology. 1999;210:11-6.
2. Nishino M, Ashiku SK, Kocher ON, Thurer RL, Boiselle PM, Hatabu H. The thymus: a comprehensive review. Radiographics. 2006;26:335-48.
3. Miller J. Immunological function of the thymus. Lancet. 1961; 2:7489.
4. Boehm T, Bleul CC. The evolutionary history of lymphoid organs. Nat Immunol. 2007;8:131-5.
5. Blackburn CC, Manley NR. Developing a new paradigm for thymus organogénesis. Nat Rev Immunol. 2004;4:278-89.
6. Miller JF. The discovery of thymus function and of thymus-derived lymphocytes. Immunol Rev. 2002;185:7-14.
7. Ribatti D, Crivettato E, Vacca A. Miller's seminal studies on the role of thymus in immunity. Clin Exp Immunol. 2006; 144:371-5.
8. Anderson G, Jenkinson WE, Jones T, Parnell SM, Kinsella FA., White AJ et al. Establishment and functioning of intrathymic microenvironments. Immunol Rev. 2006;2009:10-27.
9. Itoi M, Tsukamoto N, Yoshida H, Amagai T. Mesenchymal cells are required for functional development of thymic epithelial cells. Int Immunol. 2007;19:953-64.
10. Bhandoola A, Sambandam A. From stem cell to T cell: one route or many? Nat. Rev. Immunol. 2006,6:11726.
11. Krueger A, Willenzon S, Lyszkiewicz M, Kremmer E, Forster R. CC chemokine receptor 7 and 9 double-deficient hematopoietic progenitors are severely impaired in seeding the adult thymus. Blood. 2010;115:1906-12.
12. Wurbel MA, Malissen B, Campbell JJ. Complex regulation of CCR9 at multiple discrete stages of T cell development. Eur J Immunol. 2006;36:73-81.
13. Uehara S, Grinberg A, Farber JM, Love PE. A role for CCR9 in T lymphocyte development and migration. J Immunol. 2002;168:2811-9.
14. Abbas AK, Lichtman AH. Lymphocyte Maduration and Expression of Antigen Receptor Genes. In: Abbas AK, Lichtman AH, ed. Cellular and Molecular Immunology. Philadelphia: Elsevier Saunders; 2005. p. 129-62.
15. Schwarz BA, SambandaM A, Maillard I, Harmar B, Looe P, Bhandoola A. Selective thymus settling regulated by cytokine and chemokine receptors. J Immunol. 2007;178:2008-17.
16. Nunes H, Ribot JC, Serra A, Silva B, Sousa AE. Foxp3 induction in human and murine precedes the CD4+ CD8+ stage but requires early T-cell receptor expression. Immunol Cell Biol. 2010;88:523-8.
17. Starr, TK, Jameson SC, Hogquist KA. Positive and negative selection of T cells. Annu Rev Immunol. 2003;21:139-76.
18. Hayday AC. ãæ T cell: a right time and a right place for a conserved third way of protection. Ann Rev Immunol. 2000;18: 975-6.
19. Maggi E, Cosmi L, Liotta F, Romagnani P, Romagnani S, Annunziato F. Thymic regulatory T cells. Autoimmune Rev. 2005 Nov;4:579-86.
20. Klein L, Jovanovic K. Regulatory T cell lineage commitment in the thymus. Semin Immunol 2011 Dec;23:401-9.
21. Winrsberger G, Hinterberger M, Klein L. Regulatory T-cell differentiation versus clonal deletion of autoreactive thymocytes. Immunol Cell Biol 2011 Jan;89:45-53.
22. Al-Shami A, Spolski R, Kelly J, Fry T, Schartzberg PL, Pandey A, et al. A role for thymic stromal lymphopoietin in CD4(+) T cell development. J Exp Med 2004; 200:159-68.
23. Liu Y-J, Soumelis V, Watanabe N, Ito T, Wang Y-H, Malefyt RdW, et al. TSLP: an epithelial cell cytokine that regulates T cell differentiation by conditioning dendritic cell maturation. Annu Rev Immunol 2007;25:193-219.
24. Reche PA, Soumelis V, Gorman DM, Clifford T, Liu MR, Travis M, et al. Human thymic stromal lymphopoietin preferentially stimulates myeloid cells. J Immunol 2001;167: 336-43.
25. Appay V, Sauce D, Prelog M. The role of the thymus in immunosenescence:lessons from the study of thymectomized individuals. AGING 2010;2:278-81.
26. Caruso C, Buffa S, Cardore G, Colonna-Romano G, Dunn-Walters D, Kipling D, et al. Mechanisms of immunosenescence. Immun Ageing 2009;6:10-4.
27. Sansoni P, Vescovini R, Fagnoni F, Biasini C, Zanni F, Telera A, et al. The immune system in extreme longevity. Experimental Gerontology 2008;43:61-5.
28. Weiskopf D, Weinberger B, Grubeck-Loebenstein B. The aging of the immune system. Transpl In 2009;22:1041-50.
29. Herrero C, Celada A. Inmunosenescencia. Inmunología. 2000;19(2,3): 75-80.
Recibido: Enero 7, 2013
Aceptado: Abril 3, 2013
Dra. Vianed Marsán Suárez. Instituto de Hematología e Inmunología. Apartado 8070, La Habana, CP 10800, CUBA.
Tel (537) 643 8695, 8268
Fax (537) 644 2334
Email: rchematologia@infomed.sld.cu


{kind=link}
{kind=link}